Staphylococcus lugdunensis does not exert competitive exclusion on human corneocytes
Tianqi Zhang, Ran Luo, Marcus Ehrström, Keira Melican

TL;DR
This study investigates how Staphylococcus lugdunensis interacts with other bacteria on human skin and finds it does not block other bacteria from attaching.
Contribution
The study reveals that S. lugdunensis does not exhibit competitive exclusion on human corneocytes despite its antibiotic properties.
Findings
S. lugdunensis attaches to human skin stratum corneum but not in a way that prevents other bacteria from attaching.
S. lugdunensis is an efficient long-term colonizer despite reduced initial attachment.
The attachment pattern of S. lugdunensis does not overlap significantly with S. aureus or S. epidermidis.
Abstract
Human skin is our primary physical barrier and largest immune organ, and it also hosts a protective microbiota. Staphylococci are prominent members of the skin microbiota, including the ubiquitous coagulase-negative staphylococci (CoNS). The coagulase-positive Staphylococcus aureus is found as part of the microbiota, but it poses clinical concern due to its potential pathogenicity and antibiotic resistance. Recently, a CoNS, Staphylococcus lugdunensis, has been shown to inhibit S. aureus growth via the production of a novel antibiotic, lugdunin. In this study, we use human skin models to understand the spatial relationships between the CoNS Staphylococcus epidermidis and S. lugdunensis with S. aureus during colonization of human skin. We investigated the attachment patterns of the bacteria, both individually and in competition. Surprisingly, we found that attachment did not always…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
