Yamadazymaakebiae sp. nov. and Y.hainanensis sp. nov. (Debaryomycetaceae, Saccharomycetales) from leaf in China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
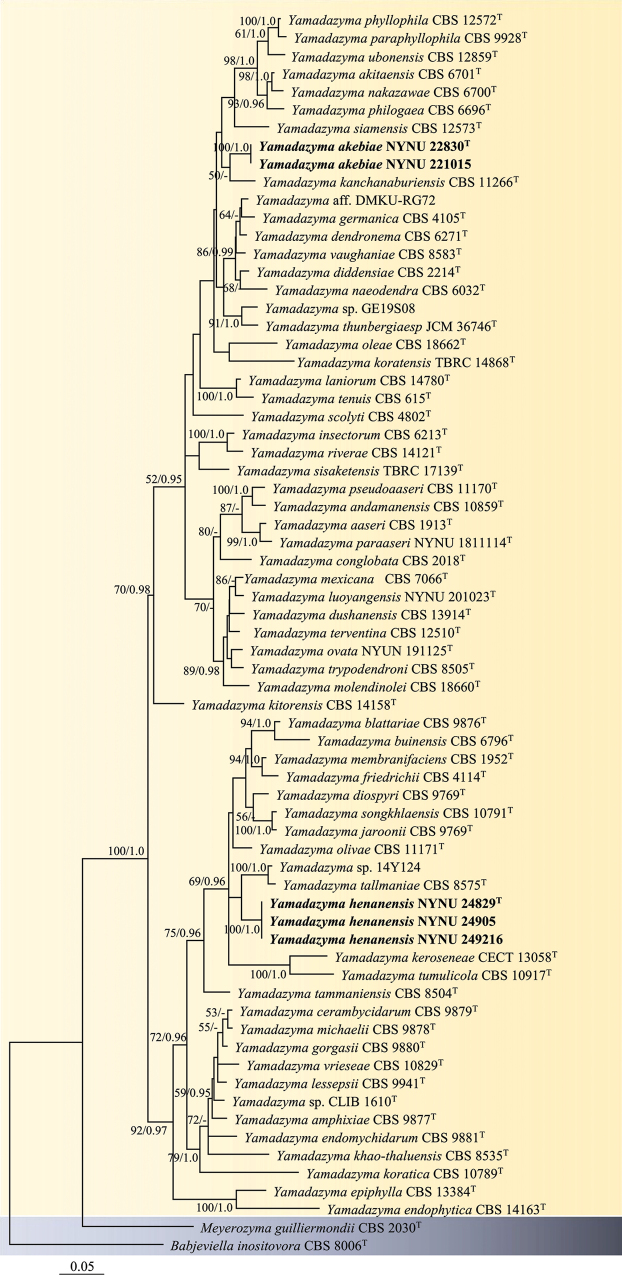 Figure 1
Figure 1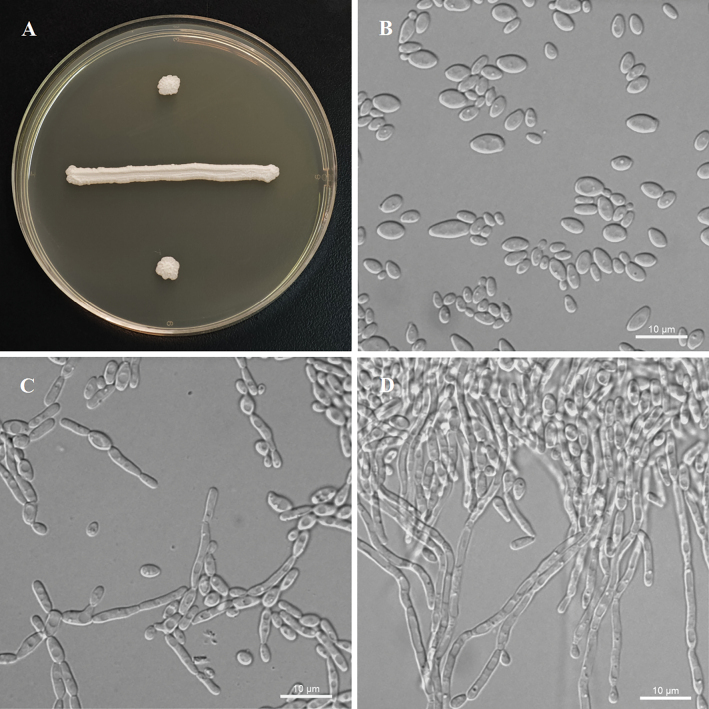 Figure 2
Figure 2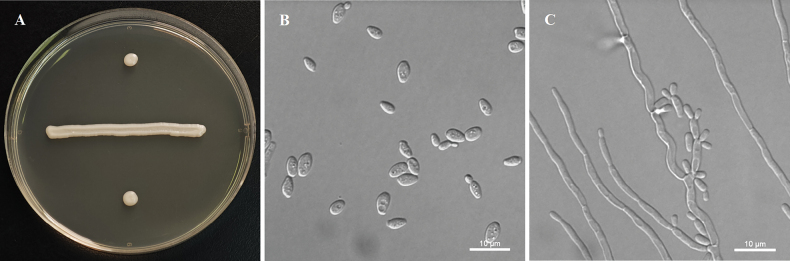 Figure 3
Figure 3| Species | Strain no. | Locality | GenBank accession no. | |
|---|---|---|---|---|
| ITS | LSU D1/D2 | |||
|
| Norway |
|
| |
|
|
|
|
|
|
|
|
|
|
| |
|
| Japan |
|
| |
|
| Panama |
|
| |
|
| Thailand |
|
| |
|
| Panama |
|
| |
|
| Papua New Guinea |
|
| |
|
| Panama |
|
| |
|
| — |
|
| |
|
| South Africa |
|
| |
|
| USA |
|
| |
|
| China |
|
| |
|
| China |
|
| |
|
| Panama |
|
| |
|
| Thailand |
|
| |
|
| Thailand |
|
| |
|
| Germany |
|
| |
|
| Germany |
|
| |
|
| Panama |
|
| |
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
| |
|
| South Africa |
|
| |
|
| Thailand |
|
| |
|
| Thailand |
|
| |
|
| CECT 13058T | UK |
|
|
|
| Thailand |
|
| |
|
| Japan |
|
| |
|
| Thailand |
|
| |
|
| Thailand |
|
| |
|
| USA |
|
| |
|
| Panama |
|
| |
|
| NYNU 201023T | China |
|
|
|
| India |
|
| |
|
| Agria cactus |
|
| |
|
| Italy |
|
| |
|
| Panama |
|
| |
|
| South Africa |
|
| |
|
| Japan |
|
| |
|
| Italy |
|
| |
|
| Greece |
|
| |
|
| NYUN 191125T | China |
|
|
|
| NYNU 1811114T | China |
|
|
|
| Taiwan |
|
| |
|
| South Africa |
|
| |
|
| Thailand |
|
| |
|
| Germany |
|
| |
|
| Brazil |
|
| |
|
| USA |
|
| |
|
| Thailand |
|
| |
|
| Thailand |
|
| |
|
| Thailand |
|
| |
|
| French Guiana |
|
| |
|
| USA |
|
| |
|
| Russia |
|
| |
|
| Italy |
|
| |
|
| JCM 36746T | Japan |
|
|
|
| Canada |
|
| |
|
| Japan |
|
| |
|
| Thailand |
|
| |
|
| French Guiana |
|
| |
|
| Brazil |
