The impact of zinc supplementation on carbapenem MICs among bacteria expressing IMP metallo-beta-lactamase
Susan V. Grooters, Dixie F. Mollenkopf, Gregory A. Ballash, Thomas E. Wittum

TL;DR
This study investigates whether adding zinc affects the accuracy of antibiotic resistance tests for bacteria that produce a specific enzyme linked to antibiotic resistance.
Contribution
The study reveals that adding zinc does not improve carbapenem MIC accuracy for IMP-producing bacteria.
Findings
Morganellaceae showed higher imipenem MICs compared to other blaIMP-harboring bacteria.
Supplemental ZnSO4 had no effect on carbapenem MICs for IMP-expressing environmental bacteria.
Zinc supplementation does not enhance the accuracy of MIC testing for IMP metallo-beta-lactamase.
Abstract
Antibiotic-resistant infections cause an estimated 2.8 million illnesses and 35,900 deaths annually in the USA. Carbapenems are a class of antibiotics that are generally reserved to treat life-threatening invasive infections including sepsis. Accurate diagnosis of carbapenem-resistant infections is critical for early and appropriate treatment. blaIMP encodes bacterial production of the IMP metallo-beta-lactamase (MBL), which can confer resistance to all the beta-lactams including carbapenems. Zinc is an essential co-factor in the IMP MBL enzymatic hydrolysis of carbapenems. Tests for the presence of IMP carbapenemase, such as the Carba NP, include zinc sulphate (ZnSO4) although broth dilution methods for determining MIC for carbapenems may vary. We hypothesized that ZnSO4 availability would improve the accuracy of carbapenem MIC determination for bacteria expressing blaIMP. Thus, the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
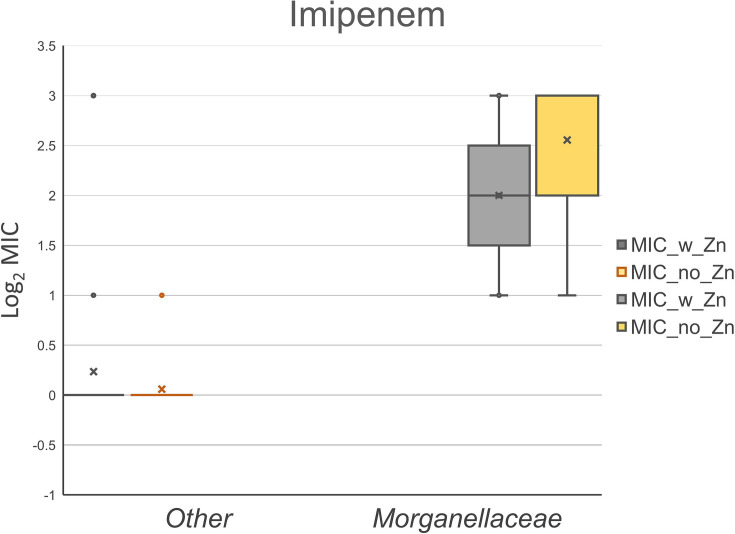 Fig. 1
Fig. 1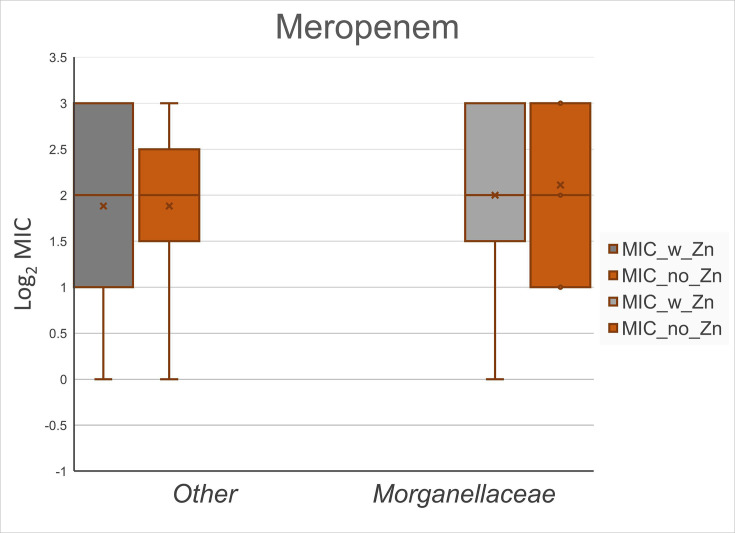 Fig. 2
Fig. 2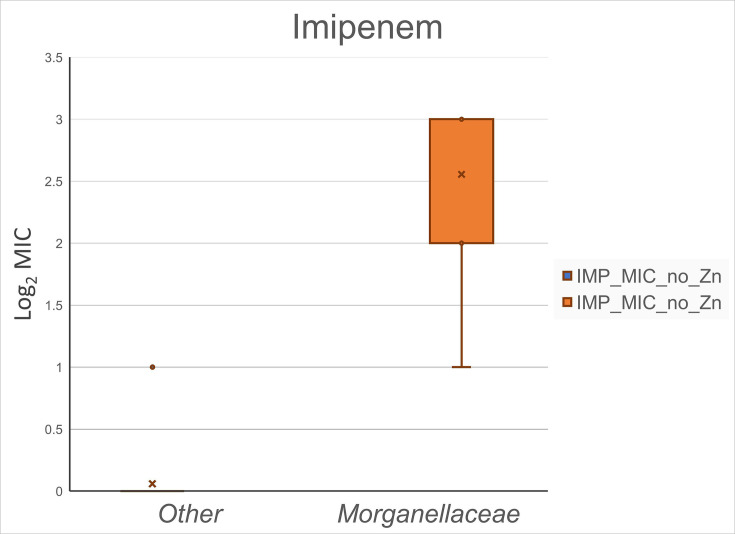 Fig. 3
