Late gene regulation by the alternative sigma factors of Chlamydia trachomatis
Syed M. A. Rizvi, Asha Densi, Owais R. Hakiem, Michael McClelland, Ming Tan

TL;DR
This study identifies how two alternative sigma factors in Chlamydia trachomatis regulate specific genes during the conversion of bacterial forms inside host cells.
Contribution
The study reveals that σ28 and σ54 sigma factors regulate only a small subset of late genes in Chlamydia, with distinct regulatory mechanisms.
Findings
σ28 binds to hctB and tsp genes only at late stages of Chlamydia's life cycle.
σ54 binds to ctl0021 and ctl0052, with conserved promoter sequences across Chlamydia species.
Euo regulates σ28 expression but not σ54, indicating multiple mechanisms for late gene regulation.
Abstract
The pathogenic bacterium Chlamydia reproduces via two specialized forms inside a eukaryotic host cell. The dividing form called the reticulate body (RB) must convert at late times into the infectious elementary body (EB) for spread to new host cells. Late genes are a temporal class of chlamydial genes believed to be responsible for RB-to-EB conversion, but late gene regulation is incompletely understood. In this study, we used chromatin immunoprecipitation (ChIP) to investigate two alternative sigma factors, σ28 and σ54, that alter the promoter specificity of Chlamydia trachomatis RNA polymerase. σ28 ChIP-seq identified hctB and tsp as the only promoters bound by σ28, and binding only occurred late, around the time of RB-to-EB conversion. Overexpression of σ28 confirmed that these genes are transcribed in a σ28-dependent manner. σ54 ChIP-seq showed that σ54 only bound ctl0021 and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
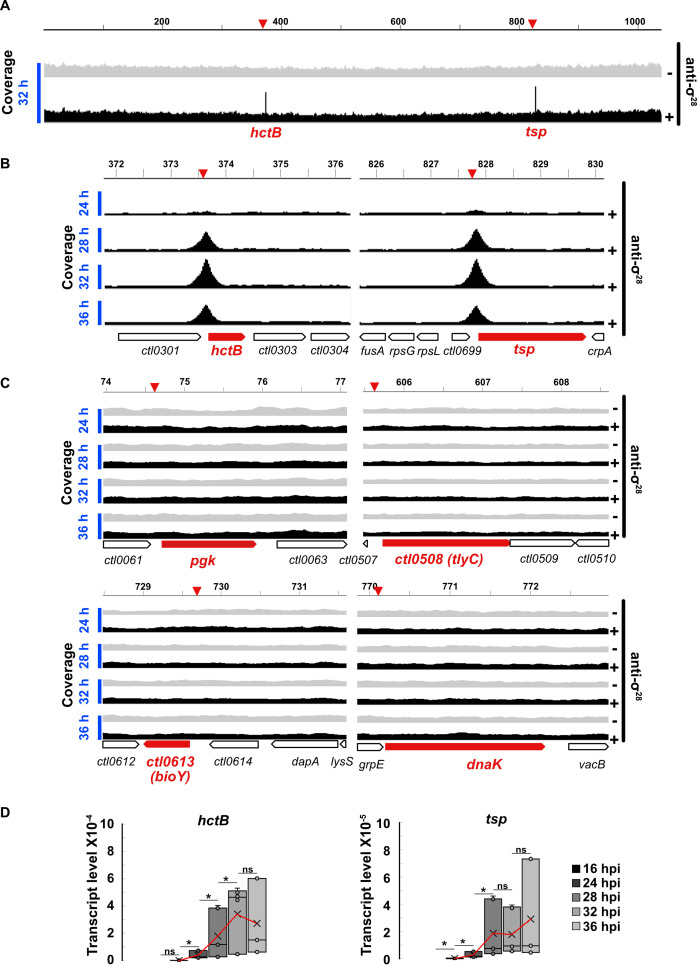 Fig 1
Fig 1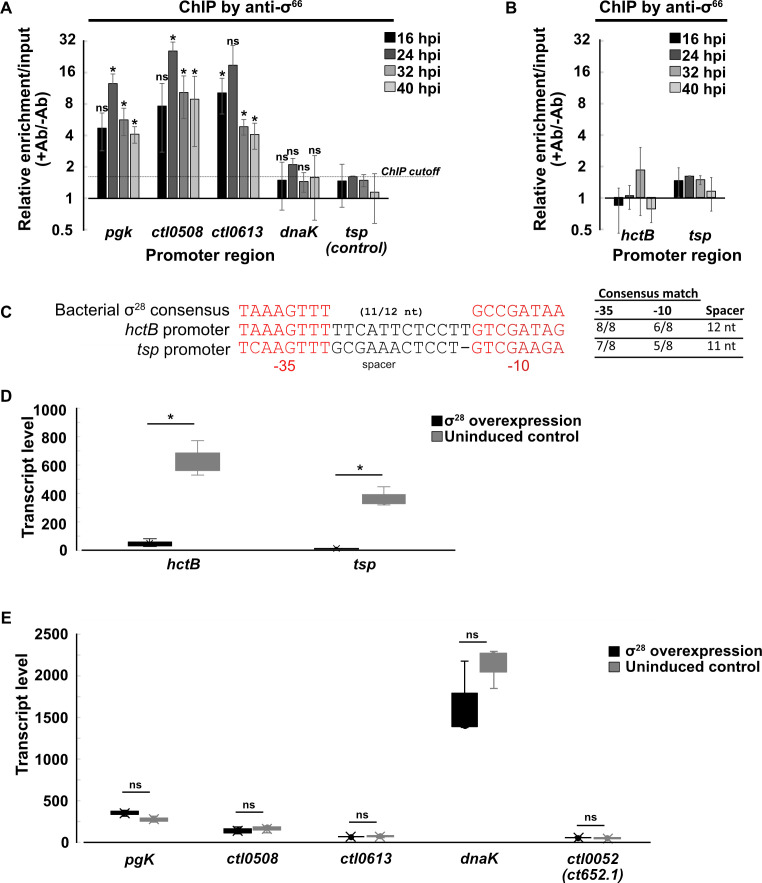 Fig 2
Fig 2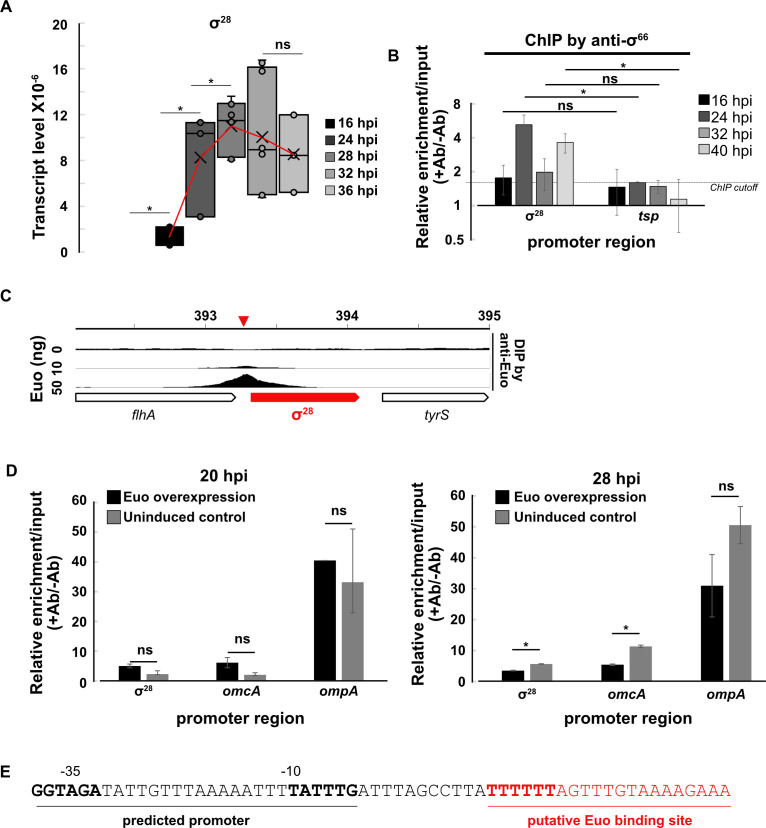 Fig 3
Fig 3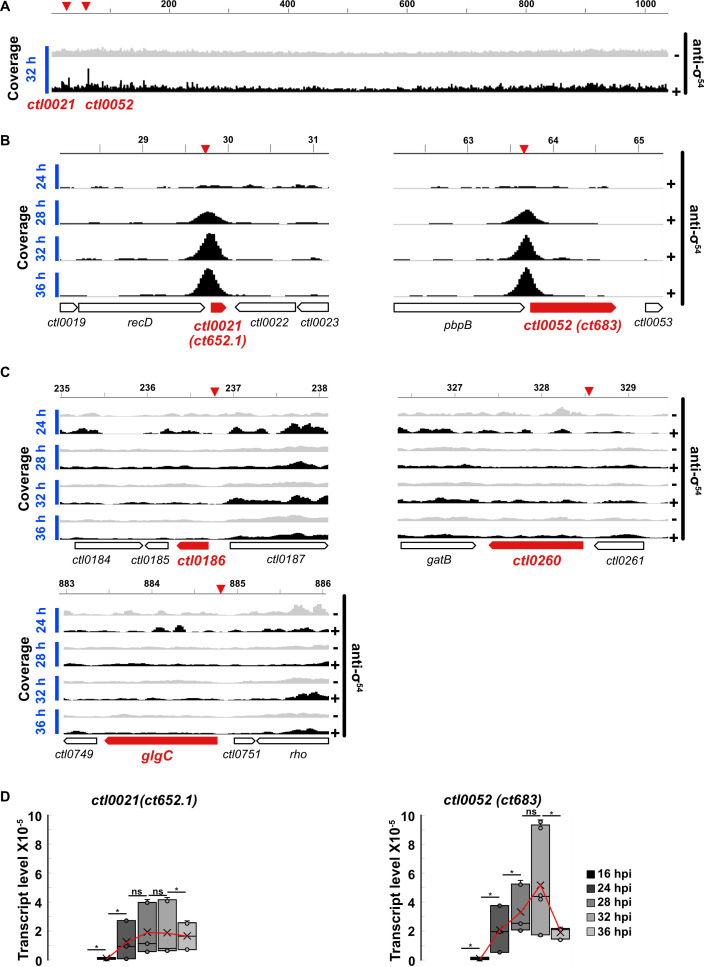 Fig 4
Fig 4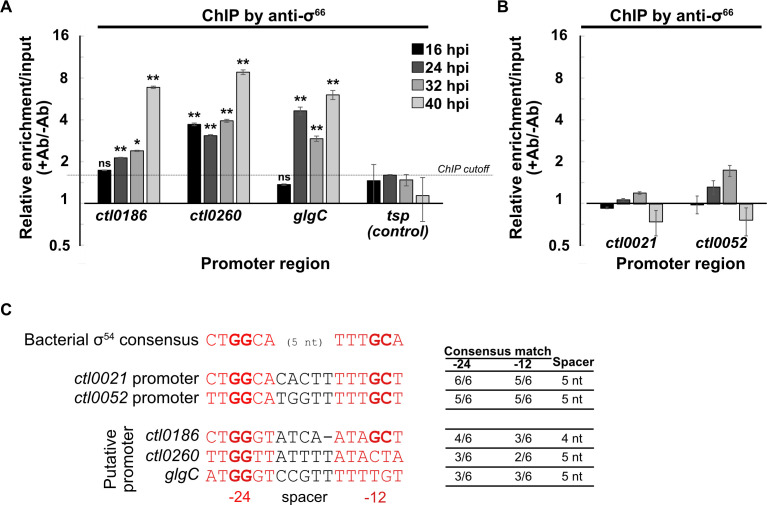 Fig 5
Fig 5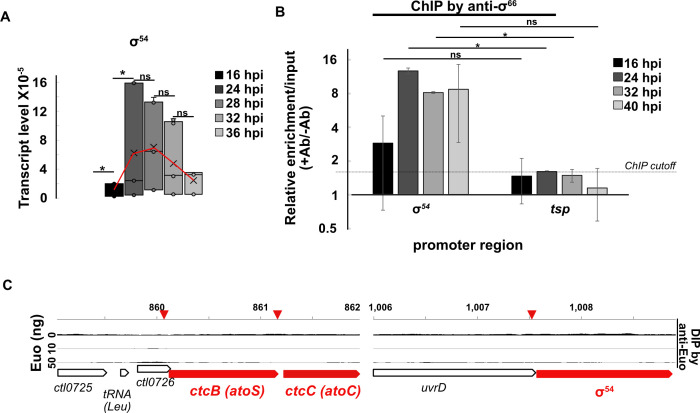 Fig 6
Fig 6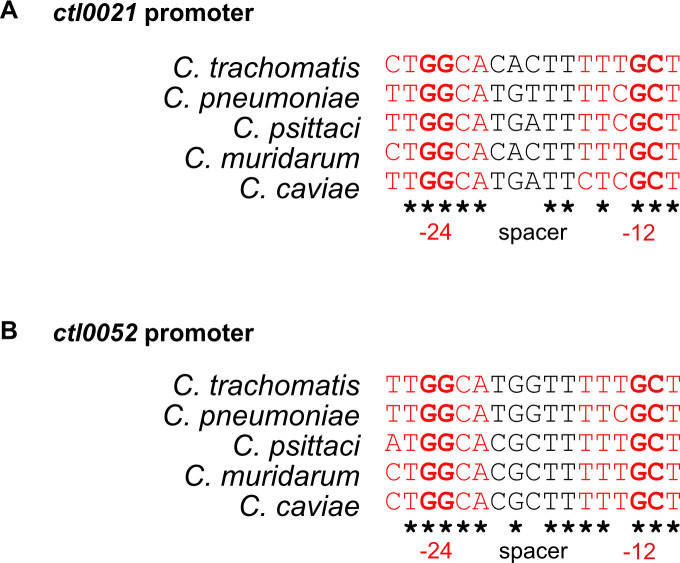 Fig 7
Fig 7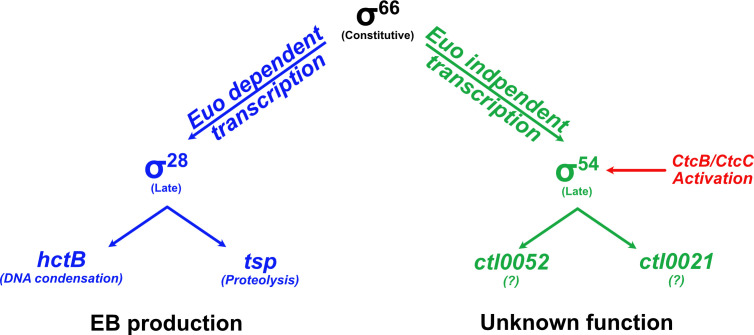 Fig 8
Fig 8- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —National Institute of Food and Agriculturehttp://dx.doi.org/10.13039/100005825
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReproductive tract infections research · Cervical Cancer and HPV Research · Adolescent Sexual and Reproductive Health
INTRODUCTION
