From cantaloupe to cattle: Pseudomonas alabamensis sp. nov. described from diseased cantaloupe (Cucumis melo) foliage and a bovine (Bos taurus) nasopharynx
Kiersten R. Fullem, Michelle P. MacLellan, Fanny B. Iriarte, Mousami Poudel, Sarah Capik, Keith Dedonder, James L. Bono, Dayna M. Harhay, Gregory P. Harhay, Erica M. Goss, Neha Potnis, Gerald V. Minsavage, Jeffrey B. Jones, Mathews L. Paret

TL;DR
Scientists discovered a new species of bacteria, Pseudomonas alabamensis, from a diseased cantaloupe plant and a cow in the U.S.
Contribution
The paper introduces Pseudomonas alabamensis as a novel bacterial species based on genomic and phenotypic analyses.
Findings
Genomic and phylogenetic analyses confirmed the isolates belong to a new Pseudomonas species.
ANIb and isDDH scores confirmed the isolates as members of the same species.
Biochemical and mass spectrometry profiling failed to match the isolates to existing species.
Abstract
In 2022, a fluorescent bacterial isolate, designated 22-AL-CL-001, was isolated from diseased cantaloupe (Cucumis melo) foliage collected in Alabama, USA, displaying marginal necrosis and general blighting. Whole-genome sequencing and subsequent comparisons to publicly available bacterial genomes identified Pseudomonas isolate USDA-ARS-USMARC-56711, isolated from a clinically healthy calf (Bos taurus) in Kansas, USA, as genetically similar to isolate 22-AL-CL-001. The two isolates were then characterized using phenotypic and genetic analyses, including fluorescence on King’s medium B; LOPAT reactions; pathogenicity assays on cantaloupe, watermelon and squash plants; 16S rRNA gene sequence analysis; phylogeny based on multi-locus sequence analysis (MLSA) of four housekeeping genes; average nucleotide identity based on blast (ANIb); in silico DNA–DNA hybridization (isDDH) including…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
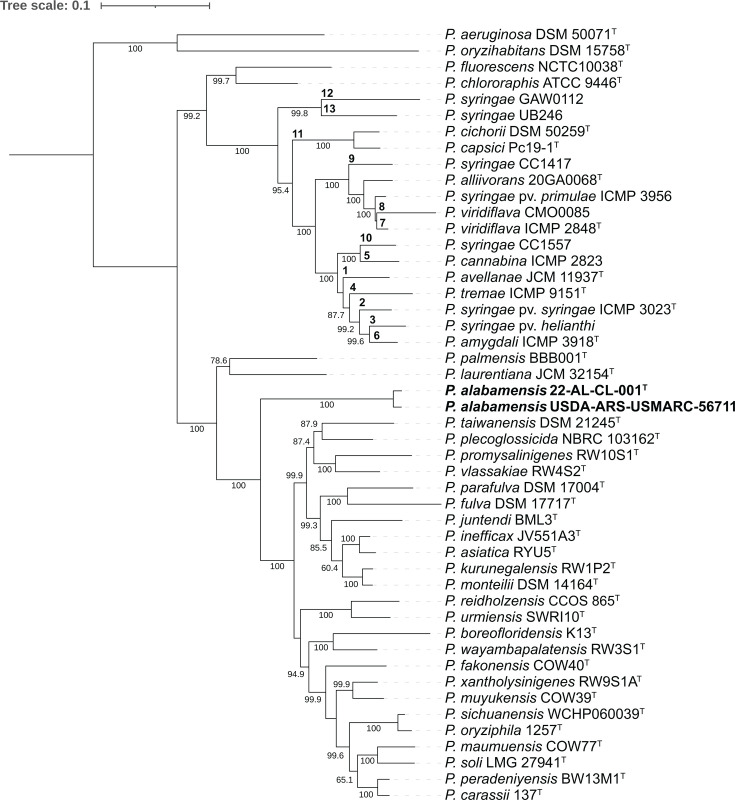 Fig. 1
Fig. 1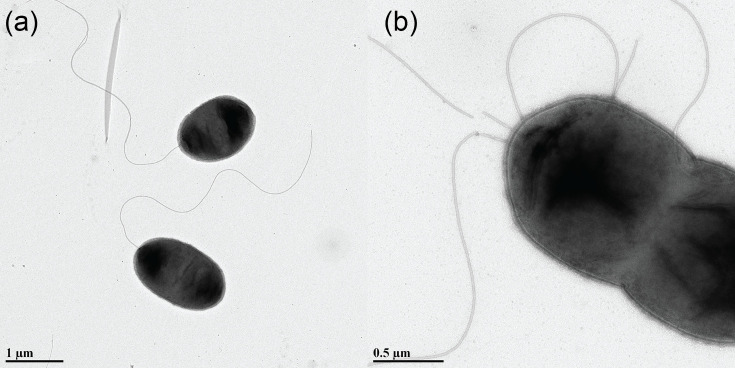 Fig. 2
Fig. 2|
|
| ||
|---|---|---|---|
| Phenotype | 22-AL-CL-001T | USDA-ARS-USMARC-56711 | DSM 14164T |
| Fluorescence | + | + | + |
| Levan | − | − | − |
| Oxidase | − | − | + |
| Pectolytic activity | − | − |
|
| Arginine dihydrolase production | − | − | + |
| Hypersensitive response on tobacco | − | − |
|
| Hypersensitive response on tomato | − | − |
|
| Isolate | 22-AL-CL-001T | USDA-ARS-USMARC-56711 |
|---|---|---|
| Genome coverage | 91 | 113 |
| Genome length (Mb) | 4.66 | 4.7 |
| N50 value | 267674 | 4714359 |
| G+C content (mol%) | 64.5 | 64.5 |
| Number of contigs | 67 | 1 |
| Completeness (%) | 99.58 | 99.57 |
| Contamination | 0.48 | 0.62 |