Fig. 3| Isolate | Taxonomic family |
| MICIPM | MICIPM ( | MICMEM | MICMEM ( |
|---|---|---|---|---|---|---|
|
|
|
|
|
| 2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| 2 |
|
|
|
|
|
|
|
| 2 | 2 |
|
|
|
| 2 |
|
|
|
|
|
|
| ≤1 | ≤1 |
|
|
|
|
|
| ≤1 | ≤1 |
|
|
|
|
|
| ≤1 | ≤1 |
|
|
|
|
|
| ≤1 | ≤1 |
|
|
|
|
|
| ≤1 |
|
|
|
|
|
|
| ≤1 | ≤1 | 2 | 2 |
|
|
|
| ≤1 | ≤1 |
|
|
|
|
|
| ≤1 | ≤1 |
|
|
|
|
|
| ≤1 | ≤1 | ≤1 | ≤1 |
|
|
|
| ≤1 | ≤1 | 2 | 2 |
|
|
|
| 2 | 2 | 8 | 8 |
|
|
|
| ≤1 | ≤1 | 4 | 4 |
|
|
|
| ≤1 | ≤1 | 4 | 4 |
|
|
|
| ≤1 | ≤1 | 4 | 4 |
|
|
|
| ≤1 | ≤1 | 4 | 2 |
|
|
|
| ≤1 | ≤1 | ≤1 | ≤1 |
|
|
|
| ≤1 | ≤1 |
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Infections and bacterial resistance · Antimicrobial Resistance in Staphylococcus
Data Summary
Sequence reads have been deposited in the NCBI Sequence Read Archive and are available under BioProject accession PRJNA1197433. Individual sequence accession numbers include SAMN45808700, SAMN45808699, SAMN45808692, SAMN45808698, SAMN45808672, SAMN45808671, SAMN45808680, SAMN45808675, SAMN45808685 and SAMN45808687.
Introduction
In the USA, antibiotic-resistant infections cause an estimated 2.8 million illnesses and 35,900 deaths annually [1]. Of those illnesses, ~9,000 are estimated to be from carbapenem-resistant bacterial infections [1]. The carbapenem antibiotics are generally reserved to treat life-threatening invasive infections including sepsis. Acquired bacterial resistance to carbapenems is often the result of transmissible carbapenemase genes. One such gene, blaIMP, encodes for the production of the IMP carbapenemase, which can confer reduced susceptibility or resistance to carbapenems and other beta-lactam antibiotics. Bacterial carbapenemase genes are readily transferred horizontally within and between bacterial taxa on mobile genetic elements such as plasmids and transposons [2].