Chlamydia is a genus of bacteria that replicates via a developmental cycle inside a eukaryotic host cell. During the 48–72 hours of intracellular infection, chlamydiae repeatedly divide and then convert into a specialized form, the elementary body (EB) (1, 2). This conversion step is critical because the replicating form, which is called a reticulate body (RB), is non-infectious, and only the EB can spread the infection to a new host cell. Chlamydia genes are expressed in three main temporal waves that are linked to this developmental cycle (3–6). Early genes are involved in establishing the intracellular infection, and midcycle genes are expressed during RB replication. Late genes are first transcribed at ~24 hours post-infection (hpi) when RB-to-EB conversion commences and includes genes that are critical for EB production (7–10). Transcriptomic studies have shown that temporal gene expression in Chlamydia trachomatis is regulated at the transcriptional level (3). The importance of transcriptional control is further demonstrated by studies showing that disruption of the transcriptional program via overexpression or depletion of transcription factors caused major defects in the developmental cycle (11, 12).
Like other bacteria, Chlamydia utilizes a multi-subunit enzyme called RNA polymerase to transcribe its genes (5). The core enzyme is the active component that synthesizes RNA, but the sigma (σ) subunit allows RNA polymerase to initiate the transcription of specific genes by recognizing and binding target promoters. The major form of chlamydial RNA polymerase contains σ^66^, which is the ortholog of σ^70^, the housekeeping sigma factor in Escherichia coli (13). Chlamydia also encodes two alternative sigma factors, σ^28^ and σ^54^, which substitute for σ^66^ to alter the promoter specificity of RNA polymerase. The promoter sequence recognized by C. trachomatis σ^28^ is similar to the E. coli σ^28^ promoter, and seven C. trachomatis promoters were transcribed by σ^28^ RNA polymerase in vitro (14). However, a σ^28^ overexpression and knockdown study reported only two σ^28^ target genes (15). σ^54^ belongs to a different family of sigma factors, and its promoter architecture differs from σ^66^ and σ^28^ (16). Sequences resembling bacterial σ^54^ promoters have been identified upstream of two genes in C. trachomatis (ctl0021/ct652.1 and ctl0052/ct683) (17, 18). However, studies using σ^54^ depletion reported 67 target genes (15), and a σ^54^ activation approach identified 33 target genes (12), with little overlap in these putative σ^54^ regulated genes.