|
| |
| CLIB 1610 | France |
|
| |
| 14Y124 | Japan |
|
| |
| GE19S08 | Taiwan |
|
| |
| Thailand |
|
| ||
|
| USA |
|
| |
|
| — |
|
| |
| Characteristics |
|
| ||
|---|---|---|---|---|
| Fermentation of | ||||
| d-Glucose | + | w | – | + |
| Galactose | + | w | – | n |
| Assimilation of | ||||
| Inulin | + | – | w | n |
| Sucrose | + | + | – | + |
| Maltose | + | + | – | + |
| Melezitose | + | + | – | + |
| Methyl-α-D-glucoside | + | + | – | + |
| DL-Lactate | + | – | – | – |
| D-Glucosamine | + | s | – | + |
| Growth tests | ||||
| 10%Nacl/5%glucose | – | n | – | + |
| Vitamin-free medium | + | – | + | n |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Yeasts and Rust Fungi Studies · Mycorrhizal Fungi and Plant Interactions
Introduction
The genus Yamadazyma, belonging to the ascosporogenous yeasts within the family Debaryomycetaceae, was initially established by Billon-Grand (1989) and later delineated through phylogenetic analyses by Kurtzman and Suzuki (2010). Currently, Yamadazyma encompasses 68 recognized species, including 35 anamorphic taxa previously classified within the Yamadazyma clade of the genus Candida (Khunnamwong et al. 2023; Avesani et al. 2024; Seike et al. 2024). Members of this genus are characterized by several shared phenotypic features, such as the synthesis of coenzyme Q-9 (CoQ-9), the ability to form pseudohyphae, sugar fermentation capacity, and a requirement for external vitamins to support growth (Billon-Grand 1989; Kurtzman 2011). Furthermore, seven teleomorphic species—Y.akitaensis, Y.mexicana, Y.nakazawae, Y.philogaea, Y.riverae, Y.scolyti, and Y.triangularis—are known to produce one to four hat-shaped ascospores on sporulation media (Kurtzman 2011; Lopes et al. 2015).