Carbapenemases are classified as serine-beta-lactamases or metallo-beta-lactamases (MBLs) [3]. Class B MBLs can catalyse the hydrolysis of a wide range of beta-lactam antibiotics, including carbapenems [4]. IMP carbapenemases are a subgroup of acquired MBLs of the B1 subclass, with strictly conserved zinc ligand residues and zinc-binding motifs that distinguish them from the other B subclasses [5]. B1 enzymes are most active with two zinc ions bound in the active site and have been shown to have a broad-spectrum substrate profile [5]. blaIMP encodes for the production of a B1 MBL, which can confer reduced susceptibility or resistance to carbapenems and other beta-lactam antibiotics. Zinc is an essential co-factor in the MBL enzymatic hydrolysis of the beta-lactam ring of carbapenems, which is why zinc sulphate (ZnSO_4_) is included in the tests for carbapenemase production, including the Carba NP [6].
The impact of differing levels of zinc availability on in vitro and in cell inhibition of carbapenemase activity by chelation or replacement among clinically relevant MBLs has been reported [7]. Others have indicated that zinc concentration in media is not a driver of allele mutation or increased cell resistance to carbapenems [8]. This suggests that adequate zinc is still essential for carbapenemase activity, particularly for IMP compared with other MBLs, which have adapted to zinc-limited environments [7].
However, an additional source of zinc is not standardized in media utilized for carbapenem MIC determination [9]. If a minimum threshold of available zinc is required for effective hydrolysis of carbapenems by IMP B1 MBLs, then inadequate available zinc in media used for MIC determination of Enterobacterales or other bacteria might result in MIC values that are incorrectly low and could inappropriately influence their clinical interpretation. Therefore, the addition of available ZnSO_4_ might improve the accuracy of carbapenem MIC estimation for bacteria producing the IMP MBL.
The impact of zinc concentration in Mueller–Hinton (MH) broth on MICs of MBL-producing Enterobacteriaceae has been previously reported [9]. They observed zinc variability among commercial lots of MH broth, resulting in different classifications of meropenem susceptibility among multiple MBL genotypes. They concluded that a consensus on the appropriate amount of zinc in culture media is needed to ensure that variations in the zinc content of commercial media do not significantly influence MBL antimicrobial susceptibility test results [9].
In our laboratory, we have previously observed that different taxonomic families of bacteria, all harbouring the class 2 integron and blaIMP gene cassette, can have different carbapenem resistance phenotypes. We have reported that species within the family of Enterobacteriaceae isolates expressing blaIMP appear fully susceptible to imipenem, while species within the Morganellaceae taxonomic family expressing blaIMP appear to have reduced susceptibility [10]. This finding was in contrast to MIC data for Enterobacteriaceae and Morganellaceae producing NDM MBLs, which produce similar carbapenem resistance phenotypes. We hypothesized that these observed differences in carbapenem MICs among bacteria expressing blaIMP could be due to a lack of available exogenous zinc. There is relatively limited scientific characterization of bacterial isolates expressing blaIMP and the specific alleles that are included in this study, IMP-95, IMP-64 and IMP-27, for which limited MIC values are available [2].
We hypothesized that supplemental ZnSO_4_ in media will improve the accuracy of carbapenem MIC determination for bacteria expressing blaIMP and that variability in phenotypic imipenem susceptibility among taxonomic families is attributable to differences in IMP carbapenemase requirements for exogenous zinc availability. Therefore, the objectives of this study are to document the role of exogenous ZnSO_4_ availability on phenotypic imipenem susceptibility of Morganellaceae, Enterobacteriaceae, Shewanellaceae and Moraxellaceae isolates producing the IMP MBL.
Methods
A total of 25 bacterial isolates were utilized in this study that originated from environmental samples: on farms, wastewater treatment plants and surface waters. Selective media used in the initial screening for isolates showed reduced susceptibility of the isolates to meropenem using methods that have been previously described [1012]. Isolates utilized for this study were all confirmed to produce carbapenemase using the Carba NP test [6]. These isolates represent a relatively rare genotype in the USA and were collected over time beginning in 2016 through 2021, some of which have been previously reported [1013].
Isolates in this study phenotypically express reduced susceptibility to carbapenems, and bacterial carriage of blaIMP was confirmed by PCR and Sanger sequencing of amplicons. Taxonomic family and genus were identified using MALDI-TOF (Bruker Scientific, LLC, Billerica, MA) and sequence-based bioinformatic tools for 17 isolates for which whole-genome sequence data were available [1415]. The 25 study isolates included 10 Enterobacteriaceae, 8 Morganellaceae, 5 Shewanellaceae and 2 Moraxellaceae and were presented by taxonomic family rather than species to be consistent with phenotypic distributions that we previously observed [10]. Sequence data were compared with online databases [1619] to determine the index number of the blaIMP gene allele (Table 1).