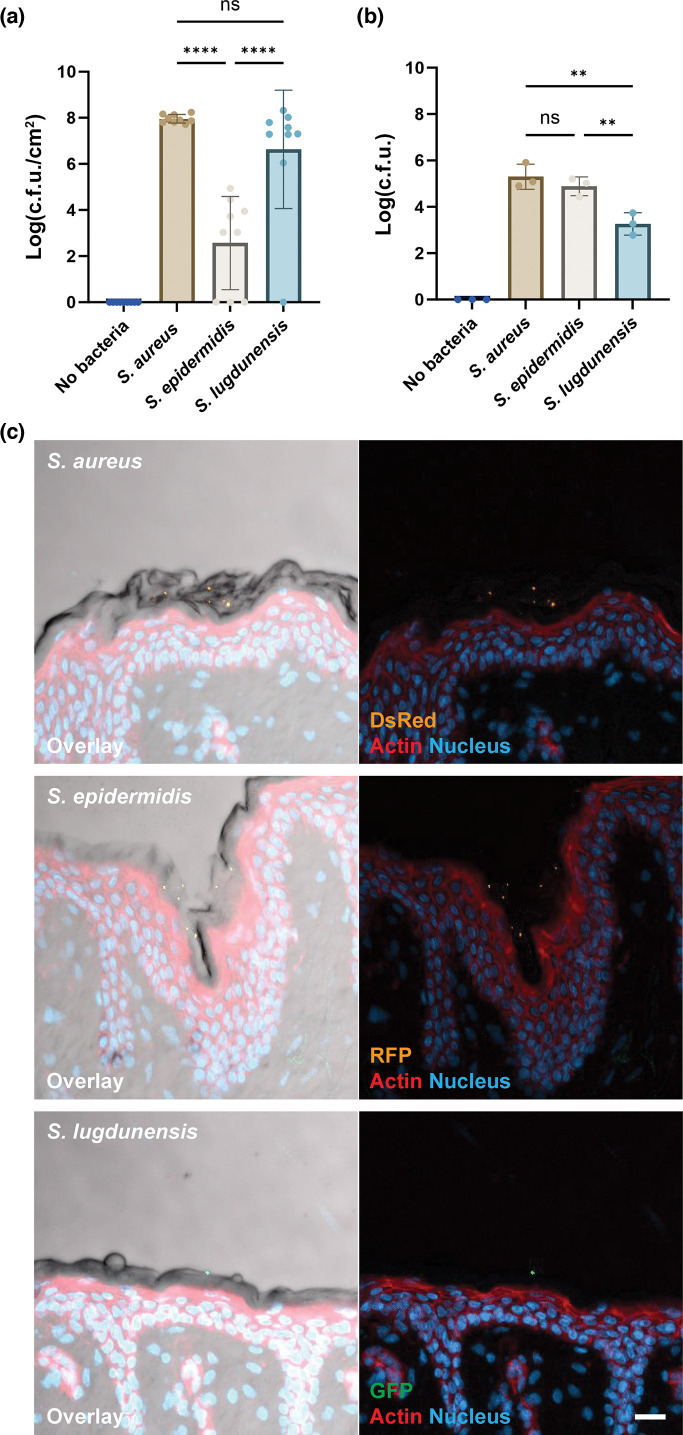 Fig. 1
Fig. 1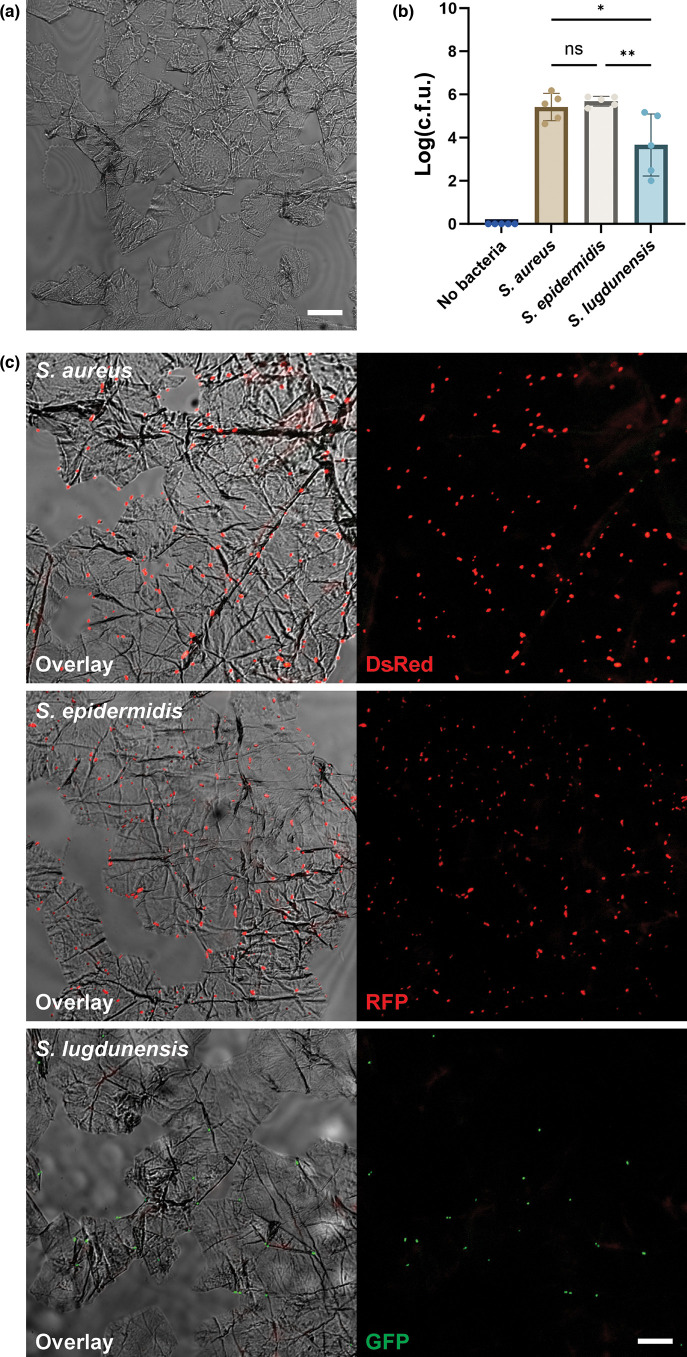 Fig. 2
Fig. 2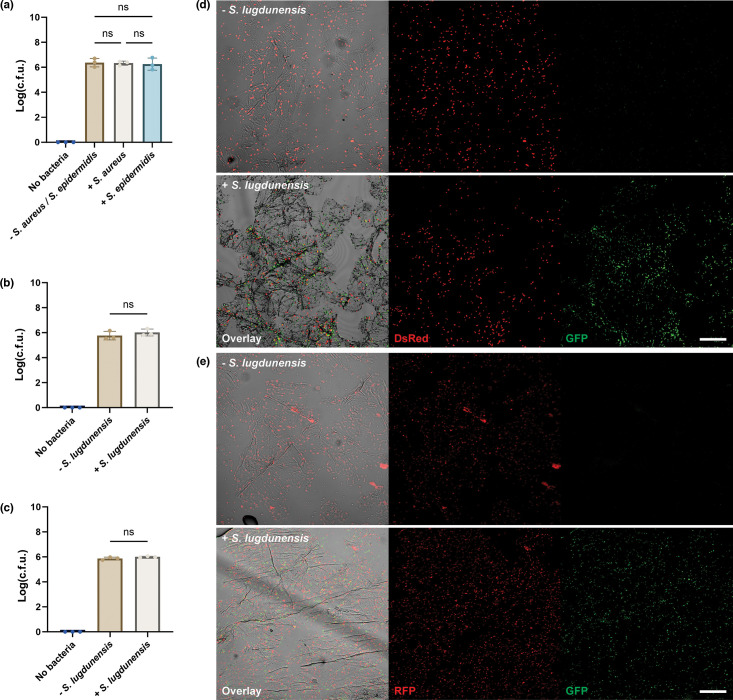 Fig. 3
Fig. 3| Strains | Description | Selective antibiotics | Source |
|---|---|---|---|
|
| AH3849 - USA300 CA-MRSA, Erms (LAC*) (AH1263) pHC48 (DsRed) | Chloramphenicol 25 µg ml−1 | Ibberson et al. (2016) [ |
|
| Trimethoprim 10 µg ml−1 | Mack et al. (1992) [ | |
|
| Erythromycin 5 µg ml−1 | Flannagan et al. (2018) [ | |
|
| |||
| pSRFPS1 | Vector for RFP | Trimethoprim | BEI Resources NR-51164 |
- —http://dx.doi.org/10.13039/501100004359 Vetenskapsrådet
- —http://dx.doi.org/10.13039/501100009708 Novo Nordisk Fonden
- —http://dx.doi.org/10.13039/501100004047 Karolinska Institutet
- —http://dx.doi.org/10.13039/501100004359 Vetenskapsrådet
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDermatology and Skin Diseases · Nail Diseases and Treatments · Contact Dermatitis and Allergies
Introduction
The skin is the largest human organ and serves as one of the body’s first-line defences against invasive pathogens [14]. A key feature of this barrier is the outermost layer of the epidermis, the stratum corneum, composed of anuclear, terminally differentiated corneocytes and intercellular lipids [45]. Human skin is home to a diverse bacterial microbiota believed to actively