Transcription factors DksA and PsrA are synergistic contributors to Legionella pneumophila virulence in Acanthamoeba castellanii protozoa
Christopher I. Graham, Andrew J. Gierys, Teassa L. MacMartin, Tiffany V. Penner, Jordan C. Beck, Gerd Prehna, Teresa R. de Kievit, Ann Karen C. Brassinga

TL;DR
This study shows that two bacterial transcription factors, DksA and PsrA, work together to help Legionella pneumophila survive and grow inside protozoa.
Contribution
The study reveals a novel synergistic relationship between DksA and PsrA in regulating Legionella virulence traits.
Findings
DksA1 mutation partially rescues growth defects when PsrA is expressed in trans.
DksA and DksA1 can rescue ΔpsrA growth defects, but PsrA cannot rescue ΔdksA defects.
Structural modeling suggests DksA1 impairs Mg2+ coordination in RNA polymerase, affecting transcription.
Abstract
The environmental bacterium Legionella pneumophila, an intracellular parasite of free-living freshwater protozoa as well as an opportunistic human pathogen, has a biphasic lifestyle. The switch from the vegetative replicative form to the environmentally resilient transmissive phase form is governed by a complex stringent response-based regulatory network that includes RNA polymerase co-factor DksA. Here, we report that, through a dysfunctional DksA mutation (DksA1), a synergistic interplay was discovered between DksA and transcription regulator PsrA using the Acanthamoeba castellanii protozoan infection model. Surprisingly, in trans expression of PsrA partially rescued the growth defect of a dksA1 strain. Whilst in trans expression of DksA expectantly could fully rescue the growth defect of the dksA1 strain, it could also surprisingly rescue the growth defect of a ΔpsrA strain.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
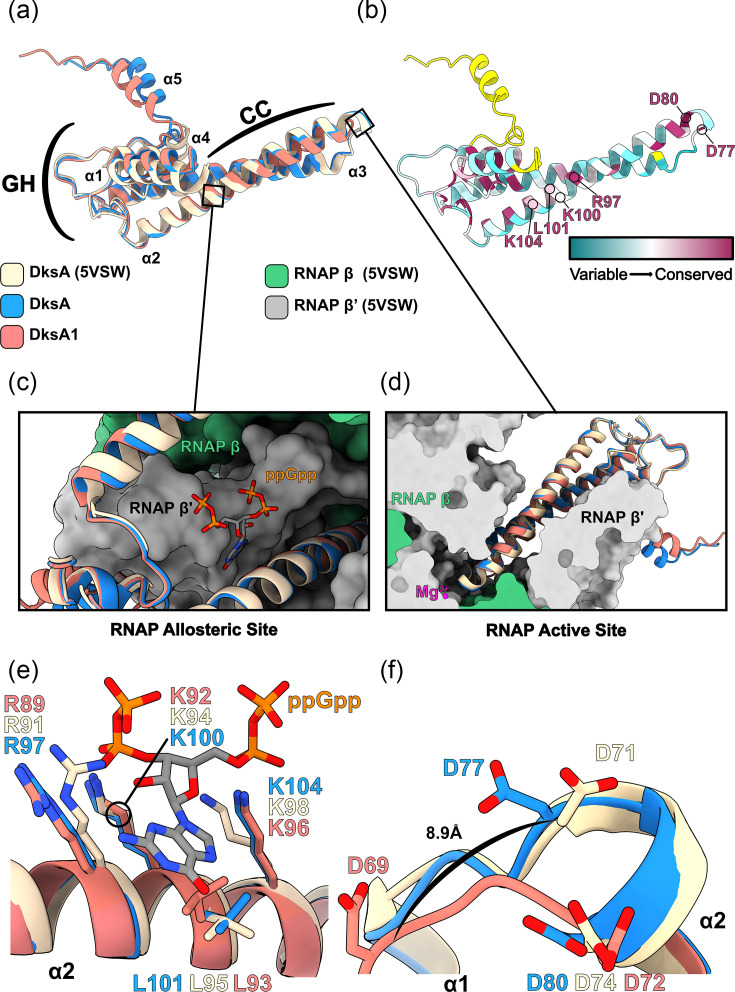 Fig. 1
Fig. 1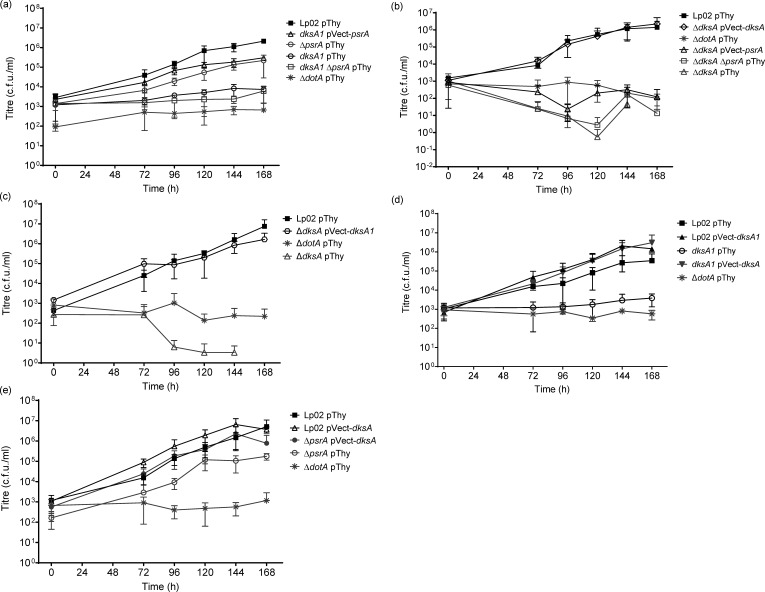 Fig. 2
Fig. 2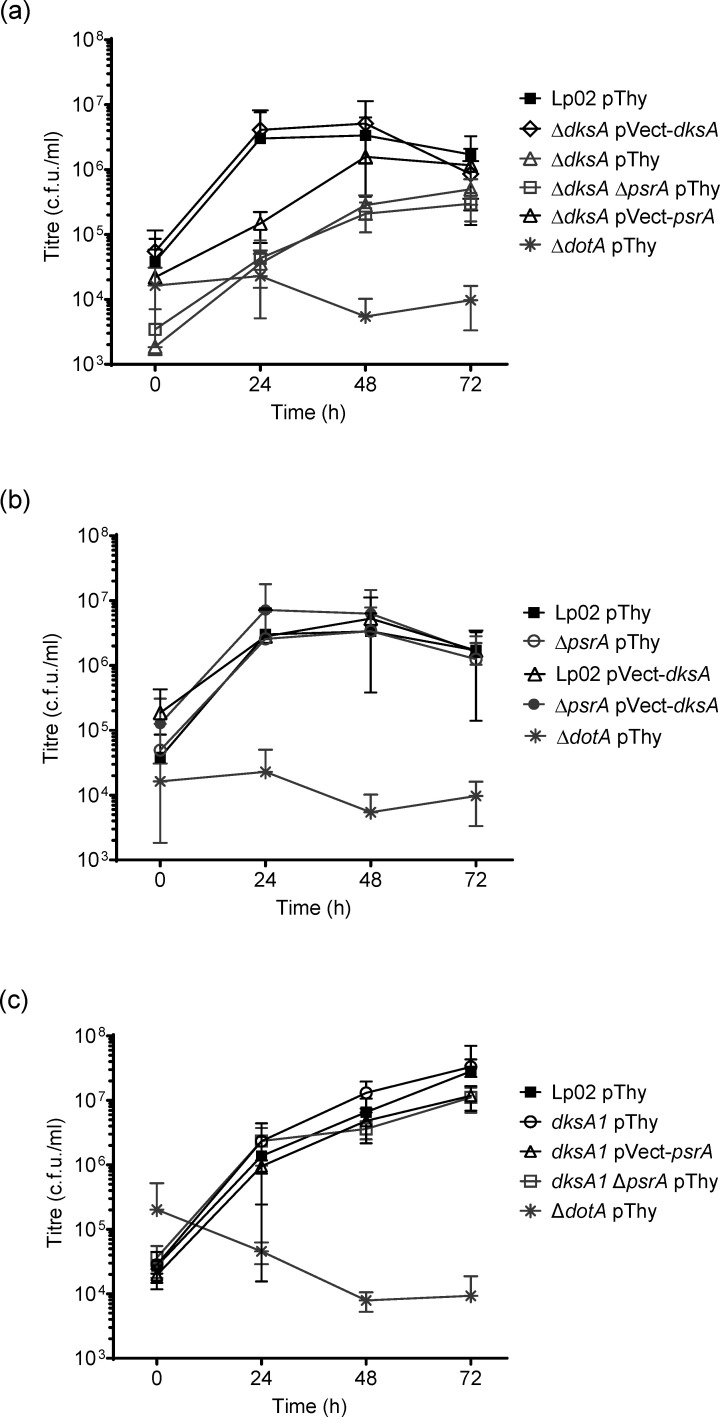 Fig. 3
Fig. 3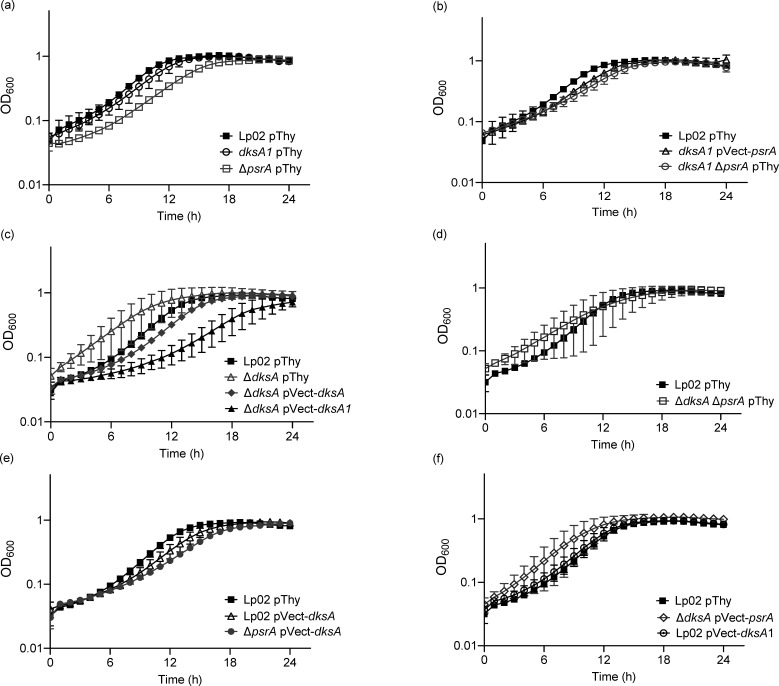 Fig. 4
Fig. 4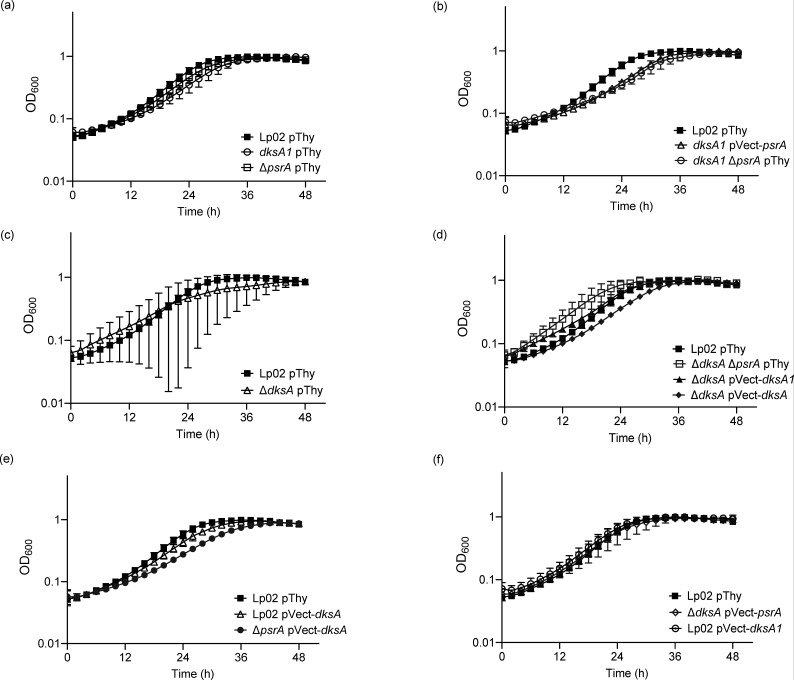 Fig. 5
Fig. 5 Fig. 6
Fig. 6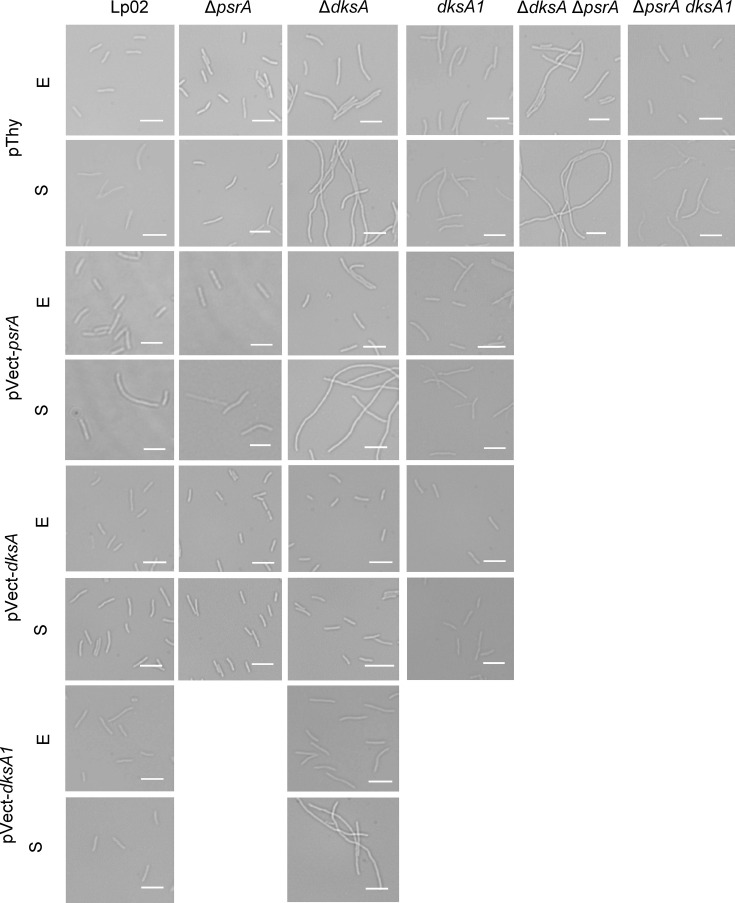 Fig. 7
Fig. 7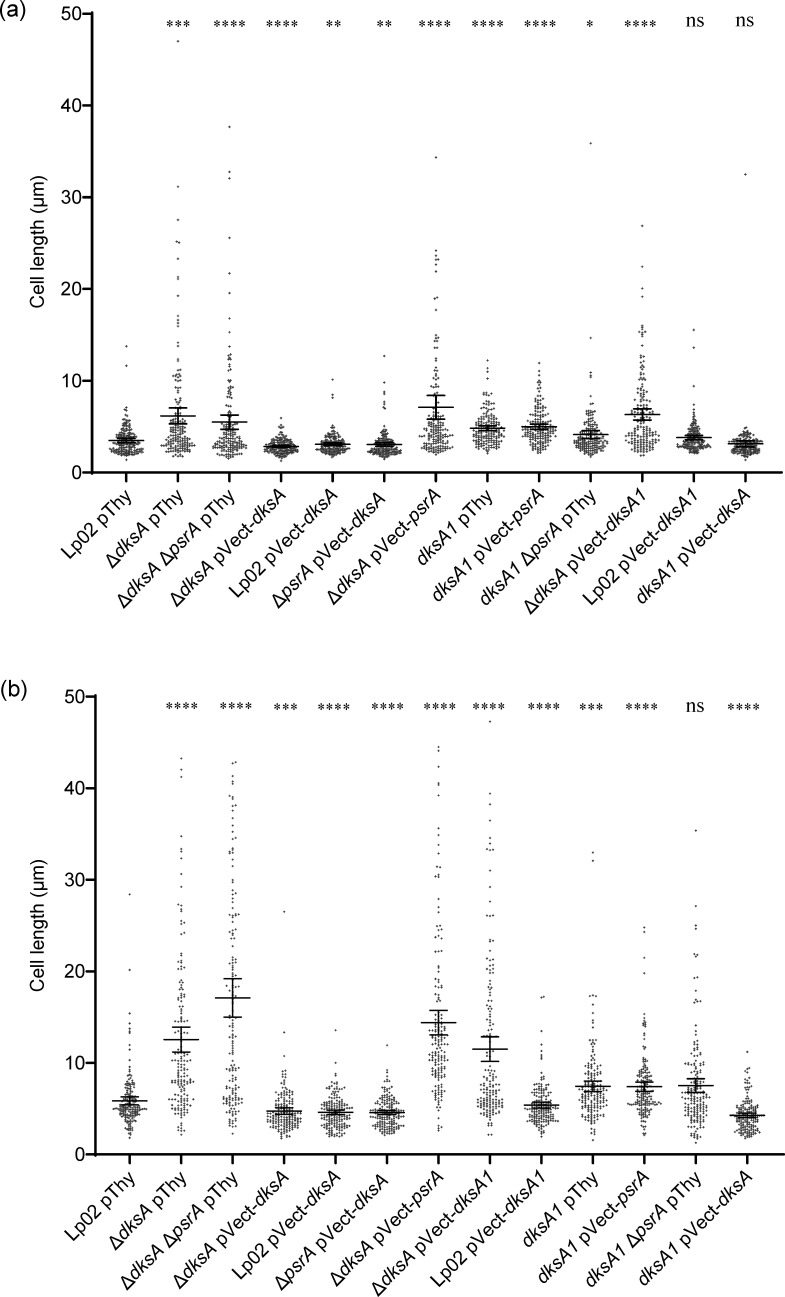 Fig. 8
Fig. 8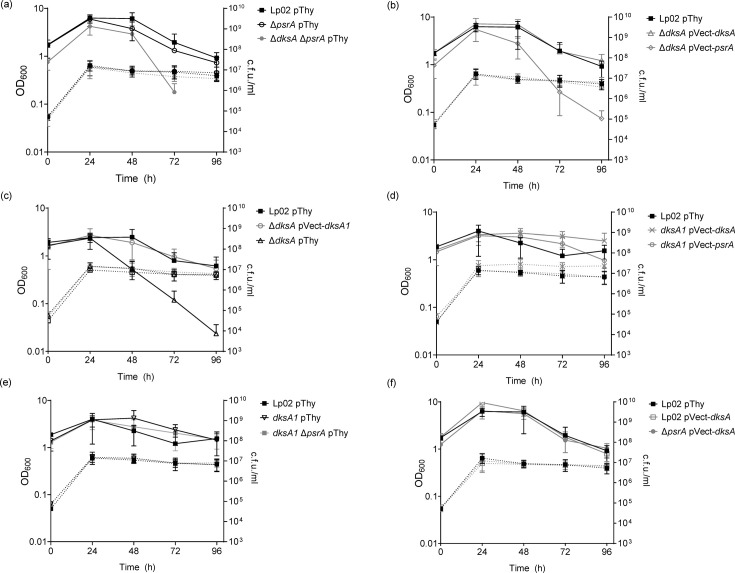 Fig. 9
Fig. 9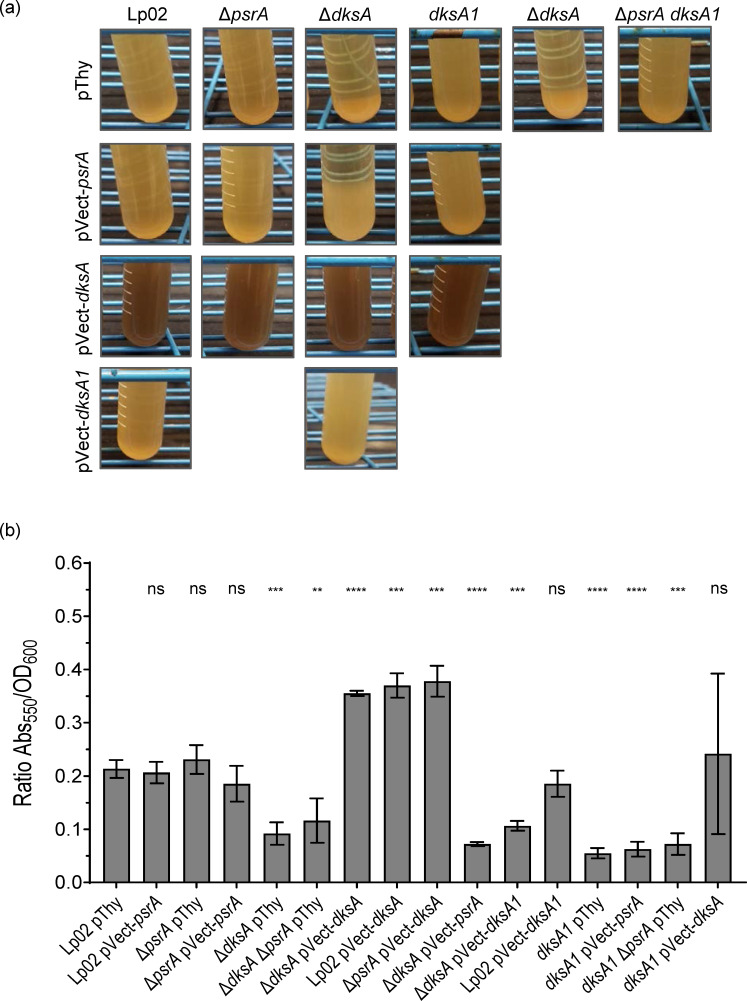 Fig. 10
Fig. 10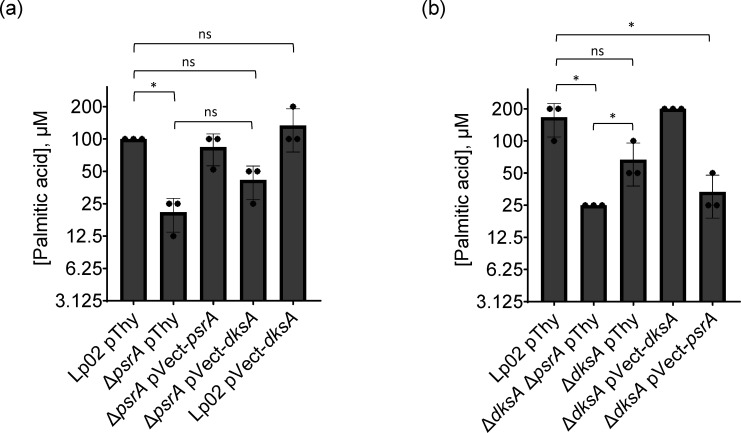 Fig. 11
Fig. 11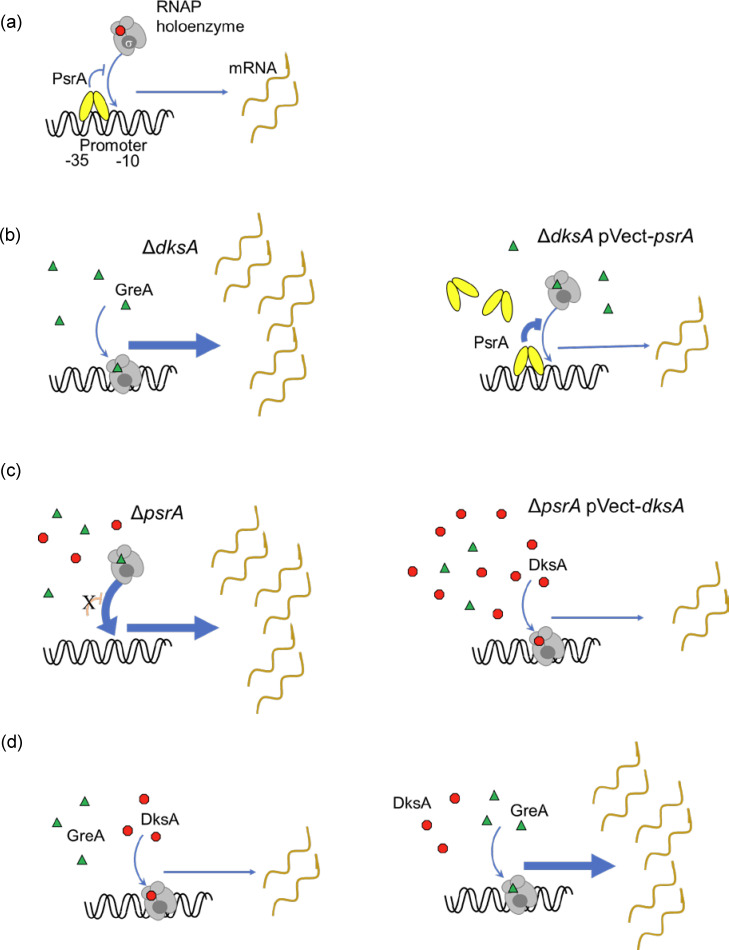 Fig. 12
Fig. 12| No. | Name | Description | Source/reference |
|---|---|---|---|
|
| |||
| DH5α | F′ | New England Biolabs | |
| DH5α λpir | K-12 F- φ80 | Bryan | |
|
| |||
| CG858 | Lp02 | Strr, Thy-, HsdR- derivative of Philadelphia-1 strain | Rao |
| KB265 | Lp03 | Lp02 Δ | Berger and Isberg [ |
| CG859 | Δ | Lp02 Δ | [ |
| CG860 | Lp02 pThy | Lp02 pBH6119 | [ |
| JT708 | Δ | Lp03 pBH6119 | [ |
| CG861 | Lp02 pVect- | Lp02 pJB908- | [ |
| CG862 | Δ | Δ | [ |
| CG863 | Δ | Δ | [ |
| CG799 |
| Lp02 | This study |
| CG819 | This study | ||
| CG813 | Lp02 | This study | |
| CG826 | This study | ||
| CG1053 | Δ | Lp02 Δ | This study |
| CG1054 | Δ | Δ | This study |
| CG1055 | Δ | Δ | This study |
| CG1056 | Δ | Δ | This study |
| CG1057 | Δ | Δ | This study |
| CG1059 | Lp02 pVect- | Lp02 pJB908- | This study |
| CG1060 | Δ | Δ | This study |
| CG1061 | Δ | Δ | This study |
| CG1101 | Δ | Δ | This study |
| CG1102 | Lp02 pVect- | Lp02 pVect- | This study |
| CG1146 | This study | ||
|
| |||
| pJB908 | RSF1010 cloning vector, | Sexton | |
| pBH6119 | RSF1010 promoterless GFP vector, | Hammer | |
| pSR47S | oriTRP4 oriR6K Kanr
| Merriam | |
| pPP619 | pVect- | pJB908- | [ |
| pCG1051 | pVect- | pJB908- | This study |
| pCG1080 | pVect- | pJB908- | This study |
| pCG1069 | pSR47S-5′flk- | pSR47S with 5′ and 3′ flanking regions for Δ | This study |
| Name | Sequence | Annealing temp. (°C) | Product size (bp) |
|---|---|---|---|
|
| |||
| PF 5-flk dksA SalI | gcgataGTCGACgttagaggcagaacttttactctg | 62 | 915 |
| PR 5-flk dksA BamH1 | gcgataGGATCCatcaattaattgtcctgtcatgcc | ||
| PF 3-flk dksA BamH1 | gcgataGGATCCcagggtgcttgattctttcttg | 62 | 978 |