The mechanisms that regulate the delayed transcription of late genes are of special interest because they may control the timing of RB-to-EB conversion. The best-studied regulator of late gene expression is the transcription factor Euo. Euo has been shown to be a repressor of late gene promoters and has been proposed to prevent the expression of late genes until late times (5, 11). However, not all late genes are regulated by Euo, raising the possibility of additional mechanisms to control late gene expression (11). Unlike midcycle genes, promoters of late genes do not appear to be regulated by DNA supercoiling levels (19). σ^28^ and σ^54^ have been proposed to be regulators of late gene expression (12, 14, 17) because they each have a late expression pattern (3) and have putative target genes that are late genes. However, the uncertainty about their target genes makes it difficult to draw firm conclusions about the roles of the two alternative chlamydial sigma factors in late gene regulation.
In this study, we used time-course chromatin immunoprecipitation (ChIP)-seq to identify the regulons of the two alternative sigma factors in C. trachomatis. This approach allowed us to directly measure the binding of σ^28^ or σ^54^ to their respective target genes in the genome. Our results demonstrate that each alternative sigma factor regulates fewer genes than previously reported, but all of these target genes are late genes. Thus, C. trachomatis utilizes its alternative sigma factors as additional mechanisms to control late gene expression during the developmental cycle.
MATERIALS AND METHODS
Plasmids, strains, and antibodies
All cloning was done using NEBuilder HiFi DNA Assembly kit and the parent plasmid pBOMB4 (20). σ^28^ (fliA) was cloned downstream of the tet-promoter in pBOMB4 plasmid, and the resulting pBOMB4-σ^28^ plasmid was used to transform C. trachomatis L2 434/Bu strain for overexpression. Similarly, σ^54^ (rpoN) and the ATPase domain of ctcC (Leu_131_–Stop_387_) were cloned downstream of the conditional tet-promoter and constitutive nMen-promoter, respectively, in pBOMB4, and the resulting plasmid was used to transform C. trachomatis L2 434/Bu strain to overexpress a constitutively activated σ^54^ (12). A previously reported C. trachomatis strain (11) was used to perform σ^66^ ChIP-qPCR under Euo overexpression conditions. Available rabbit polyclonal antibodies against σ^28^ (21) and σ^54^ were used for all ChIP experiments.