MICs were determined for each isolate using the Sensititre broth microdilution system (Thermo Fisher Scientific, Oakwood Village, OH) following the Clinical and Laboratory Standards Institute (CLSI) guidelines [2021]. Isolates were processed in duplicate, with and without added ZnSO_4_ at 0.1 mmol l^−1^, the same concentration of zinc present in the Carba NP test [6], in cation-adjusted MH broth (Thermo Fisher Scientific) suspensions to a 0.5 McFarland standard. While we did not measure the zinc concentration of the media and it is not reported by the manufacturer, reports from the literature of zinc concentrations ranging from 0.01 to 0.1 mmol l^−1^ in commercial media suggest that this amount of added ZnSO_4_ will provide significant free zinc available to the cell [2223]. Standard carbapenem-resistant Enterobacterales and extended-spectrum beta-lactamase Sensititre MIC panels CMV3AGNF and GNX2F, respectively (Thermo Fisher Scientific), were utilized to produce paired MIC results for each isolate.
ZnSO_4_ enriched and non-enriched pairs’ MIC data were log_2_ transformed for analysis and compared using the Wilcoxon signed-rank test in Stata IC13 (StataCorp LP, College Station, TX, USA) for paired data. This test evaluated the null hypothesis that the median difference among MIC pairs was zero. The Kruskal–Wallis test in Stata IC13 (StataCorp LP) was used to assess the null hypothesis that there was no difference in MIC between taxonomic groups.
Results
Isolate MICs against imipenem and meropenem, with and without ZnSO_4_ added to the media, are presented in Table 1. We observed no difference in carbapenem MIC values based on supplemental ZnSO_4_ included in the media. This observation was similar for both imipenem MIC (Fig. 1) and meropenem MIC (Fig. 2). A unit of increase in the figures corresponds to a doubling of the MIC value, as graphical MIC values are log_2_ transformed. As expected, serine carbapenemase-producing control isolates were not impacted by additional zinc, and MIC values were consistent for three control isolates expressing blaKPC with and without additional zinc for imipenem and meropenem (data not shown). Morganellaceae imipenem MIC without additional ZnSO_4_ present was higher (P<0.001) than other taxonomic families (Fig. 3), which is consistent with expected intrinsic carbapenem resistance [21].
Observed MIC values against imipenem for 8 Morganellaceae and 17 other taxonomic family isolates (10 Enterobacteriaceae, 5 Shewanellaceae and 2 Moraxellaceae) expressing IMP carbapenemase with and without supplemental ZnSO4. The supplemental ZnSO4 did not impact imipenem MIC (P>0.05).
Observed MIC values against meropenem for 8 Morganellaceae and 17 other taxonomic family isolates (10 Enterobacteriaceae, 5 Shewanellaceae and 2 Moraxellaceae) expressing IMP carbapenemase with and without supplemental ZnSO4. The supplemental ZnSO4 did not impact meropenem MIC (P>0.05).
Observed MIC values against imipenem for 8 Morganellaceae and 17 other taxonomic family isolates (10 Enterobacteriaceae, 5 Shewanellaceae and 2 Moraxellaceae) expressing IMP carbapenemase without supplemental ZnSO4. Morganellaceae isolates had higher (P<0.001) MIC values than the other taxonomic family isolates.
Discussion
We found no difference in paired carbapenem MIC values with the addition of ZnSO_4_ to the media for the inoculation of Sensititre microdilution panels. Therefore, we conclude that supplemental ZnSO_4_ does not influence the phenotypic presentation of the enzymatic activity of these MBL blaIMP gene alleles. Our results suggest that there is adequate available zinc for effective IMP MBL hydrolysis of carbapenems in the current guidelines for MIC determination, and there is no need to recommend supplemental zinc in the media for isolates expressing blaIMP.
Available zinc in media can vary and impact the interpretation of enzymatic efficiencies [9,2224]. It has been previously reported that variability in zinc concentration in commercial MH media can impact carbapenem MICs of Enterobacteriaceae that produce MBLs, indicating a need for standardization of zinc levels in media [9]. They observed that media with inadequate zinc resulted in higher meropenem MICs, with enough variability to influence the clinical interpretation [9]. We did not measure the zinc concentration of the MH broth that we used for this study, and it was not reported by the manufacturer, but our results indicate that zinc was not limiting for determining carbapenem MICs. This suggests that beyond a threshold concentration of available zinc, there is no additional benefit of supplemental zinc for the determination of carbapenem MICs in strains that produce IMP carbapenemase.