| PR 3-flk dksA SacI | gcgataGAGCTCgtaataagggctctgtagtgagg | ||
| PF comp dksA BamHI | gcgataGGATCCgatttcagcaaagaggcattgg | 62 | 1,016 ( |
| PR comp dksA SacI | gcgataGAGCTCgaatcaagcaccctgattttgg | ||
| PF dksA ext conf | caagcaatcggaataacaaacc | 62 | 842 |
| PR dksA ext conf | cactgtaagccttgttgattttc | ||
| PR dksA conf int | gtcttgcttcaagacgtttcag | 57.5 | 533 (with PF ext dksA conf) |
| pF dksA1 missing* | ggaagaagtagatcgtactgtctc | 57.5 | 320 with pF ext dksA conf* |
| PR dksA1 missing* | gagacagtacgatctacttcttcc | 57.5 | 546 to pR ext dksA conf* |
|
| |||
| PF pSR47s seq | ctgttgcatgggcataaagttg | – | – |
| PR pSR47s seq | cacaggaaacagctatgacc | – | – |
| PF pJB908 seq | ctccgtgatggaatgacaac | – | – |
| PR pJB908 seq | gaccgcttctgcgttctga | – | – |
- —http://dx.doi.org/10.13039/501100000038 Natural Sciences and Engineering Research Council of Canada
- —http://dx.doi.org/10.13039/501100000038 Natural Sciences and Engineering Research Council of Canada
- —http://dx.doi.org/10.13039/501100000038 Natural Sciences and Engineering Research Council of Canada
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLegionella and Acanthamoeba research · Neutrophil, Myeloperoxidase and Oxidative Mechanisms · Vibrio bacteria research studies
Data Availability
The Lp02 dksA1 genome sequence has been deposited into GenBank, under accession number CP184461.
Introduction
Legionella pneumophila, a Gram-negative gammaproteobacterium, is commonly found in piped water distribution systems where it is found as an intracellular parasite of free-living bacterivorous protozoa [13]. These environments are not only amenable to bacterial colonization and proliferation but can also aerosolize Legionella-laden water droplets that can be inhaled by susceptible individuals, thereby facilitating bacterial infection of alveolar macrophages resulting in the atypical pneumonia Legionnaires’ disease [35].
L. pneumophila is well adapted for its intracellular lifestyle in protozoa, and these adaptations also enable human infection due to highly conserved eukaryotic cellular biology [35]. Successful infection is enabled by the Dot/Icm type IV secretion system that facilitates the delivery of ≥330 effectors into the host cell effecting modifications of cellular physiology [68]. Such modifications include subversion of phagosome trafficking to prevent fusion of the bacteria-containing phagosome with lysosomes, and conversion of the phagosome into a replicative niche referred to as the Legionella-containing vacuole (LCV) [911]. As the infection progresses within a protozoan host cell, L. pneumophila exhibits a biphasic lifecycle, alternating between vegetative replicative phase (RP) forms found primarily in the LCV and transmissive phase (TP) forms that develop upon depletion of host-derived nutrients in anticipation of bacterial release to the environment. The physiology and ultrastructure of TP cells are optimized for environmental survival and resistance to stress, with expressed virulence factors enabling infection of a new host [41213].
The biphasic switch between RP and TP is controlled by a stringent response-based regulatory network activated by the accumulation of alarmone guanosine tetraphosphate (ppGpp), which is synthesized by RelA and SpoT in response to nutritional deprivation. Synthase RelA senses low aa concentrations [14], whereas bifunctional synthase and hydrolase SpoT sense fatty acid flux via acyl carrier proteins (ACPs) [1516]. Accumulated ppGpp is associated with activation of the LetA/LetS two-component system (TCS), which in turn induces transcription of non-coding RNAs (ncRNAs) RsmY and RsmZ [1720]. These ncRNAs target and sequester the RNA-binding protein CsrA, thus modulating CsrA-mediated post-transcriptional regulation of transcripts associated with the transition to TP. Such transcripts include those encoding Dot/Icm effectors and those linked to cellular metabolism [2122]. Other TCS also play a pivotal role in bacterial differentiation and are often required for virulence in host cells, such as CpxR/A [2324], PmrA/B [2526] and quorum sensing via LqsS/T/R [2730]. In addition, a subset of transcriptional regulators plays an important role in TP induction. Integration Host Factor assists LetA in the upregulation of RsmY and RsmZ and is required for full cyst-like form differentiation and virulence in Acanthamoeba castellanii [3132]. OxyR regulates transcription of the operon encoding the CpxR/A TCS, linking it to virulence in A. castellanii [33]. Recently, the autorepressor PsrA was found to be required for optimal intracellular growth and cyst-like form biogenesis in A. castellanii [34]. Whilst the genetic targets of PsrA have not yet been identified, a potential linkage between PsrA and the core stringent response network has been described with a focus on fatty acid flux and an associated ACP [34].
Accumulated ppGpp also directly affects transcriptomic changes associated with the transition from RP to TP. For example, the expression of stationary phase (SP) sigma factor RpoS is increased [16], which is required for the expression of TP traits and intracellular growth in protozoa such as A. castellanii [3538]. Whilst the precise mechanism has not yet been ascertained in * L. pneumophila*, ppGpp influences expression most likely by binding directly to RNA polymerase (RNAP), as has been described in Escherichia coli where ppGpp binds with equal affinity either to a site encompassed by the ω and β’ subunits or at the allosteric site interface between RNAP and secondary channel co-factor DksA. Regarding the latter event, DksA features a pair of conserved aspartate residues to enable ppGpp binding, which together results in a conformational change in RNAP [3942]. This affects the stability of the open complex generated by the RNAP holoenzyme, thereby altering the efficiency of transcription initiation [41,4347]. DksA can act independently of ppGpp, compensate for its absence if overexpressed or even act antagonistically to ppGpp at different promoters [4851]. DksA exists in a dynamic equilibrium with structural homologues GreA and GreB in * E. coli*, which compete with DksA for secondary channel binding. If either GreA or GreB binds, then DksA-independent ppGpp binding to RNAP still occurs to influence expression [52].
L. pneumophila encodes both GreA and DksA orthologues, although GreA has not yet been targeted for study. Like its E. coli orthologue, DksA acts independently of, or in coordination with, ppGpp to regulate genes encoding TP traits [16]. Previously, DksA was shown to be required in a ppGpp-dependent manner for TP differentiation including flagellar biosynthesis. Likewise, DksA and ppGpp are both required for optimal intracellular growth in A. castellanii. Conversely, DksA, but not ppGpp, is dispensable for intracellular growth in murine macrophages, indicating that ppGpp may act independently of DksA in L. pneumophila [16]. Thus, the contributions of ppGpp and DksA to the stringent response are well characterized among the proteobacteria, though the mechanisms and targets may vary between them [53].
In this study, we describe an interplay between DksA and PsrA in the regulation of virulence in protozoa, discovered through a hypomorphic mutant of DksA (DksA1). We show that nominal intracellular growth can be fully achieved with DksA compensating for the lack of PsrA but only partially achieved with PsrA supplementing DksA1, suggesting that PsrA transcriptionally assists DksA. Structural modelling of DksA1 suggests that the transcription efficiency of this RNAP co-factor is compromised, thereby highlighting the proposed synergistic role of PsrA. Further, only DksA is essential for the in vitro phenotypic traits of extended culturability, cell morphology and pigmentation. Conversely, in vitro evidence suggests that the long-chain fatty acid (LCFA) metabolic response is mediated by PsrA together with DksA. Taken together, we show that PsrA and DksA have distinct and overlapping biological roles in the regulation of TP, highlighting a complex regulatory interaction between these two regulatory factors.
