Metatranscriptomic detection of rabbit hemorrhagic disease virus 2 in karoro (southern black-backed gulls)
Stephanie J. Waller, Chris N. Niebuhr, Jessica A. Darnley, Kate McInnes, David Winter, Edward C. Holmes, Jemma L. Geoghegan

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
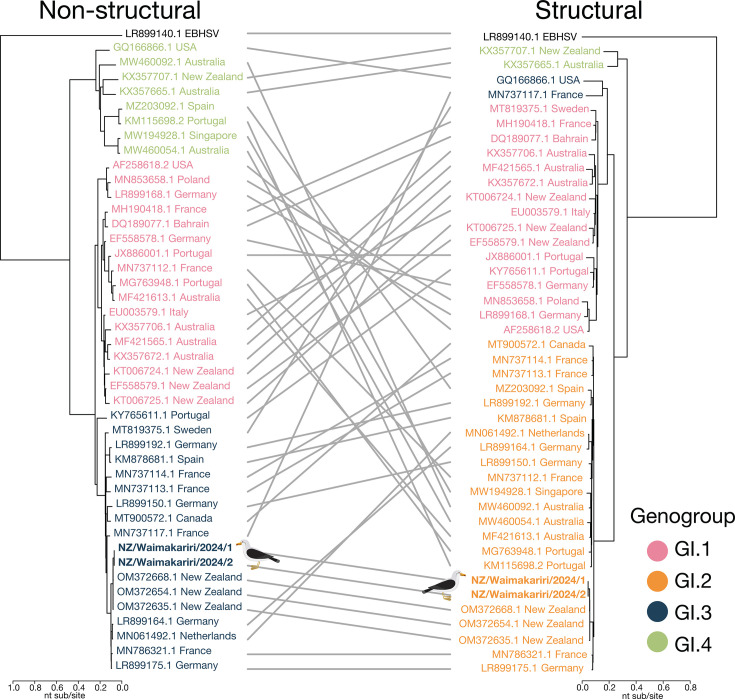 Fig 1
Fig 1- —Royal Society of New Zealandhttp://dx.doi.org/10.13039/501100001509
- —Ministry of Business, Innovation and Employmenthttp://dx.doi.org/10.13039/501100003524
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Virus Infections Studies · Viral Infections and Vectors · Viral gastroenteritis research and epidemiology
LETTER
Rabbit hemorrhagic disease virus 2 (RHDV2; species Lagovirus europaeus) is a highly pathogenic lagovirus (Caliciviridae) responsible for a lethal disease in rabbits and hares (1). First identified in Europe in 2010 (2), RHDV2 has spread to over 35 countries (3). RHDV2 is believed to have arrived in New Zealand’s North Island in 2016 and has become established in both wild and domestic rabbit populations (3). In contrast, rabbit hemorrhagic disease virus 1 has been present in New Zealand since its illegal release as a biological control agent in 1997 (4).
The RHDV2 strain circulating in New Zealand belongs to the GI.3P-GI.2 variant—a recombinant of the non-structural protein of a benign GI.3P lagovirus with the structural protein of GI.2 RHDV2 (3, 5) (Fig. 1). This variant has been detected in Europe (3, 5–7), North America (3, 8, 9), and China (3, 10), but notably not in Australia where RHDV2 has circulated since 2014 (3, 11). This suggests that the New Zealand incursion likely originated from outside Australia, although its precise source remains unknown (3).
Tanglegram of New Zealand and representative global lagovirus sequences highlighting the recombinant viral genome. Representative lagovirus nucleotide sequences were obtained from NCBI/GenBank and aligned with the New Zealand RHDV2 sequences (bold) using MAFFT (12). Maximum likelihood phylogenetic trees were inferred for the non-structural (left) and structural genes (right) (nucleotide positions 28–5304 and 5305–7382, respectively [3]) using IQTREE version 1.6.12 (13), with the best-fit model as determined by ModelFinder (14). Phylogenies were rooted using European brown hare syndrome virus (EBHSV) (LR899140.1). A tanglegram was created using the ape (15) and dendextend (16) packages with the tanglegram function in RStudio version 4.3.1. Sequences are colored by genogroup, and scale bars indicate the number of nucleotide substitutions per site.
Since its introduction, RHDV2 has spread within New Zealand, including at least two transmission events from the North Island to the South Island (3). The Cook Strait, which separates the islands, is approximately 23 km wide at its narrowest point. Although the exact mechanism of inter-island transmission is unknown, potential pathways include human-mediated movement, contaminated fomites, wind-assisted dispersal of fly vectors, or scavenging birds (3, 17, 18).