contribute to skin defence by competitive exclusion of potential pathogens and by educating the local innate immune response [13,6]. The staphylococci are one of the most abundant bacterial species of the skin microbiota [6]. The coagulase-negative (CONs) Staphylococcus epidermidis is found across multiple skin sites [7], while the clinically important pathogen Staphylococcus aureus is referred to as a ‘pathobiont’, a symbiont with significant pathogenic potential [689].
To successfully colonize human skin, bacteria must first attach. Previous work with human skin models in our lab highlighted the outer stratum corneum as a primary colonization site for S. aureus on human skin [1011]. Other labs have shown that cell wall-anchored (CWA) protein accumulation-associated protein (Aap) from S. epidermidis and the orthologous surface protein G (SasG) from S. aureus are important to attachment to healthy human corneocytes [1214]. As is often the case for staphylococci, there appears to be redundancy in this process. Many S. aureus strains, including JE2, appear to possess premature stop codons in the SasG sequence, suggesting that the presence of other binding mechanisms is also likely [14]. CWA clumping factor B (ClfB) and fibronectin-binding protein B (FnBPB) have been implicated in facilitating S. aureus colonization of pathologic atopic dermatitis (AD) skin [1517]. However, as AD skin has very different characteristics, whether similar mechanisms are present in healthy human skin remains to be studied.