Species of the genus Yamadazyma exhibit a cosmopolitan distribution and have been isolated from a wide array of substrates. These yeasts are frequently recovered from plant-associated materials (Lu et al. 2004; Groenewald et al. 2011; Kaewwichian et al. 2013; Lopes et al. 2015; Maksimova et al. 2020; Gao et al. 2021; Khunnamwong et al. 2023), mushrooms (Nakase et al. 2008; Khunnamwong et al. 2023), various food sources (Nisiotou et al. 2010; Avesani et al. 2024), marine environments (Burgaud et al. 2011, 2016), insects (Suh et al. 2005), soil samples (Seike et al. 2024), estuarine water (Am-In et al. 2011), and even the atmosphere (Kurtzman et al. 2001). Nevertheless, a significant proportion of known species are predominantly associated with decaying wood, insects, and insect frass (Groenewald et al. 2011; Kurtzman 2011; Gao et al. 2021). Notably, the genus also includes clinically important species, such as Y.aaseri, Y.conglobata, Y.mexicana, Y.pseudoaaseri, and Y.triangularis (Kurtzman 2011; Lachance et al. 2011; Al-Sweih et al. 2017; Giachos et al. 2024). In recent years, interest in the ecological roles, biocontrol capacities, and biotechnological applications of Yamadazyma has increased. For instance, Y.cocois, an endophytic yeast, resides within coconut tissues without causing harm to the host plant (Maksimova et al. 2020). Y.mexicana has been shown to suppress phytopathogenic fungi, offering effective control of anthracnose in avocados at both preharvest and postharvest stages (Gonzбlez-Gutiйrrez et al. 2024). A yet undescribed species, referred to as Yamadazyma sp. 1, demonstrated the ability to ferment cellobiose, indicating potential for bioethanol production (Lopes et al. 2018). Moreover, Y.triangularis exhibits probiotic potential and has been utilized to produce novel antioxidant peptides (Cai et al. 2022). These findings underscore the ecological, agricultural, and biotechnological significance of Yamadazyma, highlighting the need for continued and in-depth biodiversity investigations within this genus.
China harbors a high diversity of Yamadazyma species, with current records documenting nine taxa reported from various regions across the country (Gao et al. 2021). The first species described from China was Y.diospyri, isolated from Kaki fruit by Lu et al. (2004). Subsequently, Y.paraphyllophila was identified on phylloplane from Taiwan, China (Kaewwichian et al. 2013). Over the past decade, our research group has conducted extensive taxonomic investigations on Yamadazyma in China, particularly focusing on isolates from decaying wood. These efforts led to the identification of seven species, including three previously known taxa—Y.insectorum, Y.akitaensis, and Y.olivae—and four novel species: Y.dushanensis, Y.luoyangensis, Y.ovata, and Y.paraaseri (Gao et al. 2021). More recently, Y.triangularis, originally described from Japan, was newly recorded in China from dry-cured Xuanwei ham (Cai et al. 2022). Accordingly, China has been identified as a biodiversity hotspot for the discovery of unexpected novel species and newly recorded species of Yamadazyma.