Cell culture and C. trachomatis transformation
Cell culture and C. trachomatis transformation were done as reported previously (11). HeLa cells were seeded and allowed to grow to ~90% confluency and infected with wild-type C. trachomatis L2 434/Bu strain at a multiplicity of infection (MOI) of ~3 by centrifugation at 750 × g for 1 hour at room temperature (RT). EBs were transformed with 5–10 µg of plasmid in 50 µL CaCl_2_ buffer.
Total RNA extraction, cDNA synthesis, genomic DNA extraction, and RT-qPCR
Total RNA was extracted from infected HeLa cells in TRIzol reagent following the manufacturer’s instructions. The aqueous phase was collected, and total RNA was precipitated by adding an equal volume of 100% ethanol. The precipitated sample was then loaded on a silica column supplied with Qiagen’s RNeasy Mini Kit. Total RNA was eluted in 50 µL nuclease-free water. RNA was aliquoted, quantified, and stored at −80°C. cDNA was constructed using 10 µL (~1 µg) of total RNA using Bio-Rad’s iScript dDNA Clear cDNA Synthesis kit. Genomic DNA (gDNA) was collected by solubilizing infected HeLa cells in 500 µL lysis solution containing 1 mM EDTA, 10 mM Tris-HCl (pH 7.5), and 0.1% SDS, and DNA was fragmented by sonicating samples for 10 cycles of 1 second on/1 second off at 30% amplitude. For qPCR, sonicated gDNA was diluted 1:100, and 3 µL of diluted gDNA was used for each 20 µL qPCR mixture. mRNA level and genome copy number for each condition were measured by qPCR.
Chromatin immunoprecipitation
Confluent (~90%–100%) HeLa cells in six-well cell culture plates infected with wild-type or transformed C. trachomatis strains at an MOI of 3, and DNA-protein interactions were fixed by incubating with 1.1% formaldehyde in 0.5 mL/well phosphate-buffered saline (PBS) for 5 minutes (σ^28^ or σ^66^) or 10 minutes (σ^54^) at RT. The reaction was quenched with 25 µL of 2.5M Glycine (RT). Fixed samples were washed (3×) with PBS at RT. For each ChIP, fixed samples from one (σ^66^) or two (σ^28^, σ^54^) six-well plates were collected in a total of 1 mL sonication buffer (1 mM EDTA, 10 mM Tris-HCl [pH 7.5], and 0.1% SDS) by scraping. Chromatin was sheared using the Covaris S220 Focused-ultrasonicator for 3 minutes for an average fragment size of ~300 bp. Dynabeads protein G magnetic beads were prepared by incubating 30 µL of a 50% slurry with polyclonal antibodies against σ^28^, σ^54^, and σ^66^ in lysis buffer for two hours at 4°C, followed by washing three times with the lysis buffer. Immunoprecipitation was done overnight by incubating sonicated lysate with the antibody-coated magnetic beads. Library was constructed using NEXTflex ChIP-seq Kit and NEXTflex barcodes (Bioo Scientific), and high-throughput sequencing was performed using Illumina Novaseq 6000 sequencing platform as reported earlier (11). Samples from all time points were sequenced to a total coverage of at least ~100-fold.
Data analysis
High-throughput sequencing data were analyzed using Qiagen’s CLC genomics workbench (v22.0.2) software. Reads were mapped to the C. trachomatis L2 434/Bu genome AM884176 using high stringency settings, and ChIP peaks were called using the “transcription factor ChIP-seq” tool. Relative enrichment values were calculated by dividing the enrichment/input for the test (+Ab) by the control (−Ab) samples. Transcript level for reverse transcriptase-quantitative polymerase chain reaction (RT-qPCR) experiments was calculated using Ct values and PCR efficiency to normalize (22) mRNA to C. trachomatis gDNA levels (11).
Prediction of σ54 promoters
Homer software (23) was used to perform promoter prediction and search. To examine which genes have σ^54^ promoters, we performed a BLAST analysis on coding sequences downstream of putative promoters in C. trachomatis L2/434/Bu genome to identify their C. trachomatis orthologs.
RESULTS
Identification of the σ28 regulon
To identify genes that are transcribed by σ^28^ RNA polymerase, we performed a σ^28^ ChIP-seq analysis on HeLa cells infected with C. trachomatis serovar L2. This approach detects σ^28^ binding sites in the C. trachomatis genome at the time of analysis in the intracellular infection. Using polyclonal antibodies against σ^28^, we immunoprecipitated DNA fragments bound by σ^28^, which were then identified by deep sequencing. σ^28^ ChIP-seq analysis at 32 hpi revealed that σ^28^ only bound two sites in the C. trachomatis genome. These ChIP-peaks were located at hctB and tsp (Fig. 1A), which are genes that have been reported to be σ^28^ targets (14, 15). We then performed a time course σ^28^ ChIP-seq from 24 to 36 hpi, which revealed that σ^28^ bound hctB and tsp at 28 hpi and later time points, with maximum binding at 32 hpi, but no σ^28^ binding was detected at 24 hpi (Fig. 1B; Fig. S1A). The σ^28^ ChIP-peaks for hctB and tsp were both centered upstream of the gene, around the −10 element of their experimentally defined σ^28^ promoters (14) (Fig. 2C).
*(A) Coverage of mapped reads showing ChIP-peaks in hctB and tsp promoter regions at 32 hpi. Light gray shows the coverage of no-antibody negative control, and black shows coverage for immunoprecipitated samples. (B) Temporal change in the binding of σ28 at the promoters of hctB and tsp. (C) The absence of σ28 ChIP-peaks in the promoter regions of previously identified σ28 transcribed genes. Genes are shown in red. Inverted wedges (red) on top of each panel show the expected location of the ChIP-peak in the promoter region. (D) Temporal expression of hctB (left) and tsp (right) measured as a ratio of mRNA to gDNA. Red line traces the mean transcript level. P < 0.05, ns: not significant, t-test, n = 3. Genomic loci are shown in kb.
*(A) ChIP-qPCR showing binding of σ66 at the promoters of pgk, ctl0508, ctl0613, and dnaK. Relative enrichment is measured as the ratio of enrichment when anti-σ66 antibodies (+Ab) are used vs mock enrichment with beads without antibody (−Ab). The specificity of σ66 ChIP is shown by the absence of enrichment of the promoter region of σ28 transcribed gene tsp. *P < 0.05, t-test, n = 2. (B) ChIP-qPCR showing binding of σ66 at the promoters of tsp and hctB. (C) Comparison of hctB and tsp promoters with the consensus bacterial σ28 promoter. (D) Overexpression of σ28 resulted in increased expression of hctB and tsp compared to wild type (un-induced control). P < 0.05, t-test, n = 3. (E) No increase was observed in the expression of pgk, ctl0508, ctl0613, or ctl0052.