| 16S rRNA gene sequence similarity | ANIb | isDDH | |||||
|---|---|---|---|---|---|---|---|
| Strains | NCBI accession | 1* | 2 | 1* | 2 | 1* | 2 |
|
| GCF_030580815.1 | 100 | 99.93 | 100 | 97.25 | 100 | 78.5 |
|
| GCA_001534745.1 | 99.93 | 100 | 97.25 | 100 | 78.5 | 100 |
|
| |||||||
| GCF_000621245.1 | 98.2 | 98.2 | 81.2 | 81.21 | 24.8 | 24.8 | |
| GCF_014268935.2 | 98.68 | 98.71 | 80.68 | 80.74 | 25.9 | 26.3 | |
| GCF_019139535.1 | 98.96 | 99.02 | 80.29 | 80.33 | 26 | 26 | |
| GCF_014268885.2 | 98.82 | 98.89 | 80.23 | 80.3 | 25.9 | 26 | |
| GCF_003231305.1 | 98.68 | 98.77 | 80.09 | 80.07 | 26.2 | 26.3 | |
| GCF_036324045.1 | 98.68 | 98.71 | 80.07 | 80.07 | 25.9 | 26.1 | |
| GCF_030160515.1 | 98.89 | 98.93 | 80.06 | 80.1 | 25 | 25.9 | |
| GCF_003940825.1 | 98.84 | 98.84 | 79.98 | 80 | 26.2 | 26.4 | |
| GCF_009932335.1 | 99.03 | 99.09 | 79.98 | 79.9 | 26 | 26.2 | |
| GCF_019139895.1 | 98.82 | 98.89 | 79.86 | 79.88 | 25.9 | 25.9 | |
| GCF_019139675.1 | 98.96 | 99.02 | 79.82 | 79.8 | 25.4 | 25.7 | |
| GCA_014268975.2 | 98.91 | 98.97 | 79.62 | 79.67 | 25.2 | 25.4 | |
| GCF_900110655.1 | 98.84 | 98.89 | 79.54 | 79.56 | 25.1 | 25.3 | |
| GCF_009932375.1 | 99.1 | 99.15 | 79.4 | 79.45 | 24.9 | 25.2 | |
| GCF_030580795.1 | 99.65 | 99.55 | 79.39 | 79.33 | 24.7 | 24.8 | |
| GCF_900536025.1 | 98.54 | 98.54 | 79.38 | 79.33 | 25.5 | 25.6 | |
| GCF_000425785.1 | 98.89 | 98.91 | 79.36 | 79.29 | 25.8 | 25.1 | |
| GCF_014269035.2 | 99.02 | 99.22 | 79.33 | 79.32 | 25.3 | 25.3 | |
| GCF_900277125.1 | 98.96 | 99.02 | 79.32 | 79.4 | 26 | 26.3 | |
| GCF_014269245.2 | 99.05 | 99.16 | 79.28 | 79.17 | 24.9 | 25.1 | |
| GCF_014268815.2 | 99.03 | 99.09 | 79.08 | 79.12 | 25.2 | 25.3 | |
| GCF_000425765.1 | 99.72 | 99.73 | 78.85 | 78.83 | 24.2 | 24.4 | |
| GCF_014269025.2 | 99.02 | 99.16 | 78.66 | 78.75 | 24.3 | 24.5 | |
| GCF_017848315.1 | 97.78 | 97.92 | 78.47 | 78.39 | 24.1 | 24.2 | |
| GCF_000621265.1 | 98.96 | 98.95 | 78.45 | 78.49 | 23.9 | 24.1 | |
| GCF_014648275.1 | 99.09 | 99.12 | 77.12 | 77.13 | 22.7 | 22.8 | |
| GCF_028747405.1 | 97.1 | 97.1 | 76.56 | 76.6 | 22.7 | 22.8 | |
|
| |||||||
| GCF_900475215.1 | 96.61 | 96.84 | 75.19 | 75.28 | 21.8 | 22 | |
| GCF_012987025.1 | 95.57 | 95.46 | 75.05 | 75.03 | 21.7 | 21.5 | |
| GCF_000507185.2 | 96.82 | 96.97 | 74.57 | 74.64 | 21.3 | 21.7 | |
| GCF_012986195.1 | 98.47 | 98.56 | 74.02 | 73.66 | 21 | 21 | |
- —http://dx.doi.org/10.13039/100005825 National Institute of Food and Agriculture
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogenic Bacteria Studies · Plant Pathogens and Fungal Diseases · Plant-Microbe Interactions and Immunity
Introduction