Despite recent progress, the species richness of Yamadazyma in China remains underexplored. Comprehensive surveys encompassing diverse ecosystems and substrates, particularly in under-sampled regions, are essential to uncover the full extent of the genus’s diversity and to enhance our understanding of its ecological and evolutionary significance. In our most recent investigations conducted over the past two years, several new Yamadazyma strains were isolated from plant leaves. Further phenotypic and molecular phylogenetic analysis confirmed that these strains represent two distinct novel species: Yamadazymaakebiae sp. nov. and Y.hainanensis sp. nov. Their formal descriptions are provided herein, contributing to the expanding inventory of Yamadazyma species in China and advancing the taxonomic and ecological framework of the genus.
Materials and methods
Sample collection and yeast isolation
Living leaf samples were collected from two locations in China: the Guiyang Medicinal Botanical Garden, Guizhou Province (26°34'51"N, 106°42'36"E), and Wuzhi Mountain, Hainan Province (18°17'21"N, 109°40'55"E). Yeast strains representing putative novel species were isolated from the leaf surface using the washing and dilution approach outlined by Jiang et al. (2024). Fresh leaves were cut into small segments using sterile scissors and placed into 10 mL sterile centrifuge tubes containing sterile water supplemented with 0.05% Tween 80. The samples were shaken for approximately 10 minutes, after which the suspension was serially diluted to 10^−2^. A volume of 200 μL from each dilution was spread onto yeast extract-malt extract (YM) agar plates (0.3% yeast extract, 0.3% malt extract, 0.5% peptone, 1% glucose, and 2% agar) supplemented with chloramphenicol (200 μg/mL) to inhibit bacterial growth. Plates were incubated at 25 °C for 5–7 days. Emerging yeast colonies were subcultured on fresh YM agar to obtain pure isolates. Purified strains were suspended in 20% (v/v) glycerol and stored at −80 °C for long-term preservation.
Phenotypic characterization
Morphological, physiological, and biochemical characteristics were assessed following standardized protocols outlined by Kurtzman et al. (2011). Colony morphology was evaluated on YM agar after incubation at 25 °C for 7 days. Cellular morphology was observed in YM broth cultures incubated at 25 °C for 3 days using a LEICA DM2500 microscope (LEICA, Wetzlar, Germany) equipped with LAS V4.13 software. To determine the sexual state, single or mixed cultures of each strain were incubated on various sporulation media, including YM agar, 5% malt extract (ME) agar (5% malt extract and 1.5% agar), cornmeal (CM) agar (2% cornmeal and 1.5% agar), Fowell’s acetate agar (0.5% sodium acetate and 2% agar), and Gorodkowa agar (0.1% glucose, 0.5% sodium chloride, 1% peptone and 2% agar). Cultures were maintained at 15 °C and 25 °C for up to six weeks, with observations recorded at two-week intervals (Kurtzman 2011; Gao et al. 2021). Sugar fermentation was examined in liquid medium using Durham fermentation tubes. Carbon and nitrogen assimilation tests were conducted in liquid media, with nitrogen assimilation assessed using starved inocula (Kurtzman et al. 2011). Temperature tolerance was evaluated on YM agar plates at 15, 20, 25, 30, 35, and 37 °C. All newly proposed species names and corresponding descriptions have been deposited in the MycoBank database (https://www.mycobank.org; accessed on 25 February 2025).
DNA extraction, PCR amplification, and sequencing
Genomic DNA was extracted from actively growing yeast cells cultivated on YM agar using the Ezup Column Yeast Genomic DNA Purification Kit, following the manufacturer’s instructions (Sangon Biotech Co., Shanghai, China). The ITS region and the D1/D2 domain of the LSU rRNA gene were amplified using primer pairs ITS1/ITS4 (White et al. 1990) and NL1/NL4 (Kurtzman and Robnett 1998), respectively. PCR amplifications were conducted in a 25 μL reaction mixture containing 9.5 μL of nuclease-free water, 12.5 μL of 2× Taq PCR Master Mix with blue dye (Sangon Biotech Co., Shanghai, China), 1 μL of genomic DNA template, and 1 μL of each primer. The thermal cycling protocol consisted of an initial denaturation at 98 °C for 2 minutes, followed by 35 cycles of denaturation at 98 °C for 10 seconds, annealing at 55 °C for 10 seconds, and extension at 72 °C for 15 seconds, with a final extension at 72 °C for 5 minutes. PCR products were examined by electrophoresis on 1% agarose gels. Amplicons displaying clear single bands were purified and sequenced by Sangon Biotech (Shanghai) Co., Ltd. (Shanghai, China). Forward and reverse sequences were edited and assembled into consensus sequences using BioEdit v7.1.3.0 (Hall 1999). Sequence similarity searches were performed using the BLASTN 2.2.19 algorithm (Zhang et al. 2000) against the GenBank database. All newly generated sequences were submitted to GenBank (https://www.ncbi.nlm.nih.gov/genbank/; Table 1).