Methods
Structural predictions and modelling
The aa sequences of DksA and DksA1 were submitted to the CollabFold2 server [54] with default settings. Top rank #1 models were selected for modelling and analysis in UCSF ChimeraX [54]. Structural overlays between Lp02 DksA, DksA1 and DksA (5VSW) [46] were aligned with Matchmaker. Conservation alignments were performed using ConSurf [5556] with the AlphaFold2 model of Lp02 DksA.
Culture of E. coli and L. pneumophila
L. pneumophila Philadelphia-1 derivative Lp02 parental and isogenic strains, as described in Table 1, were grown on buffered charcoal yeast extract (BCYE) agar plates [57] at 37 °C and 5% CO_2_, or in buffered yeast extract (BYE) broth in tubes on a roller drum at 37 °C, supplemented as required with thymidine (100 µg ml^−1^), sucrose (5 % w/v) or kanamycin sulphate (25 µg ml^−1^). * E. coli* was utilized for cloning and plasmid propagation purposes [DH5α for in trans expression vector pJB908 or pThy (pBH6119)], and DH5α λpir (suicide vector pSR47S) was grown on LB agar plates, or broth in tubes on a roller drum, supplemented as required with ampicillin (100 µg ml^−1^) or kanamycin sulphate (40 µg ml^−1^) and incubated at 37 °C.
Molecular methods/reagents, cloning and sequencing
Chemicals were procured from VWR (Ottawa, ON, Canada), Thermo Fisher (Mississauga, ON, Canada) or Sigma-Aldrich (Oakville, ON, Canada). Restriction endonuclease and PCR polymerase (Taq, Q5) and other cloning reagents were procured from New England Biolabs (Whitby, ON, Canada). Oligonucleotide primers, ordered from Life Technologies (Thermo Fisher), were utilized under conditions listed in Table 2. Nucleic acid purification kits were purchased from Qiagen (Toronto, ON, Canada). Sanger sequencing for routine verification of constructs and mutant validations was done on a fee-based service by The Centre for Applied Genomics (The Hospital for Sick Children, Toronto, ON, Canada). Whole-genome sequencing was undertaken by phenol-chloroform isolation of genomic DNA followed by sequencing via PacBio SMRT at Génome Québec (Montreal, QC, Canada) and assembled using Geneious^™^ software.
dksA1 and ΔdksA mutant strains
The dksA1 mutation was identified through whole-genome sequencing of a ΔpsrA mutant strain (i.e. dksA1 ΔpsrA) during experiments for Graham et al. [34]. The dksA1 mutation was then traced back to the electrocompetent cell stock to obtain the dksA1 strain via PCR screening, using the primer set (PF dksA1 missing/PR dksA1 missing) that annealed to the missing 24 bp region within dksA1 in combination with the primer set (PF dksA ext conf/PR dksA ext conf). For these reactions, PF dksA1 missing was paired with PR dksA ext conf, and PF dksA ext conf was paired with PR dksA1 missing, such that each primer set only generated a product to identify the dksA1 allele. For the generation of an in-frame ΔdksA mutant strain, ~1,000 bp 5′ (primer set PF 5-flk dksA SalI/PR 5-flk dksA BamH1) and 3′ (PF 3-flk dksA BamH1/PR 3-flk dksA SacI) regions flanking dksA were PCR-amplified and cloned into suicide vector pSR47S to use for routine allelic exchange. For the generation of in trans expression constructs, either dksA or dksA1 was PCR-amplified from Lp02 or dksA1 mutant CG799, respectively, using primer set (PF comp dksA BamHI/PR comp dksA SacI), cloned into pJB908 and introduced into parental or isogenic mutant strains by electroporation. The in trans psrA expression vector was from a previous study (originally referred to as pComp, henceforth renamed to pVect-psrA in this study) [34]. The use of the pThy empty vector pBH6119 is used to rescue thymidine auxotrophy of Lp02. Plasmid constructs and mutations were routinely verified by sequencing isolated plasmids or PCR amplicon products, and mutant strains were routinely subjected to whole-genome sequencing to rule out secondary mutations using the reference Lp02 genome sequence [58] for comparative purposes. It should be noted that the parental Lp02 (CG858) strain used in this study contained an in-frame 21 bp insertion mutation in a region linking the receiver and histidine phosphotransfer domains of LetS. The presence of this mutation is considered benign as the biological activity of LetS was unaffected in vivo (A. castellanii protozoa) (data not shown). The GenBank accession number for the genome sequence of the dksA1 strain is CP184461.
In vitro growth kinetics
L. pneumophila strains were grown on plates for 3 days under standard conditions. Bacterial cell mass was collected with a disposable loop and suspended in pre-warmed BYE broth and normalized to OD_600_=0.15. Subsequently, 150 µl of this suspension was loaded in triplicate into wells within a 96-well flat-bottomed microtitre plate; the lid was sealed with Parafilm and loaded into a BioTek Synergy HTX plate reader, reading OD_600_ hourly for 24 h at 37 °C or every 2 h for 48 h at 25 °C with 150 r.p.m. linear shaking. Results were plotted in GraphPad Prism^™^ 9.1 and analysed via two-way ANOVA analysis.
Culture and intracellular kinetics in A. castellanii protozoa
A. castellanii (ATCC 30234) was propagated in Peptone Yeast Extract Glucose media at 25 °C as previously described [24]. Briefly, infections were undertaken in Acanthamoeba (Ac) buffer by placing ~10^6^ protozoan trophozoites in each well of a set of tissue culture-treated 24-well plates and allowing them to adhere, then overlaying them with bacteria normalized by OD_600_ to a multiplicity of infection (MOI) of 0.1 in AC buffer. After 1 h incubation at 25 °C, the wells were washed with 3×1 ml AC buffer, before incubating in 1 ml per well of AC buffer at 25 °C until the indicated time points. Protozoa were lysed mechanically via vortexing, and the lysate was serially diluted in AC buffer onto BCYE agar plates to enumerate titres. To enhance the enumeration of difficult-to-recover avirulent strains, a portion of the lysate was tenfold concentrated by centrifugation at 21,000 g in a microcentrifuge for 2 min; then, 0.9 volume of the supernatant was drawn off, and the pellet was resuspended in the remaining volume and spotted alongside the serial dilutions. After counting titres, graphs were plotted and analysed (two-way ANOVA) in GraphPad Prism^™^ 9.1.
Culture and intracellular kinetics in U937 human macrophages
Human U937 monocytic cells were propagated and differentiated into macrophages using phorbol myristate acetate (Sigma-Aldrich) as previously described [24]. Cells were grown in RPMI-1640 media (HyClone, Thermo Fisher) supplemented with 10% heat-inactivated FBS (Seradigm, Mississauga, ON) (RPMI+FBS) at 37 °C and 5% CO_2_. RPMI+FBS was used for all washes and incubations in a pre-warmed state. For infection, ~10^6^-activated U937-derived macrophages were placed into each well of a tissue culture-treated 24-well plate, allowed to adhere and overlaid with L. pneumophila strains normalized by OD_600_ to an MOI of 2, for 1 h. The bacteria were washed away with 3×1 ml washes in RPMI+FBS, followed by incubation for 1 h with RPMI+FBS with 100 µg ml^−1^ gentamicin. After a further 3×1 ml washes in RPMI+FBS, the cells were incubated in 1 ml RPMI+FBS until lysis at the indicated time points. Cells were lysed osmotically by replacing the RPMI+FBS with ice-cold HyClone water (Thermo Fisher) and repeated pipetting, then serially diluting the lysate in additional HyClone water and enumerating L. pneumophila titres by spotting the dilutions on BCYE agar plates. After counting, graphs were plotted and analysed via two-way ANOVA in GraphPad Prism^™^ 9.1.
Pigment, morphology and culturability assays
Suspensions of L. pneumophila were prepared as per in vitro growth kinetics. Pigment assays were undertaken in accordance with Wiater et al. [59]. Briefly, inoculated 5 ml BYE broth cultures were incubated for 72 h. To quantify pigment at 24 h time points, 1 ml of culture was withdrawn and pelleted at 21,000 g in a microcentrifuge for 2 min, after which 150 µl of supernatant was loaded in triplicate into 96-well plates and read for absorbance at 550 nm on a BioTek Synergy HTX plate reader [59]. After removal of residual supernatant, the pellet was suspended in 1 ml of 1× PBS pH 7.0 and similarly loaded (150 µl) into a 96-well plate and then read for OD_600_. Blank controls were the same volume of BYE or 1× PBS as appropriate. Results were plotted and analysed using GraphPad Prism^™^ (9.1). Photographed 5 ml samples were inoculated and incubated similarly and, at the specified time, allowed to sit undisturbed for 24 h at ambient temperatures (~22 °C) for cells to sediment out by gravity. Long-term culturability was assessed by inoculating 96-well plates with six wells for each strain, containing 150 µl of an OD_600_=0.15 suspension in BYE. The outermost rows and columns were filled with sterile water to reduce evaporation, and the plates were sealed with Parafilm and incubated on a shaking platform at 37 °C for 4 days. At indicated intervals, cell aggregates were dispersed with pipetting, the OD_600_ was read with a BioTek Synergy HTX plate reader across the wells containing unharvested culture and one well of each strain was harvested and serially diluted for titre on BCYE agar plates. Results were plotted and analysed using GraphPad Prism (9.1). For colony-spot morphology, 10 µl of the residual OD_600_= 0.15 suspension used to prepare these assays was spotted on a BCYE plate with spots spread out to twice the spacing of a standard multi-channel pipette, then incubated for 4 days at 37 °C and 5% CO_2_ and then photographed.
LCFA assays
Suspensions of L. pneumophila strains were prepared as per the in vitro kinetics, at OD_600_=0.15 in BYE broth, except that the medium was supplemented with two-fold serially diluted concentrations of palmitic acid (PA) as a 40 mM stock dissolved in 100% ethanol. All broth including PA dilutions, controls and blanks was normalized to 0.5% ethanol. The suspension was then subjected to a standard in vitro growth curve as described in the in vitro kinetics section. Inhibition was defined as growth of <50% in comparison to that of the same strain in a BYE+0.5% ethanol-only control at the end of 24 h. Data were graphed using GraphPad Prism^™^ 9.1.
Microscopy
Microscopy and cell lengths were conducted as per Graham et al. [34]. Briefly, broth cultures in BYE broth were inoculated at OD_600_=~0.02 and incubated for 18 h to OD_600_=0.5–1.0 at 37°C to enter exponential phase, then were mounted on a 2% w/v agarose pad prepared in water and imaged on an Axio Observer Z1 inverted microscope (Zeiss) equipped with a glycerol-immersion 150X objective. SP cells were imaged by returning the cultures to the roller drum for a further ~24 h to achieve SP and imaged similarly. Cell lengths were quantified using the length tool in ImageJ, and lengths were plotted and analyzed in GraphPad Prism™ (9.1).
Results
dksA1 mutation
In a previous study, we deleted psrA from the parental Lp02 strain [34]. Throughout this process, we noted a propensity for some of the isogenic ΔpsrA strains to acquire secondary mutations in letS or dksA as discovered through routine whole-genome sequencing. One particular ΔpsrA strain was found to contain an in-frame 24 bp deletion within the dksA CDS, which resulted in the loss of eight aa residues after the fifty-seventh residue in the DksA polypeptide (Fig. S1A and B, available in the online Supplementary Material). The fifty-seventh codon was additionally altered from encoding leucine to phenylalanine. Thus, this allele was denoted as the dksA1 mutation that is characterized in this study.