Pseudomonas is a large and ecologically significant bacterial genus, currently containing over 330 validly published species [1]. Members of the genus are genetically and phenotypically diverse and metabolically versatile, allowing them to inhabit an array of niches, including nearly all natural environments, as well as form intimate relationships with other living organisms, which may range from beneficial to parasitic [25]. Pseudomonads, in general, are greatly adaptable, and many are capable of switching between lifestyles, niches and hosts opportunistically. This is true of numerous plant-pathogenic Pseudomonas species, which can be found in a multitude of different environments, often spending part of their lifecycle as epiphytes or endophytes of plants, within seeds or in soils [25].
Remarkably, multiple species of plant-pathogenic or plant-associated Pseudomonas are also capable of inhabiting animal hosts. Pseudomonas oryzihabitans and Pseudomonas aeruginosa, which are primarily considered environmental saprophytes, are known to opportunistically infect humans as well as plants [69]. The two species cause nosocomial infections in humans, generally those who are immunocompromised or experiencing other illnesses, and P. aeruginosa is also associated with multiple diseases of other animals [3,6, 913]. Both are occasionally documented as plant pathogens, with P. aeruginosa causing general rot symptoms in such hosts as onion, sugar cane, lettuce and tobacco and P. oryzihabitans causing panicle blight of rice as well as stem and leaf rot and internal blackening of cantaloupe [8,9, 1416]. Pseudomonas fluorescens and Pseudomonas chlororaphis, both members of the P. fluorescens species complex, are also known to be opportunistic human pathogens as well as saprophytic inhabitants of plant roots [1718]. Additionally, epiphytic strains of the species Pseudomonas syringae, a prolific plant pathogen which infects many major crop species [19], have been documented as pathogenic, and sometimes lethal, to certain species of whiteflies and aphids [20].
While only a relatively small number of all described Pseudomonas species have been reported to infect or colonize both plants and animals, the versatility and opportunistic nature of the genus as a whole suggests that similar host associations within other species may exist undiscovered. As whole-genome sequencing of bacterial isolates becomes more prevalent and resulting sequence data are deposited in publicly available online databases, more associations between plant and animal-associated Pseudomonas isolates will likely be uncovered. In this paper, we present the novel species Pseudomonas alabamensis as an example of such an occurrence.
