Correction to “Incorporation of Multiple β2‑Hydroxy Acids into a Protein in Vivo Using an Orthogonal Aminoacyl-tRNA Synthetase”
Noah X. Hamlish, Ara M. Abramyan, Bhavana Shah, Zhongqi Zhang, Alanna Schepartz

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
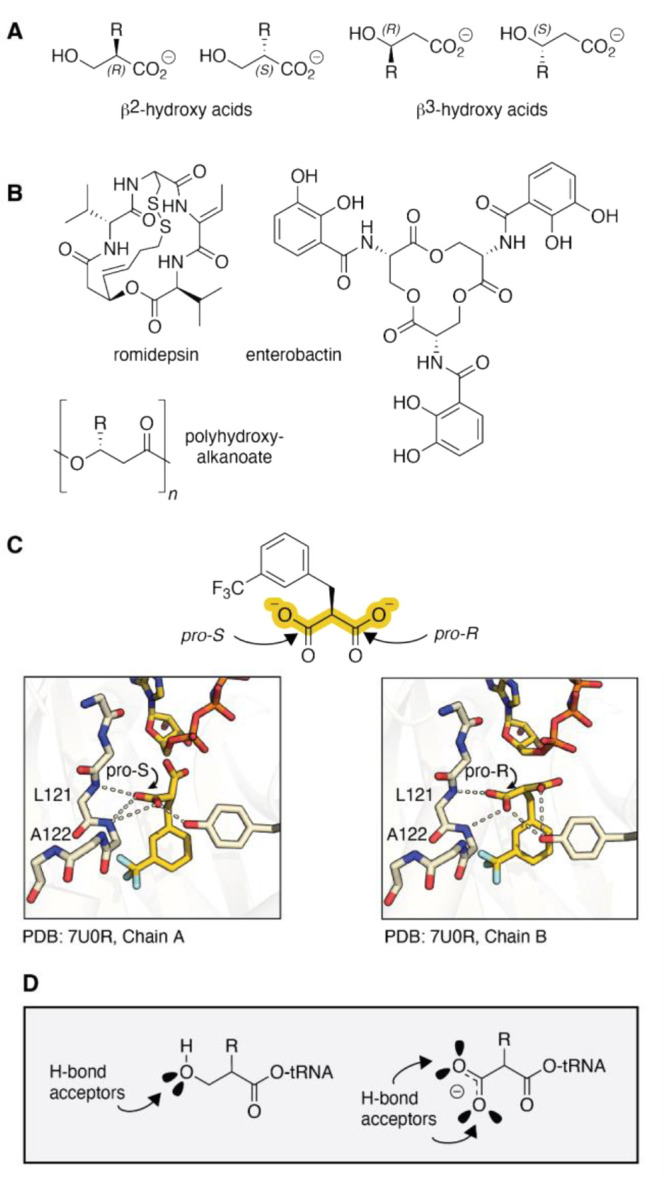 Figure 1
Figure 1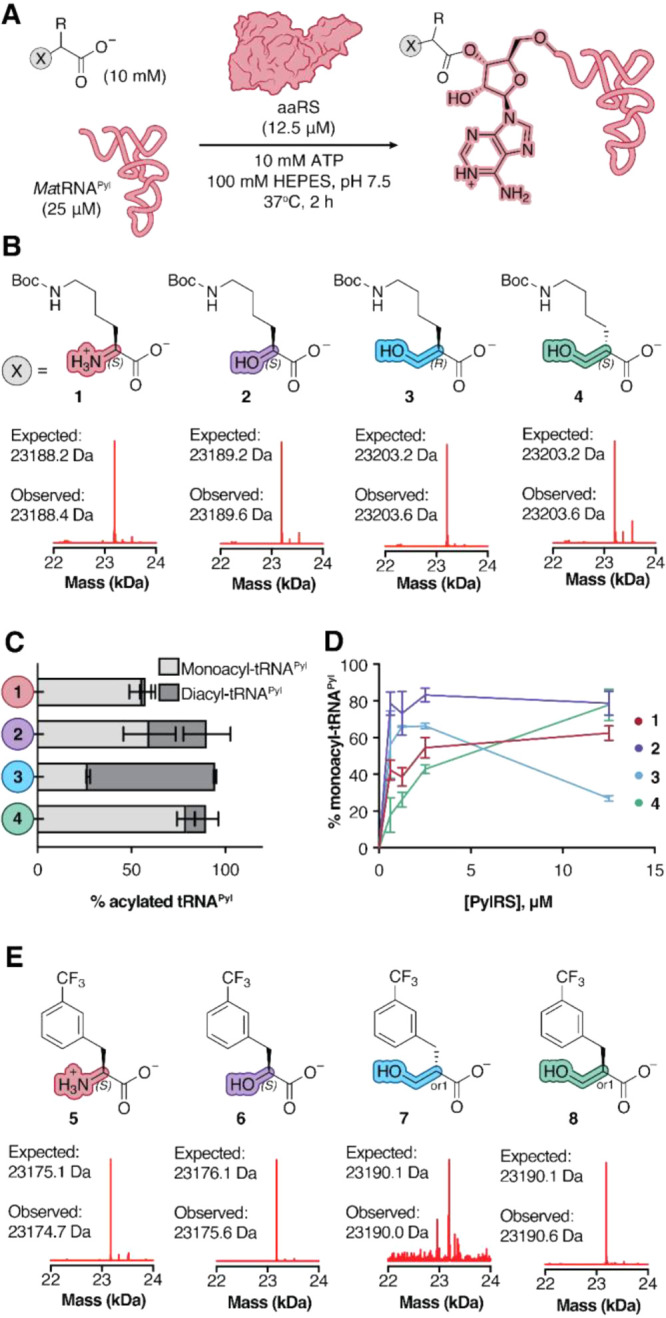 Figure 2
Figure 2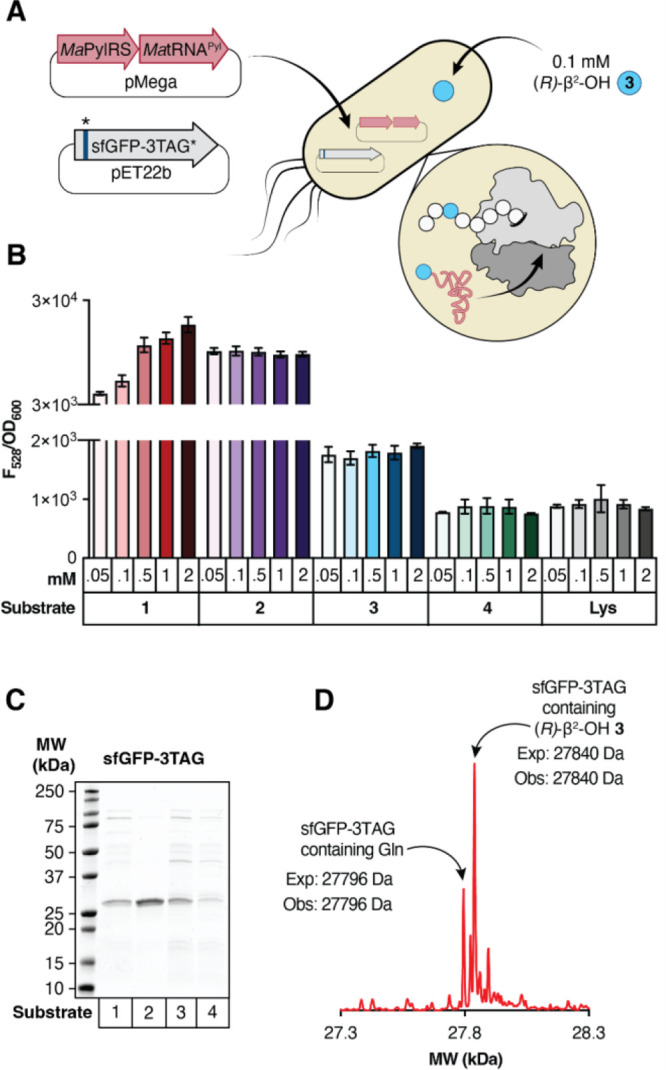 Figure 3
Figure 3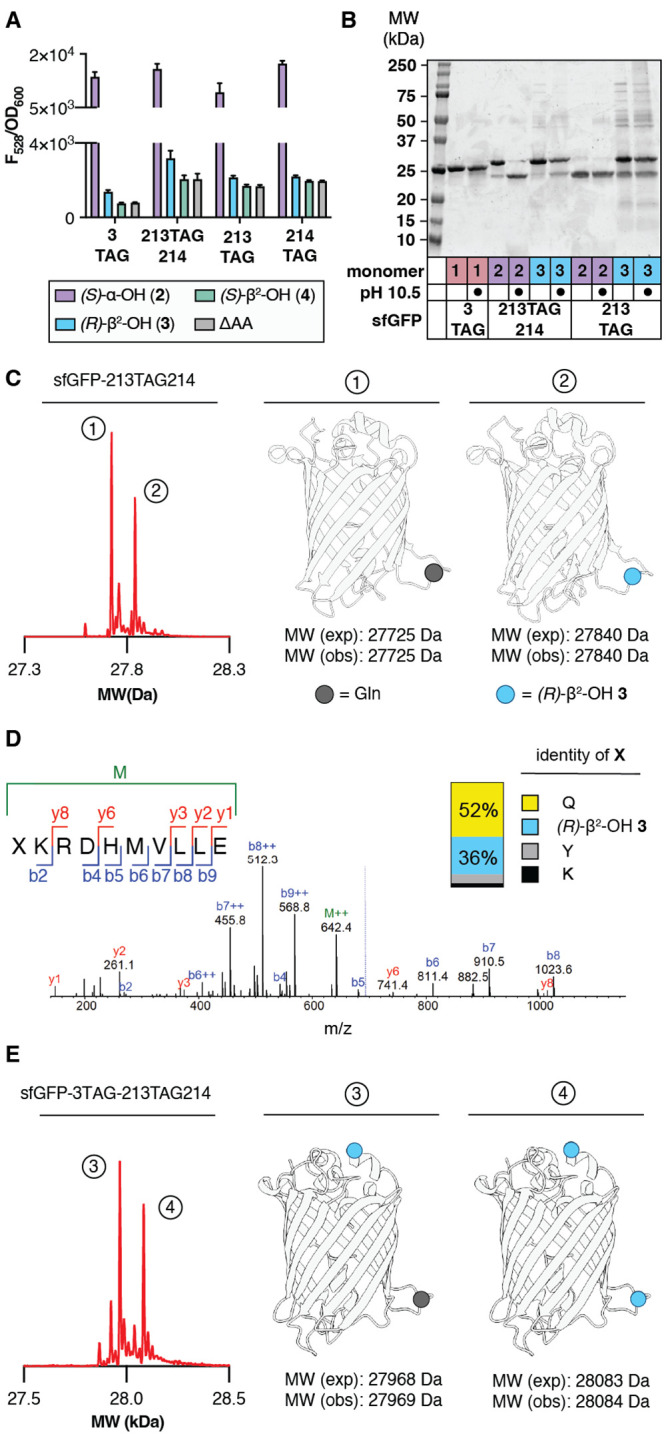 Figure 4