It is possible that intrinsic resistance and not carriage of blaIMP has the more important impact on our observed differences in the MIC results. Intrinsic antimicrobial resistance is expected in some WT bacterial isolates and is therefore representative of the species. CLSI notes that Morganellaceae, Proteus spp., Providencia spp., and Morganella spp. may have intrinsic resistance, and thus elevated MICs to imipenem that make the wild-type of the species appear resistant [21]. CLSI further notes that this is likely from mechanisms other than the production of carbapenemase [21].
The overall observed difference in MIC to imipenem compared with meropenem for Enterobacteriaceae, Shewanellaceae and Moraxellaceae may indicate that meropenem is a stronger substrate, regardless of additionally available zinc (Table 1). The similarity of imipenem and meropenem MICs in Morganellaceae suggests that it is intrinsic resistance and not carriage of blaIMP that produces the phenotypic results. Such intrinsic resistance to imipenem could be from reduced membrane permeability as has been noted in previous studies [25] or other differences unique to the Morganellaceae taxonomic family.
Others have reported meropenem to be a better substrate for IMP-27, as it is considered part of the IMP-6-like alleles and has a glycine aa residue substitute for serine, at position 262, compared with IMP-1 [26]. However, the studies indicating increases in MIC for meropenem compared with imipenem (among S262G alleles, i.e. IMP-6 and IMP-27) did not mention whether Morganellaceae were included in the analysis [26]. That IMP-27 and IMP-64 are both found in the Morganellaceae family in this study (and differ by only one SNP), their increase in imipenem resistance could be due to intrinsic resistance, other than the presence of the carbapenemase alone. Our sequenced Morganellaceae isolates did not harbour any other known carbapenemase genes, which could also contribute to reduced susceptibility to imipenem.
A study by Segawa et al. also examined IMP-6, where reduction in MICs to imipenem compared with meropenem was evidenced, but the IMP-6 were mostly in Escherichia coli and some Klebsiella spp. although the authors did not indicate a species-level association with MIC differences [27]. This is despite positive Carba NP tests for IMP-6 carbapenemase production, which was not accurately reflected in the MIC results for imipenem, but did correlate with the activity of the enzymatic hydrolysis [27].
Other resistance mechanisms could be influencing the observed MICs to imipenem and meropenem in Morganellaceae, including reduced outer membrane permeability [25]. For example, mutations resulting in reduced permeability of the ImpR outer membrane porin of Proteus mirabilis resulted in decreased susceptibility to carbapenems [28]. Morganellaceae may also exhibit intrinsic resistance to carbapenems using a combination of DHA-like beta-lactamase production and low target affinity [29]. However, these intrinsic mechanisms are likely not fully effective, which may lead these strains to sometimes acquire mobile carbapenemase genes including blaIMP. Our data are consistent with what has been reported by laboratory standard setting organizations (e.g. CLSI), indicating that Morganellaceae have intrinsic resistance to imipenem, independent of acquired carbapenemase-mediated resistance, which appears to be independent of supplemental zinc availability. A complete understanding of the mechanisms of intrinsic and acquired resistance in potentially pathogenic bacteria is critical to appropriate antimicrobial treatment recommendations and effective antimicrobial stewardship. Our results suggest the need for continued research in this area.
We have also potentially identified a substrate preference for the blaIMP alleles for meropenem over imipenem among non-Morganellaceae isolates, supporting findings reported for Enterobacteriaceae by others for blaIMP-6 and blaIMP-6-like alleles [22627].