In June of 2022, a sample of diseased cantaloupe (Cucumis melo) foliage was collected from a commercial field experiencing marginal leaf necrosis and blighting in southern Alabama and brought to the University of Florida’s Plant Diagnostic Clinic in Quincy, Florida. Bacterial isolate 22-AL-CL-001, a fluorescent pseudomonad, was isolated from a necrotic lesion from the sample as part of a study identifying and characterizing pseudomonads associated with bacterial leaf spot of cucurbits (BLS) in the Southeastern United States [21]. While BLS is most commonly attributed to P. syringae, as well as a few additional plant-pathogenic Pseudomonas species [21], isolate 22-AL-CL-001 could not be identified as a member of any existing Pseudomonas species through phenotypic and genetic analyses. Comparison of 16S rRNA gene sequence data to the National Center for Biotechnology Information’s (NCBI) nucleotide database revealed a high level of genetic similarity within the region to that of Pseudomonas isolate USDA-ARS-USMARC-56711, which was, at that time, classified within the database as a member of the species Pseudomonas monteilii. Interestingly, the two isolates had come from very different isolation sources, with isolate USDA-ARS-USMARC-56711 having been collected in 2013 from a nasopharyngeal swab of a clinically healthy calf (Bos taurus) as part of an epidemiological study of bovine respiratory disease complex [22]. The calf in question had been sourced from a Tennessee sale barn, transported to Kansas overnight and sampled the next day, whereupon USDA-ARS-USMARC-56711 was recovered from the animal. As a result, the exact geographic origin of the isolate is uncertain, as it may have been acquired by the calf during transport from other calves, the trailer or another environmental source prior to transport. Comparisons of the two isolates using average nucleotide identity based on blast (ANIb) confirmed a high level of genetic similarity, as well as a relatively low percent ANI to the type strain of P. monteilii. And so, isolate USDA-ARS-USMARC-56711 was acquired by the University of Florida for characterization and further comparison to unidentified isolate 22-AL-CL-001.
In this paper, we describe the phenotypic and genetic methods used to characterize and identify the two isolates, including fluorescence, LOPAT testing [levan production, oxidase activity, pectolytic activity on potato, arginine dihydrolase activity and hypersensitive response (HR) on tomato and tobacco] [23], whole-genome sequencing, 16S rRNA gene sequence analysis, phylogeny based on multi-locus sequence analysis (MLSA) of four housekeeping genes, average nucleotide identity based on blast (ANIb), in silico DNA–DNA hybridization (isDDH), including comparison to the Type Genome Server (TYGS) and biochemical profiling with the Biolog Gen III microplate system. Additionally, we present further characterization of isolate 22-AL-CL-001 with transmission electron microscopy and matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) as well as comparisons of 22-AL-CL-001 and USDA-ARS-USMARC-56711 to genetically similar strains identified by the Genome Taxonomy Database (GTDB).
Methods and results
Phenotypic assays
Basic phenotypic assay results were consistently identical between isolates 22-AL-CL-001 and USDA-ARS-USMARC-56711. Both isolates were determined to be Gram-negative using a standard potassium hydroxide test [24] and produced round, bright yellow, glossy colonies when grown on nutrient agar (NA, Difco) for 48 h at 28 °C. When grown on King’s medium B for 24–48 h, the two isolates produced a diffusible pigment which fluoresced under UV light. LOPAT results for both isolates indicated negative reactions for levan production, oxidase activity, pectolytic activity on potato, arginine dihydrolase production and HR in both tomato and tobacco plants (Table 1).
Isolates were also assessed for pathogenicity to cantaloupe (Cucumis melo), watermelon (Citrullus lanatus) and squash (Cucurbita pepo subsp. pepo) plants. Four-week-old seedlings were spray-inoculated until runoff with bacterial suspensions at a concentration of 10^8^ c.f.u. ml^−1^ and then placed into moistened polyethylene bags. Negative controls included completely untreated plants, plants mock-inoculated with sterile water and plants mock-inoculated with sterile water and placed into moistened polyethylene bags. P. syringae strain 13-139B, known to cause bacterial leaf spot on all three cucurbit hosts, was used as a positive control [25]. Each treatment consisted of a single pot, containing three seedlings, and was replicated in triplicate. Plants were allowed to incubate for 72 h under greenhouse conditions (day temperature, 28 °C; night temperature, 24 °C), after which, bags were removed, and plants were assessed for disease symptoms. Plants inoculated with the novel isolates exhibited no disease symptoms after 72 h. Positive control treatments exhibited typical BLS symptoms, while all negative control treatments were asymptomatic.