Research has investigated differences between the potentially pathogenic S. aureus and commensal CoNS often using the prototypical CoNS S. epidermidis. More recently, other CoNS have been gaining attention for their role in protecting against pathogens. S. lugdunensis has garnered particular consideration following the discovery of its production of a novel antibiotic, lugdunin [18]. S. lugdunensis is however far from harmless, it causes skin and soft tissue infections and a particularly aggressive form of infective endocarditis [1920]. Despite being considered a skin commensal, little is known about the mechanism behind S. lugdunensis attachment to human skin. CWA Von Willebrand factor-binding protein (vWbl) and fibrinogen-binding protein (Fbl) have been proposed to facilitate S. lugdunensis attachment to the extracellular matrix during endothelial invasion and endovascular infections [21], and the expression of autolysin AtlL promoted S. lugdunensis adherence to epithelial and endothelial cells [22]. For skin attachment, however, a specific adhesion has not been described, and S. lugdunensis does not encode ClfB and SdrD, nor any orthologue to the S. epidermidis Aap [212324].
In this work, we compared the colonization and attachment of S. aureus, S. epidermidis and S. lugdunensis in a human skin biopsy model and confirmed the outer stratum corneum as the primary binding site for the selected strains. We then used a corneocyte adhesion assay and optimized it for a comparison of competitive exclusion. We investigated how staphylococci differentially attach to the outer stratum corneum and whether any potential space exclusion occurs between the strains. We show that S. lugdunensis has a unique binding pattern compared to S. aureus and S. epidermidis and shows a limited role in competitive exclusion on human corneocytes.
Methods
Bacterial strains and culture conditions
The bacterial strains used are listed in Table 1. Strains were grown in Tryptic soy broth (TSB) (Sigma-Aldrich) with indicated antibiotic, at 37 °C with 180 r.p.m. agitation. For log-phase suspension, an overnight culture was diluted 1 : 100 in TSB with appropriate antibiotic and incubated at 37 °C 180 r.p.m. to OD600 ~0.6. The culture was washed twice with PBS using centrifugation (4000 r.p.m., 10 min) and re-suspended PBS to the required OD600.
Ex vivo human skin model (biopsy model)
After collection, the skin surface was sterilized with 70% ethanol ×3. Full-thickness ex vivo human skin biopsy was collected with an Acuderm Inc. Biopsy Punch 10 mm (Thermo Fisher Scientific). The biopsy was immersed in 70% EtOH for 10 s and washed with PBS, pH 7.4 (Gibco™ Thermo Fisher Scientific). After drying, a hydrocoll (Kontorsgiganten) hydrogel ring was placed on each biopsy to seal the edges. Biopsies were placed in a TC insert (Sarstedt) and each insert into a Corning® Costar® TC-Treated 12-Well Plate (Sigma-Aldrich). Six hundred fifty microlitres of CO_2_ Independent Medium (Thermo Fisher Scientific) supplemented with 1X CTS™ GlutaMAX™-I Supplement (Thermo Fisher Scientific) and 10% foetal bovine serum (Sigma-Aldrich) (culture medium) were added under the biopsies, with the skin surface remaining in air. The biopsies were incubated at 37 °C overnight. Colonization inoculum was prepared by diluting a log-phase bacterial suspension with PBS to an OD600 1.95 (~ 6.5×10^8^ c.f.u. ml^−1^). Two microlitres of the colonization inoculum were evenly distributed across the biopsy surface. The biopsy was incubated at 37 °C for 48 h with media changes every 24 h. After incubation, the hydrogel ring was removed, and the biopsy was collected for subsequent analysis.
Tape-stripped human skin stratum corneum model
The surface of the skin biopsy was taped once with Office Tape 31-3278 (Clas Ohlson) to remove debris. To isolate stratum corneum, dermatology tape d-Squame Standard 1up Label (Clinical and Derm) was applied to the skin surface with consistent pressure for 5 s. The tape was gently removed and placed cell-side up in a Corning® Costar® TC-Treated 6-Well Plate (6-well plate) (Sigma-Aldrich).