Figure 4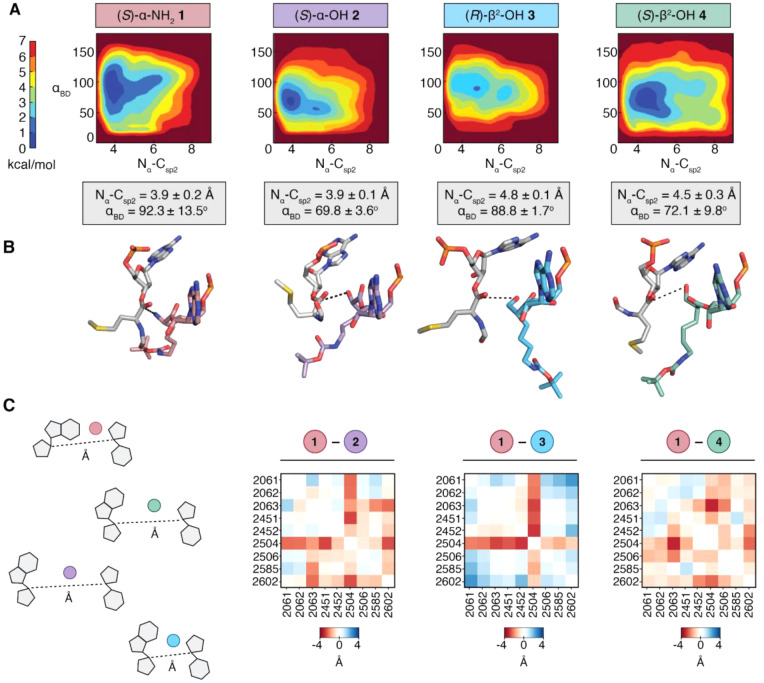 Figure 5
Figure 5 Figure 6
Figure 6- —National Science Foundation10.13039/100000001
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA and protein synthesis mechanisms · Chemical Synthesis and Analysis · DNA and Nucleic Acid Chemistry
After this manuscript was published, we became aware that the stereochemistry of the two β^2^-hydroxy acid monomers*(R)-β^2^-OH-BocK and (S)-β^2^-OH-BocKhad been inadvertently misassigned by the supplier. The correct assignment was inadvertently detected during cryo-EM analysis of ribosomes bound to tRNAs acylated with the two enantiomeric monomers. The molecule we refer to throughout the published paper as (S)-β^2^-OH-BocK is in fact (R)-β^2^-OH-BocK and vice versa. Two additional β^2^-hydroxy acid monomers(S)*-β^2^-OH-m-CF_3_-Phe and (R)-β^2^-OH-m-CF_3_-Phe (referred to as compounds 7 and 8, respectively)were discovered to be enantiopure but without characterization of the absolute configuration of their respective stereocenters.