As the truncation appeared in the designated coiled-coil (CC) domain of DksA, we investigated how the truncation could affect the fold of DksA1 using AlphaFold2 [6061]. The polypeptide sequences of DksA and DksA1 were submitted to the colabfold2 server (AlphaFold2) with default settings and were modelled with a moderate-to-high degree of confidence (Fig. S1C and D). The CC domain of DksA was well-built as was the majority of the globular head (GH) domain. Conversely, the eight-aa truncation in DksA1 resulted in a partial disruption of the tip of the CC domain, with AlphaFold predicting a ‘disordered’ loop with moderate confidence (Fig. S1D). The extent of the potential structural changes in the mis-formed tip of the CC domain in DksA1 can be observed when overlaid with a previously solved structure of a DksA:RNAP complex from E. coli (PDB ID: 5VSW) [46]. As shown in Fig. 1(a), all DksA variants align well, except for the altered CC-loop regions near helix ɑ3 and the C-terminal region helix ɑ5 not present in the DksA E. coli crystal structure. Importantly, the truncation did not delete key functional residues shown to be needed for the interaction of DksA with the RNAP allosteric site (K104, L101, K100 and R97) or with the RNAP active site (D80 and D77) [4662] (Fig. 1b). Furthermore, all DksA variant structures can be modelled into RNAP (Fig. S2) slotting into the allosteric site (Fig. 1c) and RNAP active site (Fig. 1d) without significant variation from the experimental DksA structure. However, whilst the AlphaFold predictions for Lp02 DksA and DksA1 had a properly modelled ppGpp binding site (Fig. 1e), the twin-aspartate loop of the CC domain was significantly altered in DksA1 through the shift of D69 (D71 in DksA) by ~8.9 Å (Fig. 1f). These models therefore suggest that whilst the functional residues are conserved from the truncation, DksA1 should be incapable of coordinating Mg^2+^ in the active site of RNAP. Conversely, the structural predictions show that DksA1 is likely still able to bind RNAP. Specifically, DksA1 is predicted to still contact the secondary binding site of ppGpp functionally as an allosteric inhibitor of RNAP through ppGpp-mediated recruitment.
Structural comparison of DksA and DksA1 AlphaFold models. (a) Structural overlay of DksA (PDB ID: 5VSW) with both Lp02 DksA and DksA1 as modelled by AlphaFold. (b) Residue conservation of Lp02 DksA as plotted by the server ConSurf [5556]. Yellow indicates insufficient information in the multisequence alignment. Indicated on the structure are conserved residues important for binding ppGpp and allostery with RNAP (R97 to K104), in addition to contact with the active site (d77–d80). (c) Binding of DksA with the RNAP allosteric site. A DksA:RNAP experimental structure (PDB ID: 5VSW) and aligned AlphaFold models are shown as cartoons, whereas RNAP is drawn as a molecular surface. The structure of ppGpp bound in the complex is also shown. (d) Slice view of DksA showing the contact point of residues D80 and D77 with magnesium in the RNAP active site. (d) Interactions of DksA variants with ppGpp in the RNAP allosteric site are shown and not disturbed by the truncation in DksA1. (e) Interactions of DksA variants with magnesium in the active site of RNAP are shown. (f) AlphaFold predicts that the truncation in DksA1 drastically moves the position of a critical aspartate residue (D69/71/77).
PsrA partially rescues the growth defect of the dksA1, but not ΔdksA, strain in A. castellanii
Given the previously described regulatory roles of DksA and PsrA [1634], the discovery of the ΔpsrA dksA1 strain along with the dksA1 strain presented a fortuitous opportunity to investigate potential regulatory links between the two proteins in the established A. castellanii protozoan infection model. Intracellular growth kinetics were conducted on dksA1 pThy, ΔpsrA pThy, dksA1 ΔpsrA pThy and dksA1 pVect-psrA strains, along with virulent Lp02 pThy and avirulent ΔdotA pThy strains as controls, at 25 °C over 168 h as previously described [2434] (Fig. 2a). As the parental Lp02 is auxotrophic for thymidine, empty vector pBH6119 (pThy) featuring thymidylate synthase (tdΔi) as a selection marker was introduced into the parental and isogenic strains to rescue the auxotrophy, which is more efficient than exogenous supplementation of thymidine [24]. For in trans expression that includes complementation as appropriate, select genes (psrA, dksA1 and dksA) were cloned under the control of their respective native promoter into pJB908 (henceforth referred to as pVect), which features tdΔi as a selection marker. The profiles of Lp02 pThy and ΔdotA pThy strains were as expected, with Lp02 pThy demonstrating a 1,000-fold increase in titre and titres of ΔdotA pThy staying near the initial inoculum concentration. The dksA1 pThy and ΔpsrA dksA1 pThy strains showed no significant increase in titres indicating that the chromosomal single-copy dksA1 mutation was defective for virulence. Surprisingly, in trans PsrA expression in the dksA1 background resulted in a 100-fold recovery of bacterial yield (P=0.0006) to a titre within one log_10_ of the Lp02 pThy strain. This suggested that PsrA was able to partially, but not fully, rescue the avirulent phenotype demonstrated by the dksA1 strain. Whilst a genomic copy of psrA is present in the dksA1 strain, it was shown previously that in trans expression of PsrA from pVect-psrA resulted in an elevated quantity of cellular protein that not only rescued the reduced growth phenotype associated with gene deletion of psrA but also enhanced bacterial yield of Lp02 in A. castellanii [34]. Nevertheless, the surprising ability of PsrA to partially complement the growth defect of dksA1 led us to expand our investigations to define the relationship between the two regulatory factors.
Synergistic impacts of DksA, DksA1 and PsrA on virulence of L. pneumophila in the A. castellanii protozoa. Intracellular growth kinetics of L. pneumophila strains in A. castellanii protozoa at 25 °C. Strains tested were parental Lp02 and avirulent ΔdotA along with isogenic strains featuring single or combinatorial genetic mutations (ΔpsrA, ΔdksA and dksA1) with or without in trans expression of psrA, dksA or dksA1. Error bars represent sd across the mean of three biological replicates, with two technical replicates each. Samples below the detection limit (~10 c.f.u. ml−1) were recorded as zero and omitted from the graph if they lacked countable colonies across all replicates.
A previous study elsewhere reported that an insertional dksA mutant showed a modest intracellular growth defect in A. castellanii at 37 °C with reduced yield initially, but eventually achieving similar titres to Lp02 by the end of the 96 h experiment [16]. This reported phenotype differs in comparison to the significantly reduced intracellular growth phenotype observed with the dksA1 strain in this study, thereby supporting our hypothesis that DksA1 demonstrated an altered function and distinct phenotype compared to the null mutant. To test this, and to determine if the compensatory effect of psrA observed with the dksA1 mutant strain extended to a null deletion mutant strain, we generated single ΔdksA and double ΔdksA ΔpsrA mutant strains. The intracellular growth kinetics of the strains were assessed in A. castellanii under the same conditions (Fig. 2b). Interestingly, the ΔdksA pThy and ΔdksA ΔpsrA pThy mutant strains were both fully avirulent, starting at a slightly lower titre than the ΔdotA pThy strain, then declining to, or below, the detection limit of the assay (~10 c.f.u. ml^−1^). The severe growth defect of the ΔdksA pThy strain was fully rescued by in trans expression of dksA, but not psrA; it is a notable divergence from the rescued phenotype demonstrated by the dksA1 pVect-psrA strain. Taken together, the lack of DksA results in a complete avirulent phenotype in these conditions, which cannot be rescued by PsrA, whereas the hypomorph DksA1 has a less severe impact on intracellular growth that can be partially rescued by PsrA.
DksA1 rescues the growth defect of the ΔdksA strain, and vice versa, in A. castellanii
Since DksA can rescue the severe growth defect of the ΔdksA strain in A. castellanii (Fig. 2b), we next asked if the partially functional DksA1 could likewise do so despite the truncation in the CC structure (Fig. 1). To answer this, the ΔdksA pVect-dksA1 strain was assessed in A. castellanii along with ΔdksA pThy, Lp02 pThy and ΔdotA pThy (Fig. 2c). The severely reduced bacterial yield of the ΔdksA pThy strain was robustly rescued by in trans expression of DksA1 as shown by the growth profile of the ΔdksA pVect*-dksA1* strain (P=0.0002) matching that of the Lp02 pThy strain. It should be noted that this growth phenotype contrasted with the minimal growth profile of the dksA1 pThy strain (Fig. 2a), suggesting that restoration of the virulent phenotype of the ΔdksA strain was due to in trans expression of DksA1 protein (pVect*-dksA1*) of sufficient levels to offset the structural and functional inefficiency. Not unexpectedly, in trans expression of DksA successfully rescued the avirulent phenotype of the dksA1 pThy strain to match the growth profile of Lp02 pThy. This phenotype is most likely due to the in trans expression of DksA of which levels exceeded that of chromosomally-encoded DksA1 (Fig. 2d). In the reverse scenario, the growth profile of Lp02 pVect-dksA1 still matched that of Lp02 pThy, suggesting that, despite the in trans expression of DksA1, the chromosomal-copy production levels of DksA and PsrA in parallel were each sufficient to disguise any effects caused by the dysfunctional DksA1 (Fig. 2a, d).
DksA restores the growth defect of the ΔpsrA strain in A. castellanii
To further define the relationship between DksA and PsrA, we next asked if DksA could reciprocally compensate for the lack of PsrA or generally affect intracellular growth in the presence of PsrA. To answer this, Lp02 pVect-dksA and ΔpsrA pVect-dksA strains were assessed for bacterial yield in A. castellanii along with the Lp02 pThy, ΔdotA pThy and ΔpsrA pThy strains for comparative purposes (Fig. 2e). As previously reported, the ΔpsrA pThy strain consistently maintained a tenfold reduction to that of Lp02 pThy (P=0.029) [34]. The mean bacterial yield of the Lp02 pVect-dksA strain showed a subtle increase of ~1/2 log_10_ over that of Lp02 pThy (P=0.007); this enhancement could also be seen in the ΔdksA pVect*-dksA* complemented strain (Fig. 2d; P=0.0247). Likewise, we found that in trans expression of DksA in the ΔpsrA pVect-dksA strain rescued the growth defect of the ΔpsrA strain, increasing yield by approximately tenfold to match that of Lp02 pThy strain (P<0.001 versus ΔpsrA pThy, not significant versus Lp02 pThy). Although it is tempting to remark on the subtle phenotypes observed upon in trans expression of DksA relative to the very strong phenotypes noted in the dksA mutant strains, this experiment as a whole did not generate large phenotypic differences.
DksA is not required for intracellular growth in human U937-derived macrophages
Some L. pneumophila genes exhibit host-specific requirements, such that a subset is required only for growth in one host type, but not in another. Previous work showed that the human U937-derived macrophage infection model is more permissive to L. pneumophila regulatory mutant strains than A. castellanii, as exemplified by transcriptional regulator ΔpsrA or ΔcpxRA TCS mutants [2434]. DksA was previously shown to be dispensable for growth in primary murine bone marrow macrophages [16]. However, since the phenotype that we observed for the ΔdksA strain in A. castellanii was different from that reported by Dalebroux et al. [16], we decided to ascertain the phenotype using the human monocytic U937 cell line as a representative mammalian cell model. Intracellular growth kinetics were assessed for the same set of strains previously used in the A. castellanii infection assays (Fig. 2). As expected, the bacterial yield of Lp02 pThy increased ~100-fold by the 72 h time point (Fig. 3a), and ΔdotA pThy maintained a steady titre with no increase. In comparison to that of Lp02 pThy, ΔdksA pThy and ΔdksA ΔpsrA pThy showed similarly robust, but offset growth profiles. Additionally, despite normalization of the inoculum by OD_600_, these two strains showed initial quantified titres lower than that of Lp02 pThy (Fig. 3a). Only ΔdksA ΔpsrA pThy showed a significant deviation from Lp02 pThy (P=0.049). The ΔdksA pVect-psrA showed a similar profile to that of Lp02 pThy (Fig. 3a) and did not show the reduced starting titre, but the difference to ΔdksA pThy was not significant. No significant change in phenotype was observed with the remaining set of strains featuring in trans DksA expression, either by complementation (ΔdksA pVect-dksA), overexpression (Lp02 pVect-dksA) or in ΔpsrA pVect-dksA (Fig. 3b). The growth profiles of the dksA1 pThy, dksA1 ΔpsrA pThy or dksA1 pVect-psrA were indistinguishable from Lp02 pThy (Fig. 3c) and lacked even variability in the initial uptake. Thus, these results verify that DksA is neither required for nor does it enhance intracellular growth in U937-derived macrophages.