Bacterial attachment assays
For attachment assays, the inoculum was diluted from log-phase suspension with PBS to OD600 0.15 (~ 5×10^7^ c.f.u. ml^−1^). For the competition assay, attachment inoculum was prepared to an OD600 1.5 (~ 5×10^8^ c.f.u. ml^−1^) for S. lugdunensis and OD 0.15 for * S. epidermidis* or S. aureus. Three hundred microlitres of inoculum were placed in each well of a 12-well plate. The biopsy was placed onto the drop with the outer skin surface facing downward. For the tape model attachment assay, 300 µl of inoculum was added to the centre of each tape. After 45 min at 37 °C, unbound bacteria were removed by washing extensively with PBS. For the biopsy model, the skin surface was washed with 10 ml PBS, and the bottom was washed with 2 ml PBS. For the tape model, the cell side was washed with 10 ml PBS, and the backside was washed with 2 ml PBS.
c.f.u. quantification
Biopsies or tapes were homogenized using Lysing Matrix D (MP Biomedicals) tube with 1 ml PBS. Tubes were homogenized at 6.5 m/s for 20 s ×3, with 5-min interval on ice. The homogenate was serial diluted and plated on a Tryptic soy agar (TSA) (Sigma-Aldrich) containing selective antibiotics as outlined in Table 1 and incubated at 37 °C overnight. To facilitate analysis, a c.f.u.=0 is transformed to 1, which represents the limit of detection of this model. To stabilize variance, the c.f.u. numbers were transformed into logarithmic base 10 values (log-transformation) and plotted using Prism 10 software (GraphPad). Statistical comparisons between the log-transformed results were conducted with Prism 10 using unpaired t-tests for comparisons between two groups and ordinary one-way ANOVA with Tukey correction for comparisons involving more than two groups.
Imaging analysis
Biopsies were placed in an Epredia™ Disposable Embedding Mold (Fisher Scientific) and embedded in CellPath OCT Embedding Matrix (Fisher Scientific). The embedded biopsy was flash-frozen and stored at −80 °C. Prior to staining, the OCT-embedded biopsy was sectioned 20 µm, mounted on VWR® SuperFrost® Plus slides (VWR) and fixed with 100% methanol (Sigma-Aldrich) on ice for 10 min. Immunofluorescence staining utilizing the following antibodies: NucBlue™ Fixed Cell ReadyProbes™ Reagent (DAPI) (R37606, Thermo Fisher Scientific) and Alexa Fluor™ 647 Phalloidin (A22287, Thermo Fisher Scientific). Primary antibodies were incubated at 4 °C overnight, while secondary antibodies, Alexa Fluor™ 700 (A-21038, Thermo Fisher Scientific), were incubated at room temperature for 1 h. Following staining, the sample was mounted against 50×24 mm VWR® Micro Cover Glasses (50×24 cover glass) (VWR) using Dako Fluorescence Mounting Medium (Agilent) and stored overnight at 4 °C before imaging.
Processing and staining of the tape
For visualization of bacterial attachment to the tape model, each tape was directly mounted onto a 50×24 mm cover glass and sealed against a microscopy slide (VWR). Following fixation, the cell side of the tape was stained using the stains listed above. Once stained, the tape was fitted and mounted in µ-Dish 35 mm, high (Ibidi) using Dako Fluorescence Mounting Medium. All mounted samples were imaged immediately.
Microscopy
Samples were imaged using an LSM 700 Laser Scanning Microscope (Zeiss) with a 20× air objective. Post-acquisition, images underwent minor adjustments to brightness and contrast, uniformly applied to the entire image and digitally enlarged using Fiji software (ImageJ, National Institutes of Health, USA).