Although regretful, this finding neither impacts nor alters the two key findings of the published work, namely that (1) MaPylRS acylates MatRNA^Pyl^ with either (S)-β^2^-OH-BocK or (R)-β^2^-OH-BocK in vitro; and (2) in cells, MaPylRS supports the incorporation of two copies of a β^2^-hydroxy acid monomer into protein. The difference is that the monomer introduced in cells is (R)-β^2^-OH-BocK not (S)-β^2^-OH-BocK.
Although these two key findings are unaffected, the manuscript requires the correction of structures drawn in the Table of Contents graphic, Figures, ?, ?, and ?, and Supplementary Figures 1–4, 7–8, and 10–11. It also requires a modest rewording of the paragraphs interpreting the aaRS activity of compounds 7 and 8 as well as the metadynamics simulations. The corrected main text figures and figure legends are provided here along with the reworded paragraphs. Although unrelated, we also noticed that 3 PylRS residues were un- or mislabeled in FigureC. A corrected version of that figure is also provided.
Corrections to Main Figures
Corrections to the table of contents graphic and Figures ?–? are as follows. Corrected figures appear below.
TOC graphic corrections: The structure of the monomer shown was changed to indicate that only the (R) stereoisomer is introduced into protein in cells.
Figure 1 corrections: Amino acid residues within PylRS in panel C were mislabeled in the original manuscript and have been updated with proper labeling. No corrections to text.
Figure 2 corrections: (1) In panel B, the stereochemistry of 3 and 4 has been updated to reflect the correct absolute stereochemistry. (2) In panel E, compounds 7 and 8 have been updated with an “or1” nomenclature as provided by the supplier (Enamine). This nomenclature is used to describe enantiopure compounds of unknown absolute configuration. Please also see the text correction below.
Figure 3 corrections: (1) In panel A, labeling for compound 3 has been updated with the correct stereochemistry. (2) In panel D, labeling of the deconvoluted mass spec peak has been updated with correct stereochemistry. (3) In panel F, stereochemistry has been updated for monomer 3. Text throughout the manuscript has been updated with accurate stereochemistry.
Figure 4 corrections: (1) In panel A, labeling for compound 3 has been updated with the correct stereochemistry. (2) In panel C, labeling of the blue ball representing compound 3 has been updated with the correct stereochemistry. (3) In panel D, labeling of monomers in compositional analysis updated with accurate stereochemistry. Text updated to match.
Corrected Paragraphs
Correction to Section: “Introduction”, Paragraph
5
Like the α-carboxy group of a malonic acid, a β^2^-hydroxyl group of tRNA acylated with a β^2^-HA can also accept at least one, and perhaps two H-bonds from the synthetase amide backbone (FigureD). Here we report that β^2^-hydroxy acids possessing both (R) and (S) absolute configurations are substrates for PylRS enzymes in vitro. Further, we report that only (R)-β^2^-hydroxy acidswhose absolute configuration maps onto a l-α-amino acidare substrates in cellulo. Using the orthogonal MaPylRS/MatRNA^Pyl^ synthetase/tRNA pair and classic E. coli expression strains (BL21 and C321.ΔA.exp), we report the cellular synthesis of model proteins containing up to two (R)-β^2^-HA residues at internal positions. Metadynamics simulations do not provide a clear rationale for the observed enantioselective preference for the (R)-β^2^-hydroxy acid suggesting that this selection may occur at least in part prior to ribosomal translation. As far as we know, this finding represents the first example of an orthogonal synthetase that acylates tRNA with a β^2^-hydroxy acid substrate and the first cellular biosynthesis of a protein hetero-oligomer containing multiple expanded backbone monomers.