We did not detect any other σ^28^ ChIP peaks in our σ^28^ ChIP-seq analysis at any time point (Fig. S1A). In particular, we did not detect σ^28^ ChIP-peaks for pgk, ctl0508 (tlyC), ctl0613 (bioY), and dnaK, which are C. trachomatis genes transcribed by σ^28^ RNA polymerase in vitro (14) (Fig. 1C). In contrast, a parallel σ^66^ ChIP-seq analysis showed that promoter regions upstream of pgk, ctl0508, and ctl0613 were each enriched for σ^66^ binding (Fig. 2A). The promoter region upstream of dnaK was not enriched for σ^66^ binding, suggesting that this gene does not have its own internal promoter within the hrcA-grpE-dnaK operon. These findings provide evidence that these putative σ^28^ targets are instead transcribed by σ^66^ RNA polymerase and not σ^28^ RNA polymerase under the growth conditions tested. In contrast, the tsp or hctB promoter regions (Fig. 2B) were not enriched for σ^66^ binding (Fig. 2B).
We performed RT-qPCR on C. trachomatis-infected cells to check if the late σ^28^ binding pattern at hctB and tsp correlated with the temporal expression patterns of these genes. We measured the greatest increase in hctB and tsp transcript levels between 24 and 28 hpi (Fig. 1D and E), concordant with the onset of σ^28^ binding between 24 hpi and 28 hpi in our ChIP-seq analysis (Fig. 1B top two panels). For additional evidence that hctB and tsp are regulated by σ^28^, we generated a C. trachomatis σ^28^ overexpression strain in which exogenous expression of σ^28^ is controlled by the addition of anhydrotetracycline to the tissue culture medium (11). Overexpression of σ^28^ during midcycle increased transcript levels of hctB and tsp (Fig. 2D) but not the four putative σ^28^ targets that σ^28^ did not bind in our ChIP assay (Fig. 2E). Collectively, these results provide direct evidence that σ^28^ binds to the promoters of hctB and tsp during the C. trachomatis developmental cycle.
The σ28 gene is transcribed by σ66 RNA polymerase and regulated by Euo
The late patterns of σ^28^ binding and σ^28^-regulated gene expression prompted us to investigate how σ^28^-dependent transcription is temporally regulated. Using RT-qPCR, we detected transcription of the σ^28^ gene (fliA), from 24 hpi onward but not at 16 hpi (Fig. 3A). In addition, σ^66^ ChIP-qPCR only detected enrichment of σ^66^ at the σ^28^ promoter region at 24 hpi and later times (Fig. 3B), indicating transcription by σ^66^ RNA polymerase at late times. Since Euo bound the σ^28^ promoter region in our previous Euo DNA immunoprecipitation (DIP) analysis (Fig. 3C) (11), we investigated if this late transcription of σ^28^ is controlled by Euo. Using a C. trachomatis Euo overexpression strain (11), we performed a σ^66^ ChIP-qPCR analysis and found that exogenous Euo expression significantly reduced σ^66^ binding at the σ^28^ promoter at 28 hpi (Fig. 3D), consistent with Euo-mediated repression. Together, these results demonstrate that the σ^28^ gene is transcribed by σ^66^ RNA polymerase, but transcription only occurs at late times because of temporal regulation by Euo. Intriguingly, Euo overexpression only reduced σ^66^ binding to σ^28^ and another Euo-regulated gene omcA, at a late time (28 hpi) and not during midcycle (20 hpi; Fig. 3D). This observation suggests that endogenous Euo levels are sufficient to repress its target genes in midcycle but are limiting at late times, which allows σ^66^ RNA polymerase to transcribe σ^28^ and other Euo-regulated late genes. A putative σ^66^ promoter and a putative Euo operator were identified upstream of the σ^28^ coding region (Fig. 3E).
*(A) Temporal expression of σ28 was measured as the number of transcripts normalized to bacterial gDNA. Red line traces the mean transcript level. (B) ChIP-qPCR showing increase in σ66 binding at the promoter of σ28 after 16 hpi. (C) DIP showing binding of Euo at σ28 promoter using 10 and 50 ng purified recombinant Euo. *P < 0.05, t-test, n = 3. (D) ChIP-qPCR shows reduced binding of σ66 in the promoter region of σ28 and omcA but not ompA due to Euo overexpression at 28 hpi but not at 20 hpi. (E) Predicted σ66 promoter sequence upstream of the coding region of the σ28 gene and putative Euo-binding site downstream of the promoter. P < 0.05, t-test, n = 2.
Identification of the σ54 regulon
Taking a similar approach, we performed a ChIP-seq time course to identify genes transcribed by σ^54^ RNA polymerase in C. trachomatis serovar L2-infected HeLa cells. ChIP-seq with anti-σ^54^ antibodies at 32 hpi revealed that σ^54^ only bound two sites in the C. trachomatis genome (Fig. 4A). These sites were located at ctl0021 and ctl0052 (ct652.1 and ct683, respectively, in C. trachomatis serovar D), which are the two best characterized σ^54^ promoters in C. trachomatis (17). σ^54^ binding to ctl0021 and ctl0052 was detected at 28 hpi and later, with maximum binding at 32 hpi and 36 hpi, but no binding was observed at 24 hpi (Fig. 4B; Fig. S1B). We did not detect σ^54^ binding to other genes at any time point from 24 to 36 hpi (Fig. S1B), including the ~100 genes that have been reported as σ^54^-regulated genes in C. trachomatis (12, 15). Our initial σ^54^ ChIP-seq assay was performed with sequencing coverage of ~50-fold, but we did not detect any additional σ^54^ ChIP peaks when we increased the depth of sequencing to achieve a coverage of more than 150-fold. The two σ^54^ ChIP-peaks were centered around ~50 bp upstream of the ctl0021 and ctl0052 coding regions, respectively (Fig. 4B). Consistent with this late σ^54^ binding pattern, ctl0021 and ctl0052 both had a late transcription pattern, as measured by RT-qPCR (Fig. 4D). The expression of both ctl0021 and ctl0052 increased significantly between 16 and 24 hpi, plateaued between 24 and 32 hpi, then decreased after 32 hpi.