Whole-genome sequencing and 16S rRNA gene sequence analysis
DNA extraction for isolate 22-AL-CL-001 was performed using the Wizard Genomic DNA Purification kit (Promega). DNA was sent to the Microbial Genome Sequencing Center (Pittsburgh, PA, USA) for whole-genome sequencing with the Illumina NextSeq 2000 platform. Resultant reads were assembled using a pipeline containing the genome assembly tool SPAdes (v. 3.10.1) [26]. Isolate USDA-ARS-USMARC-56711 had been previously sequenced using a PacBio RS II sequencer and P5 chemistry and then assembled using the PacBio Hierarchical Genome Assembly Process (HGAP3, v. 2.3.0). Genomes for both isolates were smaller than is typical of Pseudomonas species, with isolate 22-AL-CL-001 having a genome length of 4.66 Mb and isolate USDA-ARS-USMARC-56711 having a genome length of 4.70 Mb. However, checkM assays showed that both genomes had high completeness scores, indicating that the small genome sizes were not the result of sequencing error [27]. Genome statistics for both isolates are listed in Table 2. The 16S rRNA gene sequences of 22-AL-CL-001 and USDA-ARS-USMARC-56711 were compared using blast to NCBI’s nucleotide database, as well as a database containing 16S rRNA gene sequences from type strains of all validly published Pseudomonas species for which sequence data were available at the time of analysis (LPSN, n=329) [1]. The two isolates were found to share the highest level of 16S rRNA sequence similarity to each other, at 99.93% identity (Table 3). Both isolates shared the second-highest percent identity to Pseudomonas parafulva (strain DSM 17004^T^, 16S rRNA sequence accession number AB060132.1; 22-AL-CL-001, 99.72%; USDA-ARS-MARC-56711, 99.73%).
Phylogenetic analysis based on multi-locus sequence data
The P. syringae species complex (Pssc) is a large phylogenetic group that contains many plant-pathogenic and plant-associated Pseudomonas species, including P. syringae, the most common causal agent of BLS, and several additional Pseudomonas species associated with leaf spot diseases of cucurbit crops [2128]. This complex has been divided into 13 phylogroups based on MLSA using the four housekeeping genes cts (gltA), rpoD, gapA and gyrB [2829]. To determine the phylogenetic placement of the two isolates from this study within the Pseudomonas genus, and in relation to the Pssc, MLSA based on these four gene sequences was performed using the program AutoMLSA2 (v. 0.8.1) [30]. The two isolates were compared to Pseudomonas strains representing the 13 phylogroups of the Pssc, species to which they shared relatively high 16S rRNA sequence similarity, as well as other Pseudomonas species also known to infect both plant and animal hosts. Analysis was performed using reference genomes obtained from NCBI and gene sequences from the Plant Associated and Environmental Microbes Database (pamdb.org). A maximum likelihood phylogenetic tree was constructed using IQ-TREE (v. 2.1.3) [3132] and visualized using ITOL (v. 6.9) [33] and Adobe Illustrator (Fig. 1). In this phylogeny, isolates 22-AL-CL-001 and USDA-ARS-USMARC-56711 were placed into a single clade, which was located outside of the Pssc and was distant from all other Pseudomonas reference strains included in the analysis. These results show that the novel isolates are phylogenetically distant from both the Pssc, which contains Pseudomonas species typically associated with leafspot diseases of cucurbits, as well as other Pseudomonas species known to infect both plant and animal hosts.
Maximum likelihood phylogeny inference of P. alabamensis and reference strains, including the most closely related species as identified by genetic comparisons, as well as representatives of P. syringae phylogroups and species known to inhabit both plant and animal hosts, based on concatenated alignments of the housekeeping genes gltA, rpoD, gapA and gyrB. Bootstrap values based on 100 replicates are indicated at branching points. Bolded numbers designate Pssc phylogroups.