Phylogenetic analyses
To infer phylogenetic relationships, newly generated ITS and LSU D1/D2 sequences from five isolates were combined with reference sequences retrieved from the GenBank database (Table 1). Taxon sampling was based on the frameworks established by Avesani et al. (2024) and Seike et al. (2024). Sequence alignment for each region was conducted using MAFFT v. 7.110 with the L-INS-i algorithm, which is optimized for accuracy (Katoh and Standley 2013). Alignments were inspected and edited using BioEdit v7.1.3.0 (Hall 1999), with manual refinements applied to improve character homology across taxa. The final ITS and LSU D1/D2 alignments were concatenated into a single dataset using PhyloSuite v. 1.2.3 (Zhang et al. 2020).
Phylogenetic analyses were performed using maximum likelihood (ML) and Bayesian inference (BI) methods. The most appropriate model of DNA substitution was selected using Modeltest v. 3.04 (Posada and Crandall 1998) with the Akaike information criterion (AIC). The model GTR + I + G was selected for both ML and BI analyses. ML analysis was conducted using RAxML v. 8.2.3 (Stamatakis 2014) with 1,000 bootstrap replicates. BI analysis was conducted using MrBayes v. 3.2.7a (Ronquist et al. 2012) with 50 million generations using the parameter settings described previously (Lu et al. 2024). Each tree was visualized with bootstrap support (BS) values ≥50% and Bayesian posterior probabilities (BPPs) ≥0.95 in FigTree v. 1.4.3 (Andrew 2016).
Abbreviations
GDMCC Guangdong Microbial Culture Collection Center, Guangzhou, PR China;
PYCCPortuguese Yeast Culture Collection, Caparica, Portugal.
Results
Molecular phylogeny
Phylogenetic reconstruction was based on the alignment of the concatenated ITS + LSU D1/D2 dataset, which included 69 ITS and 69 LSU D1/D2 sequences from 69 yeast strains representing 66 species. Phylogenetic trees inferred using both ML and BI methods exhibited similar topologies; therefore, only the tree derived from the ML analysis is presented (Fig. 1). The isolates from Guizhou and Hainan Provinces formed two distinct lineages in the phylogenetic tree (Fig. 1).
Phylogenetic tree of Yamadazyma inferred from maximum likelihood (ML) analysis based on a combined ITS and LSU D1/D2 dataset, with BabjeviellainositovoraCBS 8006T as the outgroup. Bootstrap support (BS) ≥ 50% and Bayesian posterior probabilities (BPPs) ≥ 0.95 are shown. Strains marked with “T” are ex-type. The strains from this study are highlighted in bold.
Strains NYNU 22830 and NYNU 221015, isolated from Guizhou Province, shared identical D1/D2 and ITS sequences, indicating they are conspecific. These strains are closely related to Y.kanchanaburiensisCBS 11266^T^ (Fig. 1), differing by four nucleotide (nt) substitutions (~0.8%) in the D1/D2 domain and 30 nt mismatches (~4.9%) in the ITS region. In comparison with other Yamadazyma species, they exhibit more than 9 nt substitutions (~1.7%) in the D1/D2 domain and 25 nt mismatches (~4%) in the ITS region. Based on these genetic differences, strains NYNU 22830 and NYNU 2487 are proposed as a new species, named Yamadazymaakebiae sp. nov.
Strains NYNU 24829, NYNU 24905, and NYNU 249216, isolated from Hainan Province, had identical D1/D2 and ITS sequences. These strains formed a well-supported clade within the genus Yamadazyma (Fig. 1) and were closely related to Yamadazyma sp. 14Y124 and Y.tallmaniaeCBS 8575^T^, differing by 17–18 nt substitutions (~3.1–3.4%) in the D1/D2 domain and 29–33 nt mismatches (~4.7–5.2%) in the ITS region. Based on these genetic differences, these three strains are proposed as a new species, named Yamadazymahenanensis sp. nov.