*(A) Coverage of mapped reads showing ChIP-peaks in ctl0021 and ctl0052 promoter regions at 24, 28, 32, and 36 hpi. Light gray shows the coverage of no-antibody negative control, and black shows coverage for immunoprecipitated samples. (B) Temporal change in the binding of σ54 in the promoters of ctl0021 and ctl0052. (C) The absence of σ54 ChIP-peaks in the promoter region of ctl0186, ctl0260, and glgC. These genes previously identified as σ54 transcribed genes are shown in red. Inverted red wedges on top of each panel show the expected location of ChIP-peaks in the promoter region. (D) Temporal expression of ctl0021 (left) and ctl0052 (right) was measured as a ratio of mRNA to gDNA. Red line traces the mean transcript level. P < 0.05, t-test, n = 3.
Analysis of reported σ54 target genes that were not detected in our σ54 ChIP assay
To investigate the large discrepancy between the number of σ^54^ targets identified in our ChIP-seq analysis and previous RNA-seq studies, we selected three genes (ctl0186, ctl0260, and glgC) for further evaluation. These putative σ^54^ target genes were selected because they are upregulated at a late time and have the best available evidence from the Soules et al. (12) and Hatch and Ouellette reports (15), based on identification in both studies, higher levels of differential expression in response to σ^54^ depletion, and potential σ^54^ promoters identified bioinformatically (15). However, in our ChIP-seq time course, none of these genes had significant σ^54^ binding compared to a no-antibody negative control at any time point (Fig. 4C). Sequence analysis showed that the putative σ^54^ promoters of these genes each had significant differences at critical positions (GG in the −24 promoter element and GC in the −12 element) or deviation from the optimal 5 bp spacer length compared to the consensus bacterial σ^54^ promoter sequence (Fig. 5C) (24). In contrast, the predicted σ^54^ promoters of our σ^54^ targets, ctl0021 and ctl0052, showed a 100% match to these critical bases and spacer length compared to the consensus σ^54^ sequence (17). For further analysis, we performed a parallel ChIP-qPCR using anti-σ^66^ antibodies to determine if these genes are transcribed by σ^66^ RNA polymerase. We detected σ^66^ binding, as measured by σ^66^ enrichment, to ctl0186, ctl0260, and glgC (Fig. 5A), but not our σ^54^ targets, ctl0021 and ctl0052 (Fig. 5B). Collectively, these findings provide evidence that ctl0021 and ctl0052 are transcribed by σ^54^ RNA polymerase, but ctl0186, ctl0260, and glgC are transcribed by σ^66^ RNA polymerase and not σ^54^ RNA polymerase.
*(A) ChIP-qPCR showing binding of σ66 at the promoters of ctl0186, ctl0260, and glgC. Promoter region of σ28 transcribed gene tsp is used as a negative control. Relative enrichment is measured as the ratio of enrichment using anti-σ66 antibodies (+Ab) to mock enrichment with beads without antibody (−Ab). The promoter of σ28 transcribed gene tsp was not bound by σ66. *P < 0.05, *P < 0.005, t-test, n = 2. (B) ChIP-qPCR showing no binding of σ66 at the promoters of ctl0021 and ctl0052. (C) Comparison of ctl0021 and ctl0052 promoters with the consensus bacterial σ54 promoter.
The σ54 gene is transcribed by σ66 RNA polymerase but not regulated by Euo
We performed RT-qPCR to measure transcripts from the σ^54^ gene (rpoN) and detected maximal expression at late times (24 and 28 hpi; Fig. 6A). ChIP-qPCR with anti-σ^66^ antibodies showed enrichment for σ^66^ in the promoter region upstream of the σ^54^ gene (Fig. 6B). In contrast, neither σ^28^ nor σ^54^ bound this promoter region in ChIP-seq analyses with anti-σ^28^ and anti-σ^54^ antibodies (Fig. S1). These results indicate that the σ^54^ gene is transcribed at late times by σ^66^ RNA polymerase. We then investigated if σ^54^ or its predicted regulators are controlled by the late regulator Euo. The examination of our published Euo DIP peak analysis showed no Euo DIP peaks in the promoter regions of the σ^54^ gene, or ctcB (atoS) or ctcC (atoC; Fig. 6C), which encode putative two-component regulators of C. trachomatis σ^54^. Moreover, we did not detect differential expression of σ^54^, ctcB, or ctcC in a previous report where we performed differential RNA-seq in response to Euo overexpression (11). Thus, Euo does not appear to be responsible for the late expression of σ^54^ or the CtcB/CtcC activation system. Overall, these results show that the σ^54^ gene is transcribed by σ^66^ RNA polymerase, but the late expression or activation of σ^54^ is regulated by an additional mechanism other than Euo.
*(A) Temporal expression of σ54 measured as the number of transcripts normalized to bacterial gDNA. Red line traces the mean transcript level. *P < 0.05, t-test, n = 3. (B) ChIP-qPCR showing increase in σ66 binding at the promoter of the σ54 gene after 16 hpi. P < 0.05, t-test, n = 2. (C) DIP shows no binding of Euo in the putative promoter region of ctcB, ctcC, or σ54. Inverted red wedges on top of the panels show the expected location of Euo DIP-peaks if Euo binds these genes.
In silico prediction of σ54 regulated genes in Chlamydia and Chlamydia-related bacteria
To investigate if σ^54^ has a conserved role in regulating chlamydial gene expression, we performed an in silico analysis to identify σ^54^-regulated genes in other Chlamydia spp. and Chlamydia-related bacteria. We based this analysis on ctl0021 rather than ctl0052 because Ctl0021 is conserved in all sequenced Chlamydia and Chlamydia-related bacteria, with only one exception (Table S1). We aligned the non-coding regions immediately upstream of each ctl0021 gene and identified a putative σ^54^ promoter in every case (Table S1). We then used these promoter sequences to define a consensus sequence (TGGCACAATTTTCGCT), position weight matrix, and a selection cutoff, enabling us to predict σ^54^ promoters using the promoter prediction tool Homer (23). When we applied this prediction analysis to the C. trachomatis L2/434/Bu genome, we successfully predicted the σ^54^ promoter of ctl0052, along with a few hits that are likely to be false positives because they were within coding regions. We then extended this promoter prediction to other Chlamydia spp. and Chlamydia-related bacteria, limiting our analysis to promoter-like sequences in non-coding regions. Almost all the predicted promoters were upstream of ctl0021 and ctl0052 orthologs (Fig. 7; Table S1). A notable exception was Chlamydia pneumoniae, which had σ^54^ promoter-like sequence upstream of ftsK in addition to ctl0021 and ctl0052. Two Chlamydia-related bacteria, ca. Protochlamydia amaebophila and ca. Protochalmydia naegleriophila, appear to lack ctl0052 but had σ^54^ promoter-like sequence upstream of ctl0021 and additional genes not found in C. trachomatis. This in silico analysis provides evidence of a transcriptional unit composed of the alternative sigma factor σ^54^ and its target genes, ctl0021 and ctl0052, conserved in most Chlamydia spp. and Chlamydia-related bacteria.