DksA is dispensable for growth in human macrophages. Intracellular growth kinetics of L. pneumophila strains in U937-derived macrophages at 37 °C. Strains tested were parental Lp02 and avirulent ΔdotA along with isogenic strains featuring single or combinatorial genetic mutations (ΔpsrA, ΔdksA and dksA1) with or without in trans expression of psrA, dksA or dksA1. Error bars represent sd across the mean of three biological experiments with two technical replicates each. Experiments in panels a–c were conducted concurrently but presented separately for clarity.
In vitro growth is impacted by DksA, PsrA and DksA1
The differences observed with the intracellular growth kinetics of isogenic mutant strains with and without in trans gene expression constructs in A. castellanii may be in part due to defects in the growth rates of the bacteria, rather than solely virulence defects in vivo. To distinguish this possibility and more generally characterize the in vitro growth characteristics of the strains, we tested the in vitro growth rates of the strains in BYE broth at 37 °C over a 24 h time period (Fig. 4). Whilst the growth profiles of dksA1 pThy and Lp02 pThy were similar to one another, both were distinct from the growth profile of ΔpsrA pThy, which was significantly reduced in comparison to that of dksA1 pThy (P=0.0092) and Lp02 pThy (P=0.0007) (Fig. 4a) with the latter comparison consistent with our previous study [34]. The growth profiles of Lp02 pThy, dksA1 pVect-psrA and dksA1 ΔpsrA pThy were all similar to one another, indicating that lack of PsrA, or elevated levels of PsrA from in trans expression, did not affect the growth profile of the dksA1 (Fig. 4b). Interestingly, the in vitro growth kinetics of ΔdksA-based strains featured growth variability across replicates as manifested by the large error bars for hourly data points (Fig. 4c, d). Despite this variability, the growth profile of ΔdksA pThy was not considered to be significantly different from that of Lp02 pThy (Fig. 4c). Notably, the growth variability disappeared from the ΔdksA pVect-dksA growth profile, suggesting that this was a complementable phenomenon (Fig. 4c). In trans expression of dksA1 (ΔdksA pVect-dksA1) significantly reduced the growth profile in comparison to that of Lp02 pThy (P=0.0053), indicating that the elevated levels of DksA1 compromised replicative growth (Fig. 4c). As observed with dksA1 ΔpsrA pThy (Fig. 4b), the growth profile of ΔdksA ΔpsrA pThy was similar to that of Lp02 pThy (Fig. 4d) indicating the dispensability of PsrA. In trans expression of DksA in Lp02 (Lp02 pVect-dksA) did not alter the growth profile from that of Lp02 pThy (Fig. 4e). Whilst a significant reduction was observed for the growth profile of ΔpsrA pVect-dksA in comparison to that of Lp02 pThy (P=0.0053; Fig. 4e), the difference was not due to in trans expression of DksA, as a similar difference was observed for ΔpsrA pThy versus Lp02 pThy (Fig. 4a). In trans expression of PsrA did not reduce the growth variability phenotype of ΔdksA, nor was the growth profile significantly different from that of Lp02 pThy (Fig. 4f). In trans expression of DksA1 had no effect, as the growth profiles of Lp02 pVect-dksA1 and Lp02 pThy were similar (Fig. 4f).
L. pneumophila in vitro growth at 37 °C is impacted by DksA, PsrA and DksA1. In vitro growth kinetics of parental Lp02 and isogenic strains featuring single or combinatorial genetic mutations (ΔpsrA, ΔdksA and dksA1) with and without in trans expression of psrA, dksA or dksA1 genes. Plate-grown bacteria were suspended in BYE broth and normalized to initial OD600=0.15, and growth at 37 °C was monitored for OD600 hourly for 24 h. Graphs in panels a–f were experiments conducted concurrently but presented separately for clarity. Error bars are sd across the mean of three independent biological replicates with three technical replicates each.
The in vitro growth profile assays were repeated at 25 °C, albeit over a 48 h time period to account for the slower growth rate, to mimic the incubation temperature of the A. castellanii assays. The resultant growth profiles were mostly similar to those attained at 37 °C, with growth variability of ΔdksA pThy exacerbated at 25 °C as indicated by data points with widened error bars (Fig. 5c). One notable difference was that the growth profile of ΔdksA pVect-dksA1 matched that of Lp02 pThy at 25 °C (Fig. 5d), versus the reduction at 37 °C (Fig. 4c), suggesting that the slower growth rate and/or lower temperature accommodated the dysfunctional nature of DksA1.
L. pneumophila in vitro growth at 25 °C is impacted by DksA, PsrA and DksA1. In vitro growth kinetics of parental Lp02 and isogenic strains featuring single or combinatorial genetic mutations (ΔpsrA, ΔdksA and dksA1) with and without in trans expression of psrA, dksA or dksA1 genes. Plate-grown bacteria were re-suspended in BYE broth and normalized to initial OD600=0.15, and growth at 25 °C was monitored for OD600 every 2 h for 48 h. Graphs in panels a–f were experiments conducted concurrently but presented separately for clarity. Error bars are sd across the mean of three independent biological replicates with three technical replicates each.
These findings together suggest that whilst strains lacking dksA featured more variability in growth profiles, more so at 25 °C than at 37 °C, the in vitro growth assays (Figs45) illustrate that the observed in vivo growth defects are specific to A. castellanii (Fig. 2), and not due to poor growth of the bacterial cells themselves.
DksA affects cell morphology
Throughout this study, it was noted that the colony morphologies of ΔdksA and dksA1 strains grown on BCYE agar plates differed significantly from that of Lp02. In comparison to the opaque beige-white glossy colonies with varied margins featured by Lp02, ΔdksA colonies were textured matte white with defined margins, and dksA1 colonies were smooth glossy white with defined margins. The phenotypes were particularly evident when bacterial suspensions in broth were spotted onto BCYE agar plates as were done in serial dilution assays in this study (Fig. 6). The phenotype was fully rescued by in trans expression of DksA for both ΔdksA and dksA1 strains, whereas in trans DksA1 expression only partially rescued the phenotype of the ΔdksA strain. The colony morphology of the ΔdksA ΔpsrA strain was similar to that of the ΔdksA strain, but not to the ΔpsrA strain, which was similar to that of Lp02, indicating the sole influence of DksA. Finally, in trans PsrA expression did not appear to alter colony morphology in all strain backgrounds assessed (Lp02, ΔdksA, dksA1 and ΔpsrA), confirming that PsrA does not influence this phenotype.
L. pneumophila DksA and DksA1, but not PsrA, affect colony morphology. Parental Lp02 and isogenic strains featuring single or combinatorial genetic mutations (ΔpsrA, ΔdksA and dksA1) with and without in trans expression of psrA, dksA or dksA1 genes. Strains were grown on BCYE plates for 3 days and used to generate a bacterial suspension in BYE at OD600=0.15. Ten microlitres of the suspension were spotted onto BCYE plates in triplicate; then, the plates were incubated at 37 °C at 5% CO2 for 4 days prior to being photographed. Images are representative of one of three biological replicates.
We previously reported that PsrA promoted cell elongation when overexpressed in trans [34]. Conversely, it was reported elsewhere that a lack of DksA influenced cell elongation [16]. To assess whether a genetic relationship between PsrA and DksA was evident in cell structure morphology, and to further explore whether dksA1 generated a phenotype distinct from ΔdksA, bacteria in the exponential growth phase (EP) and in SP were microscopically imaged (Fig. 7) and analysed for cell lengths (Fig. 8a, b). As a control, Lp02 pThy bacteria formed typical rods (mean length 3.6 µm) in EP that elongated into SP (mean length 5.8 µm). The ΔdksA pThy bacteria were notably elongated in EP (mean length 6.2 µm) and, in SP, became extremely elongated into filamentous forms often of sufficient length to often exceed the field of view, averaging 13.2 µm among measurable cells (with the entire cell visible in one field of view). Likewise, the ΔdksA ΔpsrA pThy bacteria were also elongated in EP and strongly filamentous in SP (mean lengths 5.4 µm and 17.1 µm, respectively). Conversely, ΔpsrA pThy bacteria were morphologically indistinguishable from that of Lp02 pThy as previously observed [34]. The filamentation of ΔdksA pVect-dksA bacterial forms was rescued by in trans DksA expression, as they were slightly but significantly shortened relative to Lp02 (2.9 µm and 4.7 µm in EP and SP phases, respectively). Expression of DksA in trans in ΔpsrA pVect-dksA and Lp02 pVect-dksA strains also resulted in slightly but significantly shorter cell lengths in comparison to those of Lp02 pThy and comparable to ΔdksA pVect-dksA in both growth phases. In trans PsrA expression in ΔdksA pVect-psrA did not strongly affect the bacterial cell lengths, and elongation was still present (7.5 µm and 14.2 µm in EP and SP, respectively). These findings suggest that the shortening is mediated by DksA independently of PsrA.
L. pneumophila ΔdksA, but not dksA1 or psrA, causes filamentation. Parental Lp02 and isogenic strains featuring single or combinatorial genetic mutations (ΔpsrA, ΔdksA and dksA1) with or without in trans expression of psrA, dksA or dksA1 genes were grown on BCYE plates and used to inoculate a dilute suspension of OD600=0.02 in BYE broth, then incubated overnight at 37 °C with aeration, to exponential (E) (~18 h, OD600 =~0.5) or stationary (S; 24 h after EP) growth phases, and then live-mounted on a 2% agarose pad prepared in ddH2O and imaged on an Axio Observer Z1 inverted microscope (Zeiss) equipped with a glycerol-immersion 150X objective. Representative microscopic images of bacteria from one of three biological replicates. The scale bar is 5 µm.
*L. pneumophila ΔdksA, dksA1 and psrA contribute to cell length regulation. Parental Lp02 and isogenic strains featuring single or combinatorial genetic mutations (ΔpsrA, ΔdksA and dksA1) with or without in trans expression of psrA, dksA or dksA1 genes were grown from 3-day-old plates subcultured to BYE broth at OD600=0.02, then incubated at 37 °C with aeration to (a) exponential growth phase (overnight, OD600 ~0.5) or (b) stationary growth phase (24 h after exponential phase) and then imaged with an Axio Observer Z1 inverted microscope (Zeiss) equipped with a glycerol-immersion 150X objective, and individual bacterial cell lengths were quantified using the length tool in ImageJ. Where filamentation was evident, only individual cells for which the full length was visible were measured. Data represent >150 individual readings spread equally across three biological replicates. The bar on the graph and error bars represent the mean and 95% CI, respectively. Statistical significance determined by Student’s t-test (*P<0.05; **P<0.01; ***P<0.001; ***P<0.0001; ns, not significant, P>0.05).
The bacterial forms of dksA1 pThy, dksA1 ΔpsrA pThy and dksA1 pVect-psrA strain were all slightly elongated relative to Lp02 pThy in both EP (5.0, 5.1 and 4.3 µm, respectively) and SP (7.5, 7.7 and 7.8 µm, respectively) but were not considered filamentous and did not vary significantly from each other, supporting the hypothesis that the dksA1 allele maintained partial function. Interestingly, the ΔdksA pVect-dksA1 strain did not show rescue of the elongation to resemble dksA1 levels as might be expected (5.6 µm in EP and 12.3 µm in SP). To rule out the simple possibility of the two gene products conflicting in some way, we tested the dksA1 pVect-dksA (plasmid- and chromosomal-based alleles reversed). The bacterial forms of this strain appeared somewhat shortened relative to Lp02 pThy in SP only (4.2 µm) resembling the other in trans DksA expression strains, suggesting that relative protein concentrations may drive the phenotype. Together, this indicates that the previously reported role of PsrA is overshadowed by the more potent role of DksA and that the DksA1 protein shows an intermediate phenotype illustrating that DksA1 is a partially functional protein in these strains.