Results
Different Staphylococcus strains colonize and adhere differently to human skin
As an extension to our previous work on the human skin colonization patterns of S. epidermidis vs. S. aureus [11], we included here S. lugdunensis. We inoculated ex vivo human skin biopsies with 1.3×10^6^ c.f.u. of S. aureus, S. epidermidis or S. lugdunensis and incubated at 37 °C for 48 h. After 48 h, S. aureus and S. lugdunensis grew on the skin, colonizing the surface, with S. aureus showing a mean log_10_ c.f.u. cm^−2^ of 8.0 and S. lugdunensis 6.6 (Fig. 1a). S. epidermidis demonstrated significant donor-to-donor variation in its colonization, something we have noted in previous work [11], with a significantly lower mean log_10_ c.f.u. cm^−2^ of 2.6 (Fig. 1a). These data showed a variance between the colonization properties of the different bacterial strains on the skin biopsy model at 48 h, suggesting S. lugdunensis colonized more like S. aureus than S. epidermidis. The first step in colonization is bacterial attachment. We, therefore, sought to quantify bacterial attachment to the human skin biopsy. Using methodology based on previous assays [1214], we adhered the bacteria to the skin surface for 45 min before extensive washing. Interestingly, the results of the adhesion assay differed from colonization. S. lugdunensis demonstrated the lowest level of adhesion with a mean of 3.3 log_10_ c.f.u., significantly lower than both S. aureus (5.3) and S. epidermidis (4.9) (Fig. 1b). Individual donor variation was again evident. S. lugdunensis showed a 100-fold reduction in adhesion compared to S. aureus and a 40-fold decrease compared to S. epidermidis (Fig. 1b). Microscopy analysis showed that S. aureus, S. epidermidis and S. lugdunensis all adhered primarily to the stratum corneum (Fig. 1c), consistent with our previous work [1011]. These results suggest an apparent discrepancy in staphylococcal adhesion vs. colonization on human skin. S. aureus showed a strong ability to attach and colonize the human skin biopsy models, while S. epidermidis was able to attach but was slower to colonize. S. lugdunensis had a lower ability to attach but colonized the skin to a comparable level as S. aureus.
*Staphylococci colonization and attachment to human skin biopsy. (a) Colonization of human skin at 37 °C for 48 h. Mean log transformed c.f.u. per cm2 with sd. n=9 from three individual skin donors. Statistical significance was determined by one-way ANOVA. Statistical significance is denoted by ns (not significant) or ****P≤0.0001. (b–c) Attachment of the selected strains at 37 °C for 45 min. (b) Mean log-transformed total c.f.u. is shown. Combination of n=3 from three individual donors. Statistical significance was determined by one-way ANOVA. Statistical significance is denoted by ns (not significant) or *P≤0.01. (c) Fluorescence imaging of the location of bacterial attachment on human skin. DsRed-expressing S. aureus (AH3849), RFP-expressing S. epidermidis (1457-pSRFPS1) and GFP-expressing S. lugdunensis (S. lugdunensis-GFP) are shown. Overlay image is a combination of brightfield, DsRed channel (DsRed or RFP) in orange, GFP channel (GFP) in green, Alexa 647 channel (actin) in red and DAPI channel (nucleus) in blue. Representative images of n=3. Scale bar=20 µm.
Staphylococcal attachment to isolated corneocytes
To assess the direct role of the stratum corneum in bacterial adhesion, we focused on bacterial attachment to primary corneocytes. We adopted a previously used approach, using tape-stripped human skin to isolate the stratum corneum [2526] (Fig. 2a). We repeated the attachment assay using freshly isolated tape-stripped corneocytes. Forty-five minutes after exposure of corneocytes to bacteria, the cells were washed with PBS to remove any non-adherent bacteria. Bacterial adhesion to isolated corneocytes reflected what we had seen in the full-thickness biopsy (Fig. 2b). S. aureus (5.4 mean log_10_ c.f.u.) and S. epidermidis (5.7) adhered to a similar degree, while S. lugdunensis again demonstrated significantly lower adhesion (3.7 log_10_ c.f.u.) (Fig. 2b). To visualize the binding patterns of the staphylococci on corneocytes, we performed fluorescence imaging. Microscopy of uninfected tape-stripped cells confirmed that the majority of cells isolated by our model are anuclear skin corneocytes (Fig. 2a). Microscopy revealed a dense presence of S. aureus and S. epidermidis on the corneocytes even after extensive washing (Fig. 2c). In contrast, S. lugdunensis showed considerably less binding. Interestingly, most of the staphylococci found in this model were adhering at the edges of the corneocytes, at the cell–cell borders (Fig. 2c). Together, these data showed similarity in bacteria attachment patterns to isolated corneocytes compared to the full-thickness biopsy. The binding location of most bacteria at the cell–cell junctions of the corneocytes highlighted potential binding preference towards binding targets present at the edges of the corneocytes.