σ54 promoter-like sequences identified upstream of ctl0021 (A) and ctl0052 (B) homologs in C. pneumoniae (CWL029), Chlamydia psittaci (Cps_WRSTE30), Chlamydia muridarum (Cmu_Nigg3), and Chlamydia caviae (Cca_GPIC) are conserved with all the critical bases (bold) matching in the −12 and −24 (red) elements of the σ54 promoters C. trachomatis (Ctr_434/Bu) and ideal spacer lengths (5 bp) maintained between the −12 and −24 elements.
DISCUSSION
In this study, we used time-course ChIP-seq and additional data to identify genes bound by alternative sigma factors during the C. trachomatis developmental cycle. We found that σ^28^ and σ^54^ each bound two genes and only late in the intracellular infection. The small number of σ^28^ target genes is consistent with a published report (15), but we found evidence for far fewer σ^54^ target genes than previous studies, as will be discussed further (12, 15). The temporal pattern of σ^28^ and σ^54^ binding can be explained by the late expression of the genes encoding these alternative sigma factors. However, only the σ^28^ gene was regulated by the late regulator Euo, and an unknown Euo-independent mechanism controls the temporal expression of the σ^54^ gene. These findings indicate that there are multiple mechanisms to regulate late gene expression in Chlamydia.
ChIP-seq provides several advantages for identifying target genes regulated by a specific form of chlamydial RNA polymerase (25–27). Because it is a direct binding assay, ChIP provides stronger evidence than differential expression of a gene, which could be due to direct regulation by the sigma factor or an indirect effect via another factor. The cross-linking step in ChIP preserves the physical interaction between the sigma factor and each target gene, which allows binding to be measured within the bacterium, even inside a host cell. Binding can thus be measured during the normal developmental cycle, unlike approaches that induce differential expression but may introduce other changes to the intracellular infection. By performing the cross-linking step at different times in the developmental cycle, we were able to show that σ^28^ and σ^54^ only bound at late times. In addition, this ChIP analysis could be performed under different growth conditions, such as chlamydial persistence or hypoxia, to measure their effects on transcription. A further advantage is in promoter identification because the σ subunit binds the promoter of the gene—where RNA polymerase initiates transcription—but not the coding region since σ is released with promoter clearance (28).
We performed additional studies to bolster our ChIP-seq target identification. For σ^28^, we used a C. trachomatis σ^28^ overexpression strain to show that transcription of hctB and tsp was σ^28^-dependent (Fig. 2D). Published in vitro and differential RNA-seq studies have also identified hctB and tsp as σ^28^-transcribed genes (14, 15, 21). However, four other reported genes with σ^28^-transcribed promoters do not appear to be bona fide targets because they lacked σ^28^ binding (Fig. 1C) and σ^28^-dependent transcription in chlamydiae (Fig. 2E) and were not detected in a σ^28^ overexpression/knockdown transcriptomic study (15). This discrepancy demonstrates the limitations of identifying target genes based on transcription by σ^28^ RNA polymerase in vitro or in a heterologous system (14, 29). An additional drawback of in vitro or reporter assays is that they are usually analyzed with a single promoter in isolation, without competition from other promoters in the genome. These four genes are likely transcribed by σ^66^ RNA polymerase as they were bound by σ^66^ (Fig. 2A).
There are good data to support the two σ^54^ target genes that we identified. ctl0021 and ctl0052 each have promoter sequences that are well conserved with the consensus bacterial σ^54^ promoter sequence (Fig. 5C). These promoter sequences are in the correct location upstream of in vivo transcription start sites mapped in C. trachomatis by Timms and colleagues (17), whereas transcription start sites have not been verified for other putative σ^54^ promoters. We were unable to generate a C. trachomatis σ^54^ overexpression strain, which requires expression of σ^54^ and activation by the CtcB/CtcC two-component system (30). However, we showed a temporal correlation between σ^54^ binding (Fig. 4B) and the late transcription of ctl0021 and ctl0052 in C. trachomatis-infected cells (Fig. 4D). In addition, we used a stringent in silico analysis to identify well-conserved σ^54^ promoter sequences upstream of ctl0021 and ctl0052 homologs in all Chlamydia spp. and almost all Chlamydia-related bacteria (Fig. 7; Table S1). Collectively, these data provide evidence that ctl0021 and ctl0052 are regulated by σ^54^ as a transcriptional unit conserved in the order Chlamydiales.
Approximately 100 σ^54^ target genes have been proposed for C. trachomatis but were not bound by σ^54^ in our ChIP-seq study. These genes were identified on the basis of differential transcription in response to exogenous expression of the putative σ^54^ activator, CtcC (12), or to knockdown of σ^54^ (15). However, transcriptional changes were detected by RNA-seq, which does not indicate if the transcripts were produced by σ^54^ RNA polymerase or other chlamydial RNA polymerases. Importantly, differential expression may not have been specific for σ^54^ because σ^54^ activation and knockdown both disrupted the progression of the developmental cycle (12, 15). The likelihood of non-specific differential expression is further suggested by the inability of the σ^54^ knockdown RNA-seq study to identify either ctl0021 or ctl0052 as targets (15) and for the σ^54^ activation study to only identify ctl0052 and not ctl0021 (12). In addition, we postulate that σ^54^ knockdown may not have altered σ^54^-dependent transcription because it was measured at 24 hpi—a time in the developmental cycle before σ^54^ binding is detected (Fig. 4B; Fig. S1B).
We performed additional analyses to investigate this discordance in the identification of σ^54^ target genes. We increased the depth of sequencing for our ChIP-seq analysis by 150-fold but still did not detect σ^54^ binding to genes other than ctl0021 or ctl0052. Furthermore, our ChIP-PCR analysis of ctl0186, ctl0260, and glgC, which are putative σ^54^ target genes with the best-supporting evidence, showed that they are transcribed by σ^66^ RNA polymerase rather than σ^54^ RNA polymerase. We did not analyze all the other proposed σ^54^ target genes, but our ChIP-seq study casts considerable doubt on them, and additional experimental data are required to show that they are bona fide σ^54^ targets.
σ^54^ RNA polymerase could possibly transcribe additional genes besides ctl0021 and ctl0052 under conditions not included in our analysis, but differential expression of these genes would require a target-specific σ^54^ regulator, which has not yet been identified in Chlamydia. In contrast, CtcC, the putative activator of Chlamydia σ^54^, is likely to activate σ^54^ RNA polymerase in a general manner because it lacks a DNA-binding domain for selective binding to specific target genes (31).