Correction to Section: “MaFRSA Acylates
tRNAPyl with a β2-Hydroxy Acid Substrate in Vitro”, Paragraph 1
The PylRS derivative FRSA contains two active site mutations (N166A and V168A) that favor substrates with substituted Phe side chains.^24^ One of the best substrates for FRSA is the α-amino acid m-CF_3_-Phe 5 (FigureE). To determine if MaFRSA would also acylate tRNA with a β^2^-HA, we performed in vitro tRNA acylation reactions supplemented with m-CF_3_-Phe 5 alongside analogous reactions containing (S)-α-OH 6 and the β^2^-OH analogs 7 and 8 (enantiopure monomers whose absolute configurations have not been determined as indicated by the “or1” nomenclature, FigureE). Reactions performed with 12.5 μM MaFRSA, 25 μΜ MatRNA^Pyl^ and 10 mM 5 or 6 cleanly generated the expected monoacylated tRNA products 5-acyl-tRNA^Pyl^ (23175.6 Da) and 6-acyl-tRNA^Pyl^ (23176.8 Da) (FigureE). Quantification of the monoacyl-tRNA and unacylated tRNA pools indicates yields of 45% and 29% for 5-acyl-tRNA^Pyl^ and 6-acyl-tRNA^Pyl^ respectively (Supplementary Figure 3). Aminoacylation reactions supplemented with 10 mM enantiopure monomer 7 yielded only a single low signal peak in the TIC corresponding to the molecular weight of monoacyl-tRNA (23189.5 Da) with a yield of <1% 7-acyl-tRNA^Pyl^ (Supplementary Figure 3). By contrast, supplementation of an analogous aminoacylation reaction with enantiopure monomer 8 yielded two peaks in the TIC corresponding to monoacylated (23189.5 Da) and diacylated 8-acyl-tRNA^Pyl^ (23420.1 Da) (FigureE) in 28% and 11% yield, respectively. The observation that MaFRSA has an enantiopreference for monomer 8 over monomer 7 is consistent with our observation that the parent enzyme MaPylRS exhibits slightly higher activity for (R)-β^2^-OH 3 over (S)-β^2^-OH 4.
Reference 24.
Wang, Y.-S. ; Fang, X. ; Wallace, A. L. ; Wu, B. ; Liu, W. R.
A Rationally Designed Pyrrolysyl-tRNA Synthetase Mutant with a Broad Substrate Spectrum. J. Am. Chem. Soc. 2012, 134, 2950–2953, DOI: 10.1021/ja211972x.22289053 PMC3288562
Correction to Section: “Metadynamics Simulations Probe
Enantioselectivity of the PTC with Respect to β2-OH-Monomers”, Paragraph 3
The free energy surfaces for an RRM containing **3-**acyl-tRNA^fMet^ or **4-**acyl-tRNA^fMet^ are defined by different global minima values. For the RRM containing **3-**acyl-tRNA^fMet^ in the A-site, we observe low energy poses characterized by N_α_–C_sp2_ distances of 4.8 ± 0.1 Å and α_BD_ values of 88.8° ± 1.7°. The N_α_–C_sp2_ distance for **3-**acyl-tRNA^fMet^ falls outside the N_α_–C_sp2_ distance range for monomers predicted to be highly reactive in the ribosome and suggests that the relatively low incorporation of (R)-β^2^-OH 3 is due in part to poor sampling of conformations within the PTC that support rapid bond formation. The free energy surface of an RRM containing **4-**acyl-tRNA^fMet^ is defined by a similar averaged N_α_–C_sp2_ distance and α_BD_ but with larger standard deviations (N_α_–C_sp2_ = 4.5 ± 0.3 Å and α_BD_ = 72.1° ± 9.8°). Indeed, the energy minima of metadynamics trajectories for monomer 4 produces a minimized structure in which the β–OH of **4-**acyl-tRNA^fMet^ is turned toward the P-site carbonyl electrophile (FigureB). These data suggest that there are minimal differences in suitability of tRNAs acylated with (S)- and (R)-β^2^-OH-BocK as substrates for WT E. coli ribosomes, and that a bottleneck prior to bond formation precludes the incorporation of (S)-β^2^-OH 4. In support of this conclusion, recent data indicate that the cellular production of tRNA^Pyl^ acylated with (S)-β^2^-OH-BocK is barely detectable, while tRNA^Pyl^ acylated with (R)-β^2^-OH-BocK-acyl-tRNA^Pyl^ is highly abundant.^43^
Reference 43.