Taxonomy
Yamadazyma
akebiae
Taxon classificationFungiSaccharomycetalesDebaryomycetaceae
C.Y. Chai & F.L. Hui sp. nov.
B6EF1452-F7F5-5906-8A3B-9CA7E910ABB2
858833
Type.
China • Guizhou Prov.: Guiyang City, Guiyang Medicinal Botanical Garden, in the phylloplane of Akebiatrifoliata, August 2022, L. Zhang & F.L. Hui, NYNU 22830 (holotypeGDMCC 2.303 preserved as a metabolically inactive state, ex-type PYCC 9930).
Etymology.
The specific epithet akebiae refers to Akebia, the plant genus from which the type strain was isolated.
Description.
After 7 days of growth on YM agar at 25 °C, the colonies appear white to cream-colored, lucid, and convex with irregular surfaces and margins. After 3 days of growth in YM agar at 25 °C, cells are ovoid to ellipsoid (2.5–3.9 × 4.1–7.4 μm) and occur singly or in pairs. Budding is multilateral. Pseudohyphae are formed, but true hyphae are absent in slide culture on cornmeal agar after 7 days at 25 °C. No asci or signs of conjugation are observed on YM agar, 5% ME agar, Fowell’s acetate agar, CM agar, or Gorodkowa agar after 6 weeks at 15 or 25 °C. Glucose and galactose are fermented, while sucrose, maltose, lactose, raffinose, trehalose, and d-xylose are not. Glucose, inulin, sucrose, galactose, trehalose, maltose, melezitose, methyl α-d-glucoside, cellobiose, salicin, l-sorbose, l-rhamnose, d-xylose, l-arabinose, d-arabinose, d-ribose, ethanol, glycerol, erythritol, ribitol, d-mannitol, d-glucitol, dl-lactate, succinate, citrate, d-gluconate, d-glucosamine, N-aetyl-d-glucosamine, and d-glucono-1, 5-lactone (delayed) are assimilated. No growth is observed with raffinose, melibiose, lactose, 5-keto-d-gluconate, methanol, galactitol, myo-inositol, 2-keto-d-gluconate, or d-glucuronate. In nitrogen-assimilation tests, growth is present in ethylamine, l-lysine, and cadaverine, while growth is absent in nitrate, nitrite, and creatine. Growth occurs at 35 °C, but not at 37 °C. No growth is observed in the presence of 10% NaCl with 5% glucose, 0.01% cycloheximide, or 1% acetic acid. Starch-like compounds are not produced. Urease activity and diazonium blue B reactions are negative.
Morphology of Yamadazymaakebiae (ex-type, NYNU 22830). A. Colony on YM agar after 3 days at 25 °C; B. Budding cells on YM agar at 25 °C after 3 days; C. Simple pseudohyphae on CM agar after 3 days at 25 °C; D. Pseudohyphae with blastoconidia on CM agar after 7 days at 25 °C. Scale bars: 10 μm.
Additional isolates examined.
China • Guizhou Province, Guiyang City, Guiyang Medicinal Botanical Garden, in the phylloplane of Sapiumsebiferum, August 2022, L. Zhang & F.L. Hui, strain number: NYNU 221015.
GenBank accession numbers.
Holotype GDMCC 2.303^T^ (ITS: OP566868; LSU D1/D2: OP566866); additional isolate NYNU 221015 (ITS: MZ318445; LSU D1/D2: MZ318422).
Notes.
Physiologically, Y.akebiae sp. nov. differs from its closely related species Y.kanchanaburiensis in its ability to assimilate inulin and dl-lactate and grow in vitamin-free medium (Table 2).
Yamadazyma
hainanensis
Taxon classificationFungiSaccharomycetalesDebaryomycetaceae
C.Y. Chai & F.L. Hui sp. nov.
1926666C-49D0-57AB-BBF1-F7CD9FD80D50
858834
Type.
China • Hainan Prov.: Wuzhishan City, Wuzhi Mountain, in the phylloplane of Daemonoropsmargaritae, August 2024, S.L. Lv, NYNU 24829 (holotypeGDMCC 2.524 preserved as a metabolically inactive state, ex-type PYCC 10135).
Etymology.
The specific epithet haianensis refers to the geographic origin of the type strain, Wuzhi Mountain, Wuzhishan City, Hainan Province.
Description.