One of the major insights of this study is that there are multiple mechanisms of late gene regulation in C. trachomatis, including dedicated mechanisms to regulate specific subsets of genes (Fig. 8). As σ^28^ and σ^54^ appear to each transcribe a limited subset of late genes, most late genes are likely transcribed by the major chlamydial RNA polymerase, σ^66^ RNA polymerase. Since this polymerase transcribes early, midcycle, and late genes, additional mechanisms are required to prevent premature expression of σ^66^-dependent late genes. Euo is a Chlamydia-specific transcription factor that has been shown to repress promoters of σ^66^-dependent late genes and is thus an important late regulator (11, 32). However, not all late genes are bound by Euo, so it is likely that there are additional mechanisms to control late gene expression by σ^66^ RNA polymerase (11).
Alternative sigma factors σ28 and σ54 are under the transcriptional control of the constitutively expressed housekeeping sigma factor σ66, but they are expressed late. The transcriptional repressor of the late gene, Euo, represses the expression of σ28 before late times, acting as a temporal switch. How the late transcription of σ54 is regulated remains unknown. However, its activation may be controlled by an extracellular signal via the CtcB/CtcC two-component system.
While σ^28^ and σ^54^ provide mechanisms of late gene expression, they are downstream of σ^66^ in the hierarchy of late gene regulation (Fig. 8). The genes for both σ^28^ and σ^54^ are transcribed by σ^66^ RNA polymerase and therefore must be regulated by additional mechanisms to control their late expression/activity. σ^28^ expression is regulated by Euo (Fig. 3C through E) and has also been reported to be controlled by the transcriptional activator GrgA (33, 34). In contrast, we showed that the genes for σ^54^ and its activator (CtcB/CtcC) are not regulated by Euo (Fig. 6C), and thus, the mechanism that controls the late expression of σ^54^ is unknown. In addition, σ^54^ activation by CtcB/CtcC could be temporally controlled, perhaps in response to an environmental cue.
The small size of the σ^28^ and σ^54^ regulons suggests that these target genes have important roles that must be temporally regulated. The two targets of σ^28^ are Tsp and HctB, which have each been implicated in EB production. Tsp is an ortholog of tail-specific proteases in other bacteria, which are serine proteases that target and process periplasmic proteins (35). C. trachomatis Tsp has been shown to have both proteolytic and chaperone activity (36, 37) and was necessary for RB-to-EB conversion in C. trachomatis (38). However, it is not known if Tsp is essential because a Chlamydia muridarum temperature-sensitive tsp null mutant was still able to complete the developmental cycle in a cell culture infection, although with delayed EB-to-RB conversion (39). HctB is uncommon among bacterial proteins in having sequence similarity to eukaryotic histone H1 (40). This histone-like protein is only found in EBs and not RBs and binds and condenses DNA, producing a nucleoid structure and altering transcription (9, 41). The functions of the two σ^54^-regulated proteins, Ctl0021 and Ctl0052, have not been studied, but they are conserved in all Chlamydia (Table S1). Ctl0052 contains five tetratricopeptide repeats, which are structural motifs that mediate protein-protein interactions. This suggests that its function may involve interaction with one or more other chlamydial proteins. Ctl0021 is an uncharacterized protein conserved in all Chlamydia and Chlamydia-related bacteria with no homologs in other bacteria.
The existence of multiple mechanisms to regulate late genes in C. trachomatis indicates that late gene expression is a critical step in the developmental cycle. From this and previous studies, we now know that subsets of late genes are controlled by different mechanisms, including transcription factors and alternative sigma factors. This well-orchestrated control provides the means to regulate specific target genes in response to specific internal and/or environmental signals. These separate, and in some cases overlapping, control mechanisms can be viewed as locks, each with its own key, that must all be unlocked to allow the terminal differentiation of an RB into an EB. In the case of σ^28^ and σ^54^, C. trachomatis utilizes two dedicated forms of RNA polymerase to each transcribe just a few late genes. This exquisite level of control shows the importance of late gene regulation in controlling the production of EBs to spread intracellular infection.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Moulder JW. 1991. Interaction of chlamydiae and host cells in vitro. Microbiol Rev 55:143–190. doi:10.1128/mr.55.1.143-190.19912030670 PMC 372804 · doi ↗ · pubmed ↗
- 2Abdelrahman YM, Belland RJ. 2005. The chlamydial developmental cycle. FEMS Microbiol Rev 29:949–959. doi:10.1016/j.femsre.2005.03.00216043254 · doi ↗ · pubmed ↗
- 3Belland RJ, Zhong G, Crane DD, Hogan D, Sturdevant D, Sharma J, Beatty WL, Caldwell HD. 2003. Genomic transcriptional profiling of the developmental cycle of Chlamydia trachomatis. Proc Natl Acad Sci USA 100:8478–8483. doi:10.1073/pnas.133113510012815105 PMC 166254 · doi ↗ · pubmed ↗
- 4Nicholson TL, Olinger L, Chong K, Schoolnik G, Stephens RS. 2003. Global stage-specific gene regulation during the developmental cycle of Chlamydia trachomatis. J Bacteriol 185:3179–3189. doi:10.1128/JB.185.10.3179-3189.200312730178 PMC 154084 · doi ↗ · pubmed ↗
- 5Rosario CJ, Soules K, Hefty PS, Tan M. 2020. Chlamydia gene regulation, p 219–240. Caister Academic Press.
- 6Humphrys MS, Creasy T, Sun Y, Shetty AC, Chibucos MC, Drabek EF, Fraser CM, Farooq U, Sengamalay N, Ott S, Shou H, Bavoil PM, Mahurkar A, Myers GSA. 2013. Simultaneous transcriptional profiling of bacteria and their host cells. P Lo S One 8:e 80597. doi:10.1371/journal.pone.008059724324615 PMC 3851178 · doi ↗ · pubmed ↗
- 7Moelleken K, Hegemann JH. 2008. The Chlamydia outer membrane protein Omc B is required for adhesion and exhibits biovar-specific differences in glycosaminoglycan binding. Mol Microbiol 67:403–419. doi:10.1111/j.1365-2958.2007.06050.x 18086188 PMC 2229832 · doi ↗ · pubmed ↗
- 8Barry CE, Hayes SF, Hackstadt T. 1992. Nucleoid condensation in Escherichia coli that express a chlamydial histone homolog. Science 256:377–379. doi:10.1126/science.256.5055.3771566085 · doi ↗ · pubmed ↗