Pressimone, M. A. ; Schissel, C. K. ; Goss, I. H. ; Swenson, C. V. ; Schepartz, A.
Monitoring Monomer-Specific Acyl-tRNA Levels in Cells with PARTI. Nucl. Acids Res. 2025, 53, gkaf327, DOI: 10.1093/nar/gkaf327.40335069 PMC12058263
Correction to Section: “Metadynamics Simulations Probe
Enantioselectivity of the PTC with Respect to β2-OH-Monomers”, Paragraph 6
In contrast, the PIA plot comparing the RRMs for **1-**acyl-tRNA^fMet^ and **3-**acyl-tRNA^fMet^ is different. First, it highlights many pairwise interactions that are markedly shorter when **3-**acyl-tRNA^fMet^ occupies the A-site, including those involving U2506, U2585, and A2062. Changes involving U2506 and U2585 are especially notable, as both have been implicated as critical for induced conformational changes required for efficient bond formation.^46^ U2585 is believed to shield the P-site peptidyl-tRNA from hydrolysis in the uninduced state and rotate away in the induced state to expose the ester bond for nucleophilic attack by the A-site monomer.^45^ Indeed, recent cryo-EM structures of E. coli ribosomes with aminobenzoic acid monomers in the A-site show U2585 locked in the uninduced conformation, prohibiting access to the P-site peptidyl-tRNA^46^ These observations further suggest that (S)-β^2^-ΟΗ 4 should be able to function as a suitable monomer for the ribosome. Thus, it is possible that steps prior to translation per se contribute to the apparent preference for ribosomal incorporation of (R)-β^2^-OH 3 over (R)-β^2^-OH 4. More broadly, they emphasize that the complex mechanism of translation, including (but not limited to) EF-Tu- and tRNA-induced conformational changes and essential bound water molecules^46^ must be considered in future ribosome or monomer engineering efforts.
Reference 45.
Martin Schmeing, T. ; Huang, K. S. ; Strobel, S. A. ; Steitz, T. A.
An Induced-Fit Mechanism to Promote Peptide Bond Formation and Exclude Hydrolysis of Peptidyl-tRNA. Nature 2005, 438, 520–524, DOI: 10.1038/nature04152.16306996
Reference 46.
Majumdar, C. ; Walker, J. A. ; Francis, M. B. ; Schepartz, A. ; Cate, J. H. D.
Aminobenzoic Acid Derivatives Obstruct Induced Fit in the Catalytic Center of the Ribosome. ACS Cent. Sci. 2023, 9, 1160, DOI: 10.1021/acscentsci.3c00153.37396857 PMC10311655
Correction to Section: “Discussion”, Paragraph
2
Here we report that β^2^-hydroxy acids possessing both (R) and (S) absolute configuration are excellent substrates for pyrrolysyl-tRNA synthetase (PylRS) enzymes in vitro and that certain β^2^-hydroxy acids are also substrates in cellulo. Unsurprisingly (R)-β^2^-OH 3, the β^2^-OH monomer for which we observe incorporation possesses an absolute configuration that maps onto an l-α-amino acid. One unexpected finding is that (S)-β^2^-ΟΗ 4 which possesses an absolute configuration that maps onto a d-α-amino acid is still predicted to occupy a conformation in the ribosomal A-site that is productive for bond formation. A better understanding of the interactions necessary to facilitate P-site electrophile deshielding could be used to identify d-α-amino acids that are successfully elongated in vivo, another long-sought goal.^48^
Reference 48.
Englander, M. T. ; Avins, J. L. ; Fleisher, R. C. ; Liu, B. ; Effraim, P. R. ; Wang, J. ; Schulten, K. ; Leyh, T. S. ; Gonzalez, R. L. ; Cornish, V. W.
The Ribosome Can Discriminate the Chirality of Amino Acids within Its Peptidyl-Transferase Center. Proc. Natl. Acad. Sci. U. S. A. 2015, 112, 6038–6043, DOI: 10.1073/pnas.1424712112.25918365 PMC4434717
Corrections to Supporting Information
The Supporting Information paragraph and descriptions of changes made to the SI are included at the end of this document. A fully corrected SI is included as a separate document.
We are deeply sorry for the errors in our original manuscript and apologize for any inconvenience that these errors may have caused to others.
Supplementary Material