Whole-genome comparisons
ANIb and isDDH analyses were conducted using whole genomes of isolates 22-AL-CL-001 and USDA-ARS-USMARC-56711 as well as type strains of Pseudomonas species to which they had high 16S rRNA sequence similarity or relative phylogenetic proximity. The established threshold for species determination is considered to be greater than or equal to 95% for ANI analysis and greater than or equal to 70% for DDH [34]. ANIb analysis was performed using the Python module pyani (v. 0.2.12) [35], while isDDH was conducted using the web tool Genome-Genome Distance Calculator (v. 3.0, formula 2, http://ggdc.dsmz.de/home.php) [3637]. For both analyses, the two isolates were found to have the highest percent similarity to each other with an ANIb score of 97.25% and an isDDH score of 78.5%, identifying them as members of the same species (Table 3). Neither isolate was found to share greater than 95% ANIb or greater than 70% isDDH to any reference strain tested, suggesting that they belong to a currently undescribed species of Pseudomonas. Genetically, the most closely related Pseudomonas species to the two isolates, as determined by ANIb and isDDH, were still relatively distant. P. monteilii (type strain DSM 14164^T^) was the top ANIb match to both 22-AL-Cl-001 and USDA-ARS-USMARC-56711, though comparisons to the isolates produced scores of only 81.2% and 81.21%, respectively. For isDDH, P. oryziphila (type strain 1257^T^) was identified as the closest species match to the isolates, producing similarly low scores of 26.2% (22-AL-CL-001) and 26.4% (USDA-ARS-USMARC-56711).
Comparison to the TYGS
To further verify that the isolates did not belong to any currently described prokaryotic species, the genomes of 22-AL-CL-001 and USDA-ARS-USMARC-56711 were compared to the online Type Genome Server (https://tygs.dsmz.de/) [38]. The TYGS is a high-throughput web tool that compares user-uploaded genomes to a database of prokaryotic type strain genomes using isDDH for species identification. The TYGS was unable to identify either isolate as a member of any species contained within its database, supporting their identities as members of a new species. The results of whole-genome and 16S rRNA gene sequence-based phylogenies produced by the TYGS were overall consistent with those of the MLSA phylogeny. These phylogenetic trees are included within the supplementary materials (Figs S1 and S2, available in the online Supplementary Material).
Based on the results of the genetic and phylogenetic analyses described above, we propose the placement of isolates 22-AL-CL-001^T^ and USDA-ARS-USMARC-56711 within a novel species of Pseudomonas, named P. alabamensis after the geographical origin of isolate 22-AL-CL-001^T^ (Alabama, USA).
Physiological and chemotaxonomic analyses
Isolates 22-AL-CL-001^T^ and USDA-ARS-USMARC-56711 were further characterized with biochemical profiling using the Biolog Gen III microplate system. Overnight cultures, grown at 28 °C on Biolog universal growth agar (BUG, Biolog Inc.), were used to create bacterial suspensions of the recommended level of turbidity in Biolog inoculation fluid. For each isolate, a microplate was then inoculated with 100 µl of suspension per well and incubated at 28 °C for 24 h, after which, test results were recorded and compared to the Biolog database (Table S2). Biolog software identified both isolates as members of the genus Pseudomonas, though it was unable to further identify either as a member of any existing Pseudomonas species contained within its database (MicroLog M System, v. 5.1.1). Of the species contained within the Biolog database, P. fluorescens (22-AL-Cl-001^T^) and Pseudomonas plecoglossicida (USDA-ARS-USMARC-56711) were identified as the closest matches to the novel isolates. Comparison of the biochemical profile of isolate 22-AL-CL-001^T^ to that of P. fluorescens produced a similarity score of 0.157, while comparison of isolate USDA-ARS-USMARC-56711’s profile to that of P. plecoglossicida produced a similarity score of 0.206, with both scores being significantly lower than the accepted value of ≥0.5 necessary for species determination. Carbon utilization data for * P. monteilii*, the most closely related species according to ANIb analysis, were also obtained and compared to Biolog profiles of * P. alabamensis* strains, revealing differences in utilization patterns between the two species (Table S2) [39].
Isolate 22-AL-CL-001^T^, which was designated as the type strain of P. alabamensis sp. nov., was further analysed using MALDI-TOF MS performed by Charles River Laboratories (Newark, DE, USA). Resulting spectra were compared to Bruker Biotyper (v. 11758) and Charles River (v. 23.01) spectral reference libraries for species identification. Charles River Laboratories bases probable species identification on a similarity score between a tested isolate and a reference strain of greater than or equal to 1.75. Isolate 22-AL-CL-001^T^ did not produce a similarity score within this range to any examined reference strain and, so, was unable to be identified as any existing bacterial species based on MALDI-TOF MS.