Through metatranscriptomic sequencing, we recovered one complete and one near-complete RHDV2 GI.3P-GI.2 genome (PV602081 and PV602082) from two RNA pools derived from oral and cloacal swabs collected in 2024 from eight southern black-backed gulls (karoro, Larus dominicanus) on the South Island. Oral and cloacal swabs were stored separately in DNA/RNA Shield (Zymo Research) immediately upon collection and subsequently at –80°C until total RNA was extracted using the ZymoBIOMICS MagBead RNA kit (Zymo Research). These viral sequences, with abundances of 30 (oral) and 1,159 (cloacal) reads per million, exhibited over 98% nucleotide sequence similarity in both structural and non-structural regions to the RHDV2 previously detected in European rabbits (Oryctolagus cuniculus) from the Otago region (South Island, New Zealand) in 2019 (3) (Fig. 1). Southern black-backed gulls are opportunistic feeders that consume a variety of terrestrial and marine organisms, as well as organic waste from farms and landfills (19). The presence of RHDV2 in these gulls is therefore likely dietary in origin, which is supported by the presence of rabbit reads in the metatranscriptomic data. In addition, RHDV2 is not believed to replicate in avian hosts.
The observation that RHDV2 is present in avian samples, albeit likely of dietary origin, makes it theoretically possible that birds act as mechanical vectors for virus transmission, perhaps through contaminated feces. This may even explain the spread of RHDV2 between New Zealand’s islands. While southern black-backed gulls are largely resident and not known for long-distance migration, there is strong evidence for their dispersal between the North and South Islands (20). The detection of complete RHDV2 genomes in these scavenger birds warrants further investigation of their potential role in viral dissemination. More broadly, these birds may influence the long-range spread of pathogens, shape viral dynamics, and present challenges for disease outbreak containment.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hall RN, King T, O’Connor T, Read AJ, Arrow J, Trought K, Duckworth J, Piper M, Strive T. 2021. Age and infectious dose significantly affect disease progression after RHDV 2 infection in naïve domestic rabbits. Viruses 13:1184. doi:10.3390/v 1306118434205750 PMC 8234499 · doi ↗ · pubmed ↗
- 2Rouco C, Aguayo-Adán JA, Santoro S, Abrantes J, Delibes-Mateos M. 2019. Worldwide rapid spread of the novel rabbit haemorrhagic disease virus (GI.2/RHDV 2/b). Transbound Emerg Dis 66:1762–1764. doi:10.1111/tbed.1318930924292 · doi ↗ · pubmed ↗
- 3Hall RN, Trought K, Strive T, Duckworth JA, Jenckel M. 2024. First detection and circulation of RHDV 2 in New Zealand. Viruses 16:519. doi:10.3390/v 1604051938675862 PMC 11053765 · doi ↗ · pubmed ↗
- 4O’Keefe JS, Tempero JE, Motha MX, Hansen MF, Atkinsona PH. 1999. Serology of rabbit haemorrhagic disease virus in wild rabbits before and after release of the virus in New Zealand. Vet Microbiol 66:29–40. doi:10.1016/s 0378-1135(98)00307-110223320 · doi ↗ · pubmed ↗
- 5Abrantes J, Droillard C, Lopes AM, Lemaitre E, Lucas P, Blanchard Y, Marchandeau S, Esteves PJ, Le Gall-Reculé G. 2020. Recombination at the emergence of the pathogenic rabbit haemorrhagic disease virus Lagovirus europaeus/GI.2. Sci Rep 10:14502. doi:10.1038/s 41598-020-71303-432879332 PMC 7468141 · doi ↗ · pubmed ↗
- 6Miao Q, Qi R, Veldkamp L, Ijzer J, Kik ML, Zhu J, Tang A, Dong D, Shi Y, van Oers MM, Liu G, Pijlman GP. 2019. Immunogenicity in rabbits of virus-like particles from a contemporary rabbit Haemorrhagic disease virus type 2 (GI.2/RHDV 2/b) isolated in the Netherlands. Viruses 11:553. doi:10.3390/v 1106055331207978 PMC 6631637 · doi ↗ · pubmed ↗
- 7Fitzner A, Kesy A, Bulenger K, Niedbalski W. 2021. Evidence of independent introductions of RHDV 2 strains in Poland based on the genome analysis of viral isolates from 2016–2018. Acta Biochim Pol 68:255–263. doi:10.18388/abp.2020_554733848411 · doi ↗ · pubmed ↗
- 8Ambagala A, Schwantje H, Laurendeau S, Snyman H, Joseph T, Pickering B, Hooper-Mc Grevy K, Babiuk S, Moffat E, Lamboo L, Lung O, Goolia M, Pinette M, Embury-Hyatt C. 2021. Incursions of rabbit haemorrhagic disease virus 2 in Canada—Clinical, molecular and epidemiological investigation. Transbound Emerg Dis 68:1711–1720. doi:10.1111/tbed.1412833915034 · doi ↗ · pubmed ↗