*Staphylococci attachment to isolated corneocytes. (a) Brightfield imaging of uninfected tape-stripped stratum corneum. Representative of n=3 from three donors. Scale bar=20 µm. (b–c) Attachment of the selected strains at 37 °C for 45 min. (b) Mean log-transformed total c.f.u. Combination of n=5 from five donors. Statistical significance was determined by one-way ANOVA. Statistical significance is denoted by ns (not significant), *P≤0.05 or *P≤0.01. (c) Fluorescence imaging of the attachment. DsRed-expressing S. aureus (AH3849), RFP-expressing S. epidermidis (1457-pSRFPS1) and GFP-expressing S. lugdunensis (S. lugdunensis-GFP) were shown. The overlay image was generated in combination with the brightfield channel, DsRed channel (DsRed or RFP) in red and GFP channel (GFP) in green. Representative images of n=3 from three donors. Scale bar=20 µm.
Limited competitive exclusion on human corneocytes
Little is known regarding the binding mechanism of S. lugdunensis to human skin. Previous studies have shown that CWA proteins such as accumulation-associated protein (Aap) from S. epidermidis and its orthologous surface protein G (SasG) from S. aureus are critical to staphylococci attachment to corneocytes [25]. However, no orthologue to Aap or SasG has yet been found in S. lugdunensis [1923]. We, therefore, sought to understand if there may be a functional orthologue in S. lugdunensis by designing a competitive exclusion assay. We co-attached S. aureus or S. epidermidis together with a tenfold increased concentration of S. lugdunensis. To start, we increased the inoculum of S. lugdunensis to OD600 1.5 (~ 5×10^8^ c.f.u. ml^−1^) (Fig. 3a). The presence of neither S. aureus nor S. epidermidis in competition altered the adhesion numbers of S. lugdunensis (Fig. 3a). Comparably, the large S. lugdunensis volume did not alter the adhesion capability of S. aureus with mean log_10_ c.f.u. of 5.9 when attaching individually (Fig. 2b) and 6.0 when in competition with S. lugdunensis (Fig. 3b). No statistical difference was seen between the two conditions, suggesting limited competitive exclusion. Similar results were seen for competition between S. epidermidis and S. lugdunensis. S. epidermidis showed a mean log_10_ c.f.u. of 5.8 while attaching alone and 6.0 while attaching in competition with S. lugdunensis. No statistical difference was found between the two conditions (Fig. 3c). Microscopy of the competition assay confirmed the quantification (Fig. 3d, e). Similar amounts and patterns of * S. aureus* binding could be seen regardless of S. lugdunensis presence (Fig. 3d). This was also the case for S. epidermidis (Fig. 3d, e). Taken together, no competitive exclusion was found between S. lugdunensis and S. aureus or S. epidermidis in this model. This indicates that S. lugdunensis has a unique adhesion pattern on human corneocytes.
*Staphylococci competition on isolated corneocytes. (a) S. lugdunensis attachment was not affected by the presence of S. aureus or S. epidermidis. (b, c) The presence of S. lugdunensis did not affect the attachment of (b) S. aureus or (c) S. epidermidis. Mean log-transformed total c.f.u. is shown. n=3 from three donors. Statistical significance was determined by one-way ANOVA. Statistical significance denoted by ns (not significant), *P≤0.05 or *P≤0.01. (d, e) Fluorescence imaging the attachment of (d) S. aureus or (e) S. epidermidis without (top) and with (bottom) the presence of abundant S. lugdunensis. DsRed-expressing S. aureus (AH3849), RFP-expressing S. epidermidis (1457-pSRFPS1) and GFP-expressing S. lugdunensis (S. lugdunensis-GFP) are shown Overlay image is a combination of brightfield, DsRed channel (DsRed or RFP) in red and GFP channel (GFP) in green. Representative images of n=3 from three donors. Scale bar=50 µm.