Bacterial cells of isolate 22-AL-CL-001^T^ were imaged using transmission electron microscopy, conducted by the University of Florida’s Interdisciplinary Center for Biotechnology Research (ICBR), using a Tecnai G2 Spirit TWIN 120 kV microscope. Cells were observed to be rod-shaped, slightly rounded, and to possess one or more polar flagella (Fig. 2).
Images of (a) monoflagellate and (b) multiflagellate cells of P. alabamensis 22-AL-CL-001T as captured by a Tecnai G2 Spirit TWIN 120 kV transmission electron microscope, University of Florida’s ICBR.
Comparison to the genome taxonomy database
To identify whether any additional strains of P. alabamensis sp. nov had been previously sequenced and made publicly available online, the classification of isolates 22-AL-CL-001^T^ and USDA-ARS-USMARC-56711 within the Genome Taxonomy Database was determined [40]. The GTDB is a database that imports prokaryotic genome data from the NCBI Assembly database and organizes them into ‘species clusters’ on the basis of average nucleotide identity, allowing for the identification of genetically similar isolates. Perusal of the database revealed that both P. alabamensis isolates identified in this study had been placed within the GTDB species cluster Pseudomonas_E monteilii_A, alongside seven additional genomes, including six classified within NCBI as P. monteilii and one as P. entomophila. The type strains of these two species (P. monteilii: NBRC 103158^T^=DSM 14164^T^; P. entomophila: L48^T^) did not appear within the Pseudomonas_E monteilii_A cluster and were instead sorted by GTDB into two separate species clusters, Pseudomonas_E monteilii and Pseudomonas_E entomophila, respectively. USDA-ARS-USMARC-56711 had been declared by GTDB as the representative strain of species cluster Pseudomonas_E monteilii_A. Results of ANI, isDDH and MLSA comparisons of the isolates belonging to Pseudomonas_E monteilii_A, as well as type strains of P. monteilii and P. entomophila, are included within the supplementary materials (Table S1, Fig. S3). Both ANI and isDDH analyses revealed that the seven additional isolates belong to P. alabamensis, with an average ANI to type strain 22-AL-CL-001^T^ of 97.57% and an average isDDH score of 78.85%. None of these seven isolates shared ANI or isDDH values above the established thresholds for species identification to the type strains of P. monteilii or P. entomophila. Further inspection of the NCBI assembly records of the seven additional isolates revealed that taxonomy checks performed by NCBI had produced inconclusive results, supporting the conclusion that these organisms had been previously described as members of P. entomophila or P. monteilii in error. Additionally, all seven isolates were found to have relatively short genomes (average=4.60 Mb), similar in size to those of 22-AL-CL-001^T^ and USDA-ARS-USMARC-56711.
MLSA was performed as previously described and included strains contained within GTDB species cluster Pseudomonas_E monteilii_A, type strain genomes of P. monteilii and P. entomophila, and the ten Pseudomonas species determined to have the highest percent ANI to P. alabamensis type strain 22-AL-CL-001^T^, with P. syringae type strain ICMP 3023^T^ included as an outgroup. The results of the MLSA phylogeny were consistent with those of the ANI and isDDH analyses and showed that isolates 22-AL-CL-001^T^ and USDA-ARS-USMARC-56711 clustered with the seven additional isolates placed within the Pseudomonas_E monteilii_A species group by the GTDB and that this group was phylogenetically distinct from all other Pseudomonas species used in the analysis, including the type strains of P. monteilii and P. entomophila.
NCBI metadata associated with the genomes of the additional seven GTDB isolates show that they had been collected within a similar time frame to isolates 22-AL-CL-001^T^ and USDA-ARS-USMARC-56711, between 2017 and 2022, though from different sources, including from metal and other unspecified environmental sources. All seven of the additional isolates had also been collected from the USA, specifically, from the states of New York and North Carolina (Table S1).
Conclusion
In conclusion, we propose that isolates 22-AL-CL-001^T^ and USDA-ARS-USMARC-56711 represent a novel bacterial species, designated P. alabamensis, based on 16S rRNA gene sequence, ANIb and isDDH analyses, including comparison to the TYGS; phylogenies based on MLSA, 16S rRNA gene sequence analysis and whole-genome comparisons; biochemical profiling with the Biolog Gen III microplate system; and MALDI-TOF MS analysis. This species is notable as an example of a pseudomonad that has been isolated from both plant and animal hosts.
Description of Pseudomonas alabamensis sp. nov.