However, the phenotypic results we observed may not be indicative of the level of bacterial blaIMP gene expression. Meini et al. [26] suggested that MIC differences in substrate preferences, reported by Yano et al. [23] and Liu et al. [22], could be attributed to variations in enzymatic assays. In particular, the differences in zinc concentration in the two studies (0.01 mmol ZnCl_2_ versus 0.1 mmol ZnSO_4_, respectively) could contribute to differences in catalytic efficiencies of meropenem hydrolysis of the blaIMP under differing zinc concentrations [222326]. In MBLs including IMP, the required availability of the metal co-factor, zinc, is critical for the enzyme to be fully active in vivo, particularly during the time of folding, which occurs after translocation from the cytoplasmic membrane to the periplasm [3032]. Some experiments have shown that for MBLs to be active, they are dependent upon the concentration of extracellular zinc, as periplasmic zinc availability is not well regulated and is determined by extracellular concentrations [7]. Extracellular zinc is an important consideration in times of infection when host immune responses may restrict the availability of this important micronutrient [7]. Adding to the complexity of understanding the bacterial phenotypic expression of antimicrobial resistance and the availability of extracellular zinc, commercially available cation-adjusted MH broth varies in zinc cation concentration enough to impact the interpretation of MIC values for MBLs [924]. Also complicating the need for better in vitro testing to better approximate in vivo enzymatic reactions, most dilutions in cation-adjusted media may exceed physiological concentrations of zinc in patients [7]. Therefore, current phenotypic testing likely cannot adequately predict the clinical impacts of MBLs [7], especially those from taxonomic families other than Morganellaceae that may be intrinsically susceptible to imipenem.
Our results are directly applicable to research laboratories testing MICs of environmental isolates producing IMP carbapenemase. However, this result also suggests that supplemental zinc may be unnecessary for MIC testing of clinical isolates when cation-adjusted MH broth has adequate available zinc. The implication for clinical laboratories, including those where IMP-mediated resistance is emerging, is that standard commercial media are appropriate for determining MICs of isolates expressing blaIMP.
Our results are limited by the small sample size used for this study, but isolates expressing blaIMP are rare and additional isolates were not available for this project. However, it is unlikely that even with a much larger sample size, we would have detected such a small observed difference in MIC values by ZnSO_4_ supplementation. In addition, we utilized only environmental and livestock commensal isolates for this study, which may not be representative of clinical diagnostic isolates. Future research utilizing clinical isolates might provide additional insight into the role of supplemental zinc in the determination of clinical diagnostic MICs. There is also still a need for rapid diagnostic testing to detect the presence of MBLs by bacterial pathogens and for bacterial speciation, as resistance to carbapenems remains a serious public health concern [133].
Our results indicate that the addition of supplemental ZnSO_4_ does not affect the phenotypic expression of carbapenem susceptibility among different bacterial taxa from environmental sources that carry blaIMP. This result indicates that supplemental zinc is unnecessary for MIC determination of bacterial isolates producing IMP carbapenemase. Additional research investigating other potentially important mechanisms influencing the expression of carbapenemase genes including blaIMP and the accurate determination of clinical MICs are needed. Treatment of carbapenem-resistant infections remains critical in sepsis patients, and the availability of accurate clinical diagnostic results is important to appropriate therapeutic decision-making.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Centers for Disease Control and Prevention Antibiotic resistance threats in the United States, 20192019
- 2Bahr G González LJ Vila AJ Metallo-β-lactamases in the age of multidrug resistance: from structure and mechanism to evolution, dissemination, and inhibitor design Chem Rev 20211217957809410.1021/acs.chemrev.1c 0013834129337 PMC 9062786 · doi ↗ · pubmed ↗
- 3Ambler RP The structure of beta-lactamases Philos Trans R Soc Lond B Biol Sci 198028932133110.1098/rstb.1980.00496109327 · doi ↗ · pubmed ↗
- 4Nordmann P Poirel L Epidemiology and diagnostics of carbapenem resistance in Gram-negative bacteria Clin Infect Dis 201969 S 521S 52810.1093/cid/ciz 82431724045 PMC 6853758 · doi ↗ · pubmed ↗
- 5Palzkill T Metallo-β-lactamase structure and function Ann N Y Acad Sci 201312779110410.1111/j.1749-6632.2012.06796.x 23163348 PMC 3970115 · doi ↗ · pubmed ↗
- 6Nordmann P Poirel L Dortet L Rapid detection of carbapenemase-producing Enterobacteriaceae Emerg Infect Dis 2012181503150710.3201/eid 1809.12035522932472 PMC 3437707 · doi ↗ · pubmed ↗
- 7Bahr G González LJ Vila AJ Metallo-β-lactamases and a tug-of-war for the available zinc at the host-pathogen interface Curr Opin Chem Biol 20226610210310.1016/j.cbpa.2021.10210334864439 PMC 8860843 · doi ↗ · pubmed ↗
- 8Cheng Z Bethel CR Thomas PW Shurina BA Alao J-P et al Carbapenem use is driving the evolution of imipenemase 1 variants Antimicrob Agents Chemother 202165 e 01714-2010.1128/AAC.01714-2033468463 PMC 8097420 · doi ↗ · pubmed ↗