Conclusions
Adhesion is often the first step of host–bacterial interaction. The understanding of how different bacteria adhere, bind and colonize different tissues is an ongoing research question and a promising target for novel antimicrobial therapies. The protective properties of the human microbiota are believed to involve competitive exclusion, where the resident microbiota outcompete potential pathogens for space and nutrients. In this work, we focused on the potential skin pathogen S. aureus and the interaction with both human skin and CoNS. It has become apparent in recent years that there is great variation among the CoNS, so we included the prototypical skin commensal S. epidermidis as well as the emerging potential probiotic strain S. lugdunensis. The colonization and adhesion patterns of S. aureus and S. epidermidis were similar to previous results from our own work as well as by others [10132627]. Particularly, we see significant donor-to-donor variation in S. epidermidis, which we believe relates to the background of the individual donors and their prior exposure to skin microbiota. While the use of human skin does introduce this variance, we believe it is more representative of the real-world situation. S. lugdunensis gave interesting results, showing that while it appears to colonize over 48 h at a similar extent to * S. aureus*, its adhesion at 45 min was significantly less. S. lugdunensis is an intriguing CoNS with very particular metabolic requirements, the best described being its iron-acquisition systems [2829]. It is the only CoNS to express an iron-regulated surface determinant system (Isd) and can use this high-affinity uptake system in low-haem environments such as the skin. Distinctively, the S. lugdunensis IsdB has a reduced affinity to murine haemoglobin compared to humans, indicating a human specificity of this bacteria [1928]. Our data appear to support this specificity, showing an unexpectedly good colonization rates on human skin for S. lugdunensis, showing a colonization pattern more similar to S. aureus than S. epidermidis. Emerging data from other labs as well as our own preliminary data suggest a significant interplay between S. lugdunensis and S. aureus. This included how S. lugdunensis produces the antibiotic lugdunin [18] as well as data showing that S. lugdunensis can hijack iron acquisition and nutritional immunity of S. aureus to facilitate growth [1930]. Our data support this exciting line of research by demonstrating for the first time that the adhesion patterns of * S. lugdunensis* are significantly different from both S. aureus and S. epidermidis on human skin. This would suggest that its inhibitory properties are not related to competitive exclusion. Our data show that S. lugdunensis has a unique binding pattern compared to the other strains that correspond to other data, showing that S. lugdunensis does not possess many of the known CWA proteins present in the other staphylococci such as ClfB, SdrD and Aap [19]. Further research is now needed to define this target. Lastly, our data provide information regarding the spatial localization of the different staphylococcal strains on human skin. This is important for the ongoing work into contact-dependent inhibition, quorum repression and iron acquisition as potential treatment targets. In conclusion, this work provides new knowledge on the binding targets and patterns of different staphylococcal strains on human skin. It opens several new avenues for research into the interactions between staphylococcal strains and how we can take advantage of the human skin microbiota as a means to prevent infection.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Byrd AL Belkaid Y Segre JA The human skin microbiome Nat Rev Microbiol 20181614315510.1038/nrmicro.2017.15729332945 · doi ↗ · pubmed ↗
- 2Grice EA Segre JA The skin microbiome Nat Rev Microbiol 2011924425310.1038/nrmicro 253721407241 PMC 3535073 · doi ↗ · pubmed ↗
- 3Grice EA Kong HH Conlan S Deming CB Davis J et al Topographical and temporal diversity of the human skin microbiome Science 20093241190119210.1126/science.117170019478181 PMC 2805064 · doi ↗ · pubmed ↗
- 4Kabashima K Honda T Ginhoux F Egawa G The immunological anatomy of the skin Nat Rev Immunol 201919193010.1038/s 41577-018-0084-530429578 · doi ↗ · pubmed ↗
- 5Elias PM Stratum corneum defensive functions: an integrated view J Invest Dermatol 200512518320010.1111/j.0022-202X.2005.23668.x 16098026 · doi ↗ · pubmed ↗
- 6Otto M Staphylococci in the human microbiome: the role of host and interbacterial interactions Curr Opin Microbiol 202053717710.1016/j.mib.2020.03.00332298967 · doi ↗ · pubmed ↗
- 7Byrd AL Belkaid Y Segre JA The human skin microbiome Nat Rev Microbiol 20181614315510.1038/nrmicro.2017.15729332945 · doi ↗ · pubmed ↗
- 8Jochum L Stecher B Label or concept - What Is a Pathobiont?Trends Microbiol 20202878979210.1016/j.tim.2020.04.01132376073 · doi ↗ · pubmed ↗