Pseudomonas alabamensis (al.a.bam.en’sis. N.L. fem. adj. alabamensis, pertaining to the geographical location of isolation of type strain 22-AL-CL-001^T^, Alabama, USA).
Cells are Gram-negative motile rods (1.9–2.3 µm in length and 1.1–1.5 µm in width) with one or more polar flagella. Cells are slightly rounder than is typical of Pseudomonas species. Colonies, when grown on NA at 28 °C for 48 h, are bright yellow in color, glossy, round and ~3.0 mm in diameter. Isolates produce a diffusible fluorescent pigment when grown on King’s medium B; are negative for levan production, oxidase activity, pectolytic activity on potato and arginine dihydrolase activity; and do not elicit an HR in either tobacco or tomato plants. Isolates were not observed to cause disease when spray-inoculated on cantaloupe (Cucumis melo), watermelon (Citrullus lanatus) and squash (Cucurbita pepo subsp. pepo) seedlings. Isolates were observed to grow at pH levels of 5 and 6 as well as at salinity levels of 1%, 4% and 8% NaCl. Growth of isolate 22-Al-Cl-001^T^ suspended in tryptic soy broth (Sigma-Aldrich) was assessed at incubation temperatures of 0, 4, 15, 21, 28, 32, 37 and 41 °C. After 48 h of incubation, growth was observed only in cultures incubated at temperatures between 15 and 37 °C, with optimal growth occurring between 21 and 32 °C. Type strain 22-AL-CL-001^T^ (NCPPB 4760=LMG 33363) was isolated from cantaloupe foliage displaying marginal leaf necrosis and blighting in Alabama, USA. Isolate 22-AL-CL-001^T^ has a genome size of 4.66 Mb and a G+C content of 64.5 mol%. NCBI accession numbers for P. alabamensis genomes are as follows: 22-AL-Cl-001^T^, GCA_030580815.1; USDA-ARS-USMARC-56711, GCA_001534745.1. Accession numbers for 16S rRNA gene sequences are: 22-AL-Cl-001^T^, OR725053.1; USDA-ARS-USMARC-56711, PQ115154.1.
Supplementary material
10.1099/ijsem.0.006848Uncited Fig. S1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Parte AC Sardà Carbasse J Meier-Kolthoff JP Reimer LC Göker M List of prokaryotic names with standing in nomenclature (LPSN) moves to the DSMZ Int J Syst Evol Microbiol 2020705607561210.1099/ijsem.0.00433232701423 PMC 7723251 · doi ↗ · pubmed ↗
- 2Spiers AJ Buckling A Rainey PB The causes of Pseudomonas diversity Microbiology 20001462345235010.1099/00221287-146-10-234511021911 · doi ↗ · pubmed ↗
- 3Bergan T Human and animal-pathogenic members of the genus Pseudomonas Starr MP Stolp H Trüper HG Balows A Schlegel HG The Prokaryotes: A Handbook on Habitats, Isolation, and Identification of Bacteria Berlin Springer 1981666700
- 4Schroth MN Hildebrand DC Panopoulos N Phytopathogenic pseudomonads and related plant-associated pseudomonads Dworkin M Falkow S Rosenberg E Schleifer K-H Stackebrandt E The Prokaryotes: A Handbook on the Biology of Bacteria New York Springer 2006714740
- 5Moore ERB Tindall BJ Martins Dos Santos VAP Pieper DH Ramos J-L et al Nonmedical: Pseudomonas Dworkin M Falkow S Rosenberg E Schleifer K-H Stackebrandt E The Prokaryotes Singapore Springer 2006646703
- 6Kodama K Kimura N Komagata K Two new species of Pseudomonas: P. oryzihabitans isolated from rice paddy and clinical specimens and P. luteola isolated from clinical specimens Int J Syst Bacteriol 19853546747410.1099/00207713-35-4-467 · doi ↗
- 7Dussart-Baptista L Bodilis J Barray S Frébourg N Fournier M et al Recurrent recovery of Pseudomonas oryzihabitans strains in a karstified chalk aquifer Water Res 20074111111710.1016/j.watres.2006.10.00717097713 · doi ↗ · pubmed ↗
- 8Schroth MN Cho JJ Green SK Kominos SD Microbiology society publishing. Epidemiology of Pseudomonas aeruginosa in agricultural areas J Med Microbiol 2018671191120110.1099/jmm.0.00075830067169 · doi ↗ · pubmed ↗