DksA, or DksA1, is necessary for extended culturability in depleted media
Previously, the insertional mutant ΔdksA strain was also noted to feature reduced recovery of culturable bacteria after extended incubation in depleted media, which the authors referred to as ‘survival’ (i.e. culturability) [16]. Over the course of this study, we noticed that the ΔdksA mutants could not be consistently sub-cultured, or often failed to grow entirely, if taken from BCYE agar plates grown for more than 3 days. In view of this, we asked if this was a consequence of the previously observed culturability phenotype occurring in the ΔdksA strain, and potentially the dksA1 strains, and whether PsrA may also contribute. As this phenotype had not been assessed in the prior PsrA study [34], we decided to include ΔpsrA pThy, ΔpsrA pVect-psrA and Lp02 pVect-psrA strains from Graham et al. [34].
Over the time course, OD values peaked comparably for all strains at 24 h and held steadily to the 96 h time point (Fig. 9). Enumeration of titres for Lp02 pThy and ΔpsrA pThy was comparable to one another (Fig. 9a). The titres for ΔdksA pThy (Fig. 9c) and ΔdksA ΔpsrA pThy (Fig. 9a) were significantly reduced by 72 h, and more so by 96 h for ΔdksA pThy (Fig. 9c), noting that titre enumeration was not possible for ΔdksA ΔpsrA pThy at 96 h, as no colonies were recovered in any replicate for this strain at that time point (detection limit ~10^2^ c.f.u. ml^−1^; Fig. 9a). The reduced culturability phenotype was rescued by in trans expression of DksA (Fig. 9b) or DksA1 (Fig. 9c), but not PsrA (Fig. 9b). The extended culturability of the dksA1 pThy strain was unaffected in comparison to Lp02 pThy, thereby neither requiring nor significantly benefiting from in trans expression of DksA or PsrA (Fig. 9d). The unaltered phenotype of the dksA1 ΔpsrA pThy in comparison to the dksA1 pThy strain further supports the observation that PsrA does not contribute to extended culturability (Fig. 9e). Further, in trans expression of DksA in the Lp02 strain does not significantly improve extended culturability (Fig. 9f). Thus, in summary, PsrA plays no role in extended culturability, whilst DksA is essential for the culturability of Lp02 in depleted media as reported previously [16]. Notably, the hypomorph dksA1 allele has retained this essential function of promoting extended culturability.
Bacterial culturability in depleted nutrient media is dependent on DksA or DksA1, but not PsrA. Panels (a–f) show in vitro growth kinetics of Lp02 and isogenic strains featuring single or combinatorial genetic mutations (ΔpsrA, ΔdksA and dksA1) with or without in trans expression of psrA, dksA or dksA1 genes in BYE broth at 37 °C for 96 h with OD600 values (dotted lines; left Y-axis) and titre enumerated by serial dilution and incubation on BCYE plates (solid lines; right Y-axis) at 24 h intervals. Data are presented as mean and sd of three biological replicates. Points were omitted if results could not be calculated (no colonies recovered, below the detection limit of ~100 c.f.u. ml−1). Graphs in panels a–f were experiments conducted concurrently but presented separately for clarity.
Pigmentation requires fully functional DksA, but not PsrA
L. pneumophila pigment is a brown, melanin-like secreted protein that is released into the supernatant during the transition to SP under the control of the stringent response through RelA [1416]. Interestingly, we observed hyperpigmentation of a subset of parental, isogenic (ΔpsrA and ΔdksA) and dksA1 strains producing DksA in trans (Fig. 10a). To ascertain whether DksA1 and/or PsrA could contribute to this hyperpigmentation, we quantified the pigmentation for each strain as done elsewhere [1659] (Fig. 10b).
*Pigmentation is regulated by DksA, but not PsrA. Parental Lp02 and isogenic strains featuring single or combinatorial genetic mutations (ΔpsrA, ΔdksA and dksA1) with or without in trans expression of psrA, dksA or dksA1 genes were grown on BCYE plates for 3 days and used to generate a 5 ml bacterial suspension (BYE at OD600=0.15) and then incubated at 37 °C for 30 h. (a) Qualitative observation of pigmentation. After 48 h of incubation, tubes were set aside for 24 h at room temperature to allow cells to sediment out and then photographed. Representative image of three replicates. (b) Quantification of pigment at 30 h. After 30 h of incubation, 1 ml of culture was centrifuged at 20,000 g. The supernatant was measured for pigment levels via absorbance at 550 nm, and then, the pellet was resuspended in the same volume of 1X PBS pH 7.0 to measure cell density via OD600. Readings were taken with a plate reader in a 96-well plate relative to appropriate BYE or PBS blanks. Readings reflect Abs550 /OD600. Error bars represent sd across three biological replicates. Statistical significance calculated via Student’s t-test and relative to parental Lp02 pThy is indicated above bars (*P<0.05; **P<0.01; ***P<0.001; ***P<0,0001; ns, not significant, P>0.05).
The Lp02 pThy strain developed moderate pigmentation (Abs_550_/OD_600_=~0.20) that was similarly achieved by ΔpsrA pThy, ΔpsrA pVect-psrA and Lp02 pVect-psrA strains, suggesting that PsrA does not influence pigmentation (Fig. 10b). This supposition is further borne out by the significantly reduced pigmentation (Abs_550_/OD_600_=0.10–0.12) featured by both of the ΔdksA ΔpsrA pThy and ΔdksA pThy strains. Significantly increased levels (i.e. hyperpigmentation) were achieved by the ΔdksA pVect-dksA strain (Abs_550_/OD_600_=0.36–0.38), which exceeded that of Lp02 pThy likely due to in trans production of DksA (Fig. 10b). Indeed, the pigmentation phenotype was elevated by in trans DksA expression in the ΔpsrA strain, but not by in trans PsrA expression in the ΔdksA strain, clearly indicating that only DksA positively influences pigmentation (Fig. 10b). Pigmentation was also significantly and equitably reduced (Abs_550_/OD_600_=0.05–0.09) for the dksA1 pThy and dksA1 ΔpsrA strains. As the phenotype was not rescued by in trans PsrA expression in the dksA1 pThy strain, this further supports the lack of involvement in pigmentation by PsrA (Fig. 10b). Surprisingly, the hypomorph DksA1 could not rescue pigmentation in the ΔdksA strain, but the pigmentation of the dksA1 strain could be induced by in trans DksA expression. The dksA1 pVect-dksA strain showed delayed onset of hyperpigmentation such that only one of three replicates had fully pigmented by 30 h, with all replicates of this strain pigmented by 48 h (Fig. 10a), but at 30 h, the difference was not statistically significant. These findings indicate that the ability of DksA1 to interact with RNAP is impaired as suggested by structural modelling (Figs110b). Taken together, these results indicate that DksA is sufficient to drive pigmentation independently of PsrA.
Growth in LCFAs is mediated by PsrA and DksA
The activation of the biphasic switch is mediated in part by the response to fatty acid flux [1516]. It was shown elsewhere that in vitro supplementation of short-chain fatty acids (SCFAs), such as propionic acid, will trigger the expression of TP traits [15]. Further, the process of differentiation in response to excess SCFAs is mediated by DksA [16]. PsrA is also associated with differentiation, though the precise role remains to be elucidated [34]. The Pseudomonas PsrA orthologue is a sensor of LCFAs of which presence alleviates PsrA-mediated repression of the fadBA5 β-oxidation operon [63]. To investigate where PsrA also responds to LCFAs, ΔpsrA strains with and without in trans gene expression constructs were grown in BYE broth supplemented with the LCFA PA to ascertain the impact on bacterial growth. In comparison to Lp02, a twofold growth inhibition of the ΔpsrA pThy strain was observed, which was rescued by in trans PsrA expression, strongly suggesting that PsrA responds to LCFAs (Fig. 11a). Because chromosomal-based DksA expression is present in the ΔpsrA pThy strain, we next asked if DksA also responded to PA, which could be amplified through in trans DksA expression in the ΔpsrA strain background. Whilst it appears that growth of the ΔpsrA pVect-dksA strain improved in comparison to the ΔpsrA pThy strain, it was not found to be statistically significant (Fig. 11a). Likewise, in trans DksA expression in Lp02 did not significantly improve bacterial growth over that of Lp02 alone (Fig. 11a). To further investigate the role of DksA, ΔdksA-based strains were also assessed. The ΔdksA ΔpsrA pThy strain had a twofold growth inhibition in comparison to Lp02 (Fig. 11b). Growth inhibition of the ΔdksA pThy strain was not considered statistically different from that of Lp02 (Fig. 11b). Surprisingly, in trans PsrA expression in the ΔdksA strain could not fully rescue the growth inhibition phenotype. Taken together, these results suggest that, whilst PsrA is largely responsible for the response to LCFAs, DksA also contributes additively to this response, lending support to the overlapping roles observed for the two regulatory factors for some of the traits in this study.
*PsrA and DksA influence LCFA-mediated growth inhibition. Parental Lp02 and isogenic strains featuring single or combinatorial genetic mutations (ΔpsrA and/or ΔdksA), with or without in trans expression of psrA or dksA genes, grown with and without PA supplementation (as 40 mM stock dissolved in ethanol) in BYE media normalized to 0.5% ethanol, in a 96-well microplate for 24 h with shaking at 37 °C. Inhibition was defined as growth, as assessed by OD600 values, of <50% of that of the same strain in a BYE+0.5% ethanol-only control at the end of 24 h. Bars indicate the mean of the maximum concentration of PA where the indicated strain grew >50% of maximum, over three biological replicates with sd indicated. Dots in bars indicate individual replicate values. Panels a and b were conducted separately. Statistical analyses were done using Student’s unpaired t-test: ns, no significance; =P<0.05.
Discussion
The biphasic lifecycle of L. pneumophila is controlled by the convergence of multiple signals into a sophisticated regulatory network that ultimately acts to induce TP differentiation. The primary signals driving differentiation link to the nutritional status of the environment, mediated largely by induction of the stringent response and the associated downstream regulatory network in association with RNAP-binding cofactor DksA, alternative sigma factor RpoS, TCSs and a subset of transcriptional regulators including PsrA [46465].
This study investigated a spontaneous dksA1 mutation resulting in a truncated DksA1 protein featuring a deletion within its CC domain (Fig. 1) [1639]. The intermediate phenotype observed in bacteria expressing DksA1 relative to DksA can be explained by comparing their AlphaFold2 models to that of E. coli DksA bound to RNAP (Fig. 1) [46]. These models predict DksA1 to be physically incapable of coordinating Mg^2+^ in the active site of RNAP due to the truncated loop in the CC domain (Fig. 1d, f). However, DksA1 is still predicted to overall form the core CC and GH domains [4662]. This would hypothetically still allow DksA1 to bind into the allosteric binding pocket of RNAP (Fig. 1c and 2), thereby recruiting a secondary ppGpp to RNAP. This would partially explain the variance in phenotype of dksA1 as an intermediary between the WT and knockout strain. Likewise, these structural predictions also fit into the model of the interplay between GreA and DksA as allosteric inhibitors/activators of RNAP competing for the same allosteric site [4662].
DksA1 (i.e. ΔdksA1 pThy strain) affected reduced intracellular growth in A. castellanii protozoa, indicating impaired function that was partially, but not fully, rescued by in trans expression of PsrA (Fig. 2a). This surprising finding compelled us to use the DksA1 hypomorph to explore overlapping cellular functions of both regulators, inferring a subset of gene targets jointly regulated by DksA and PsrA. To explore the possibilities that either DksA and PsrA interact with each other, or act in parallel, ΔdksA pThy and ΔdksA ΔpsrA pThy strains were subsequently assessed for growth kinetics in A. castellanii protozoa (Fig. 2b). The ΔdksA pThy strain developed a more severely avirulent phenotype than the dksA1 pThy strain and was rescued by in trans expression of DksA or DksA1 (Fig. 2b and c), but not by PsrA (Fig. 2b). Notably, the avirulent phenotype of the ΔdksA pThy strain contrasts with a previous study showing that a ΔdksA insertional mutant strain only exhibited reduced intracellular growth, rather than avirulence, in A. castellanii [16]. Whilst the assays done by Dalebroux et al. [16] and in this study were done in a comparable manner, the discrepancy in findings could be due to differing experimental conditions as the former conducted A. castellanii infections at 37 °C. Temperature-dependent phenotypes have been seen previously, for example, with Δhfq mutant strains, which only showed an intracellular kinetic defect in A. castellanii at 30 °C, but not at 37 °C [66]. To remain consistent with our previous studies on transcription regulators (CpxR, OxyR and PsrA) [243334], we undertook the A. castellanii infections at 25 °C and showed that the avirulent phenotype of the ΔdksA pThy strain was fully complemented by in trans DksA expression (ΔdksA pVect-dksA), indicating that the phenotype was solely due to the genetic deletion of dksA. Another possibility for the discrepancy is the possibility of different deletion strategies causing polar effects. Sahr et al. [67] annotated L. pneumophila Paris strain dksA in an operon with an uncharacterized alpha/beta hydrolase lpp2286 encoded downstream, and Lp02-derivative strains are likely similar. However, both our study and that of Dalebroux et al. [16] showed that the respective ΔdksA mutant strains were complementable, and thus, the discrepancy is unlikely a polar effect. Additionally, in trans expression of DksA1 was sufficient to compensate for lack of DksA, indicating that the truncation did not impact the essential function of DksA. Lastly, expression of DksA enhanced and rescued intracellular growth in Lp02 and ΔpsrA strains, respectively. Thus, DksA can compensate for the lack of PsrA, and PsrA can partially compensate for the reduced activity of DksA1 but cannot compensate for the absence of DksA intracellularly in A. castellanii. Taken together, the interplay of DksA and PsrA in regulating their gene targets is not fully reciprocal, indicating that successful intracellular growth in A. castellanii is largely driven by DksA with assistance from PsrA.
The requirement of DksA for intracellular growth is host-specific. DksA was shown elsewhere to be dispensable for growth in primary murine macrophages [16] and for human U937-derived human macrophages as shown in this study (Fig. 3). Likewise, the ΔdksA ΔpsrA pThy strain was not impeded for intracellular growth in U937-derived macrophages, a finding similar to that previously found with the ΔpsrA strain [34]. In trans expression of DksA in the Lp02 strain background enhanced intracellular growth in protozoa (Fig. 2e), but with little overall impact in U937-derived macrophages (Fig. 3b), a trend also seen with overexpression of PsrA [34]. This suggests that elevated levels of either PsrA or DksA on an individual basis imbued a positive effect on replication in A. castellanii, which does not equate to a substantial phenotype in U937-derived macrophages.
To further delineate the distinct and overlapping roles of DksA and PsrA, in vitro approaches were pursued to examine the growth kinetics and phenotype characteristics of TP-associated traits of L. pneumophila. The induction of regulatory mechanisms controlling the transition switch from RP to TP is generally comparably induced in vivo (e.g. in protozoa) and in vitro [68]. In this view, in vitro kinetic assays were undertaken to show that the interactions between DksA and PsrA were largely growth temperature-independent (Figs45). Interestingly, in trans expression of DksA1 in the ΔdksA strain affected the reduced growth rate (Fig. 4c) at 37 °C, but not at 25 °C (Fig. 5d), illustrating that the predicted compromised transcriptional efficiency of DksA1 affects growth rates at higher temperatures. This is further supported by the growth profile of ΔdksA pVect-dksA1, which resembled that of Lp02 in A. castellanii (Fig. 2c). The slow in vitro growth phenotype did not apply to the Lp02 pVect-dksA1 strain bearing chromosomally expressed DksA, inferring that dksA1 is not a dominant-negative allele (Fig. 4F). The growth profiles of in vitro grown dksA1 pThy and dksA1 pVect-psrA were similar to each other at both at 25 °C (Fig. 4a, b) and 37 °C (Fig. 5a, b), but not in protozoa, where a 100-fold recovery was achieved through in trans expression of PsrA, inferring shared gene targets.
Other TP-associated phenotypes including cellular filamentation, culturability and pigmentation are mediated by DksA independently, as previously reported [16]. Lack of DksA resulted in unusual colony morphology (Fig. 6) and filamentation, particularly in SP (Figs78). Recently, DksA was ascertained to have a pivotal role in cell division in E. coli. The binding of ppGpp to DksA/RNAP indirectly activates the divisome complex component FtsZ to modulate cell length and division [69]. The absence of either DksA or ppGpp promotes cell filamentation for different reasons. Lack of DksA prevents activation of cell division by ppGpp, whereas lack of ppGpp causes DksA to switch from an activator to an inhibitor of FtsZ. Thus, by orthologous function, it is likely that DksA behaves in the same manner in L. pneumophila. The dramatically reduced bacterial culturability of the ΔdksA pThy strain (Fig. 9) demonstrates the inability of the strain to physiologically respond to nutrient deprivation and other stressors, which is consistent with the importance of DksA to the stringent response. Biologically, PsrA does not contribute to this phenotype as demonstrated by transcriptomic analyses that showed few stress-related genes under PsrA regulation [34]. Because variability in filamentation and culturability affected by lack of DksA can impact OD to titre ratios of SP bacteria, extreme care was taken in this study to ensure that ΔdksA-based strains were recovered from ≤3-day-old BCYE agar plates to minimize this variability for in vitro growth curves and inoculum titre for in vivo kinetics. Lastly, pigmentation comprises a brown tyrosine-based metabolite produced post-exponentially by L. pneumophila as an adaptation to environmental stress, potentially serving for photoprotection, iron acquisition or as a biocide to enhance L. pneumophila competitiveness in biofilms [7072]. As reported elsewhere, DksA is a major driver of pigment production (Fig. 10) [16]. Thus, these traits, other than cell length, do not seem measurably impacted by the cellular levels of PsrA, indicating that PsrA may impact virulence-specific pathways rather than cellular physiology.
Another phenotypic trait associated with TP differentiation is fatty acid flux, which is linked through SpoT-mediated stringent induction, with either DksA or ppGpp, sufficient to arrest growth upon supplementation with SCFA [1516]. Here, we show that, like its orthologue in Pseudomonas spp., PsrA is responsive to fatty acids, suggesting a conserved functional role in sensing LCFA in the environment alongside SpoT. Further, both DksA and PsrA must be present to ensure a proper LCFA response (Fig. 11) [63]. However, the precise mechanistic details of the PsrA-mediated response remain undefined, warranting further investigations.
In summary, this study explored the phenotypic roles of spontaneously mutated DksA1 and PsrA and uncovered a synergistic genetic relationship between PsrA and DksA in terms of virulence regulation. The bacterial response to LCFA suggests a novel regulatory link to the stringent response pathway, further solidifying the mechanistic importance of both DksA and PsrA as contributing regulators of the biphasic lifestyle of L. pneumophila. Because only the predicted, rather than the functional, basis of the dksA1 phenotype has been derived, these results cannot distinguish the distinct effects of ppGpp recruitment and competitive sequestration of RNAP from GreA. With these limitations in mind, our findings suggest that PsrA seems to redundantly act as a bypass for DksA for intracellular growth within A. castellanii protozoa but has little impact on other processes, suggesting a more targeted mechanism. The overlapping phenotypes presented in this study are consistent with the two distinct mechanisms by which DksA and PsrA act, and thus, the following model (Fig. 12) is proposed based on a scenario in which a promoter is repressed by both RNAP-bound DksA and PsrA (Fig. 12a). Absence of DksA would permit GreA to act as the sole secondary channel binding protein, thereby allowing increased efficiency in transcription initiation, and should there be elevated levels of PsrA, it would result in increased promoter blockage providing compensatory repression (Fig. 12b). Absence of PsrA would leave the promoter more accessible for RNAP, but this could be counteracted should there be elevated levels of DksA resulting in reduced transcriptional efficiency (Fig. 12c). Alternatively, a promoter not subject to PsrA-mediated influence would be exempt from the compensatory effects and, thus, be wholly dependent on the dynamics between GreA and DksA (Fig. 12d). Nevertheless, the proposed co-regulator duality could explain why transcriptomic analyses did not overtly reveal a large number of PsrA-regulated targets [34]. Thus, future transcriptomic studies will include the ΔdksA ΔpsrA strain to compare against data obtained for individual ΔpsrA and ΔdksA strains to discern distinct and overlapping roles. These studies could also be expanded to include the ppGpp^0^ (ΔrelA ΔspoT) strain to further explore the interface with PsrA and ppGpp in relation to the link between PsrA and the fatty acid response.
Model of independent mechanisms of DksA and PsrA. A hypothetical promoter is repressed by both PsrA and DksA. (a) RNAP holoenzyme is targeted to −10 and −35 boxes by a sigma factor (dark grey RNAP component). PsrA (oval dimers) binds a defined site in the promoter and inhibits RNAP recruitment. (b) In a ΔdksA strain, only GreA (triangles) is available for RNAP, thus elevating transcription levels, but in trans psrA overexpression generates higher levels of PsrA, strengthening inhibition of RNAP recruitment to compensate. Conversely, (c) the ΔpsrA strain allows uninhibited RNAP-promoter binding, increasing transcription, but this is offset by overexpressing dksA in trans, increasing the proportion of RNAP bound to transcription-inhibiting DksA (octagons), thereby generating the compensatory effect. (d) In an alternative scenario in which a promoter is not subject to PsrA influence, RNAP binds to GreA or DksA interchangeably, with enhanced transcription initiation rates occurring whilst bound to GreA versus DksA.
Supplementary material
10.1099/mic.0.001551Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Rowbotham TJ Current views on the relationships between amoebae, legionellae and man Isr J Med Sci 1986226786893793451 · pubmed ↗
- 2Steinert M Hentschel U Hacker J Legionella pneumophila: an aquatic microbe goes astray FEMS Microbiol Rev 20022614916210.1111/j.1574-6976.2002.tb 00607.x 12069880 · doi ↗ · pubmed ↗
- 3Newton HJ Ang DKY van Driel IR Hartland EL Molecular pathogenesis of infections caused by Legionella pneumophila Clin Microbiol Rev 20102327429810.1128/CMR.00052-0920375353 PMC 2863363 · doi ↗ · pubmed ↗
- 4Oliva G Sahr T Buchrieser C The life cycle of L. pneumophila: ecllular differentiation Is linked to virulence and metabolism Front Cell Infect Microbiol 20188310.3389/fcimb.2018.0000329404281 PMC 5780407 · doi ↗ · pubmed ↗
- 5Mondino S Schmidt S Buchrieser C Molecular mimicry: a paradigm of host-microbe coevolution Illustrated by Legionellam Bio 202011 e 01201-2010.1128/m Bio.01201-2033024033 PMC 7542358 · doi ↗ · pubmed ↗
- 6Finsel I Hilbi H Formation of a pathogen vacuole according to Legionella pneumophila: how to kill one bird with many stones Cell Microbiol 20151793595010.1111/cmi.1245025903720 · doi ↗ · pubmed ↗
- 7Burstein D Amaro F Zusman T Lifshitz Z Cohen O et al Genomic analysis of 38 Legionella species identifies large and diverse effector repertoires Nat Genet 20164816717510.1038/ng.348126752266 PMC 5050043 · doi ↗ · pubmed ↗
- 8Gomez-Valero L Rusniok C Carson D Mondino S Pérez-Cobas AE et al More than 18,000 effectors in the Legionella genus genome provide multiple, independent combinations for replication in human cells Proc Natl Acad Sci U S A 20191162265227310.1073/pnas.180801611630659146 PMC 6369783 · doi ↗ · pubmed ↗