After 7 days of growth on YM agar at 25 °C, the colonies appear white to cream-colored, buttery, and smooth, with entire margins. After 3 days of growth in YM agar at 25 °C, cells are ovoid to ellipsoid (2.1–4.6 × 3.4–8.1 μm) and occur singly or in pairs. Budding is multilateral. Pseudohyphae are formed, but true hyphae are absent in slide culture on cornmeal agar after 7 days at 25 °C. No asci or signs of conjugation are observed on YM agar, 5% ME agar, Fowell’s acetate agar, CM agar, or Gorodkowa agar after 6 weeks at 15 or 25 °C. Fermentation is negative. Glucose, inulin (weak), galactose (weak), trehalose, cellobiose (weak), salicin (weak), d-xylose, l-arabinose, d-arabinose, d-ribose (delayed), ethanol, glycerol, erythritol, ribitol, d-mannitol, d-glucitol, succinate, citrate, d-gluconate, N-aetyl-d-glucosamine, and d-glucono-1, 5-lactone are assimilated. No growth is observed with sucrose, raffinose, melibiose, lactose, maltose, melezitose, methyl α-d-glucoside, l-sorbose, l-rhamnose, 5-keto-d-gluconate, methanol, galactitol, myo-inositol, dl-lactate, d-glucosamine, 2-keto-d-gluconate, or d-glucuronate. In nitrogen-assimilation tests, growth is present in ethylamine, l-lysine, and cadaverine, while growth is absent in nitrate, nitrite, and creatine. Growth is observed at 30 °C but absent at 35 °C. No growth is noted in the presence of 10% NaCl with 5% glucose, 0.01% cycloheximide, or 1% acetic acid. Starch-like compounds are not produced. Urease activity and diazonium blue B reactions are negative.
Morphology of Yamadazymahainanensis (ex-type, NYNU 24829). A. Colony on YM agar after 7 days at 25 °C; B. Budding cells on YM agar after 3 days at 25 °C; C. Pseudohyphae with blastoconidia on CM agar after 7 days at 25 °C. Scale bars: 10 μm.
Additional isolates examined.
China • Hainan Province, Wuzhishan City, Wuzhi Mountain, in the phylloplane of Pollia sp., August 2024, S.L. Lv, strain numbers: NYNU 24905, NYNU 249216.
GenBank accession numbers.
Holotype GDMCC 2.524^T^ (ITS: PQ568984; LSU D1/D2: PQ568981); additional isolates NYNU 24905 (ITS: MZ318442; LSU D1/D2: MZ318423) and NYNU 249216 (ITS: MZ318424; D1/D2 LSU: MZ318425).
Notes.
Physiologically, Y.hainanensis sp. nov. differs from its closely related known species, Y.tallmaniae, in several aspects (Table 2). Y.hainanensis sp. nov. is unable to ferment glucose, whereas Y.tallmaniae can. Additionally, Y.hainanensis sp. nov. is incapable of assimilating sucrose, maltose, melezitose, methyl-α-d-glucoside, or d-glucosamine, unlike Y.tallmaniae. Furthermore, Y.tallmaniae can grow in the presence of 10% NaCl with 5% glucose, while Y.hainanensis sp. nov. cannot.
Discussion
Initially, the identification of Yamadazyma was primarily based on phenotypic traits, which resulted in its polyphyletic nature. However, with the advent of molecular phylogenetic techniques, more comprehensive approaches have been employed to address taxonomic challenges within the genus. For example, Y.farinosa was reclassified as Millerozymafarinosa and Y.guilliermondii as Meyerozymaguilliermondii based on concatenated LSU D1/D2 and SSU gene sequences (Kurtzman and Suzuki 2010). Further phylogenetic analysis using combined ITS and LSU D1/D2 sequences recognized Yamadazyma as a well-supported monophyletic clade (Groenewald et al. 2011). In accordance with the Melbourne Code (McNeill et al. 2012), which permits the assignment of related anamorphic and teleomorphic species to the same genus, additional anamorphic Candida species were subsequently reclassified into the genus Yamadazyma, thus broadening its diversity to encompass both teleomorphic and anamorphic forms.
In this study, two novel species—Y.akebiae sp. nov. and Y.hainanensis sp. nov.—are described based on a combination of molecular phylogenetic analyses and phenotypic characteristics. Phylogenetically, Y.akebiae sp. nov. is closely related to Y.kanchanaburiensis, forming a moderately supported clade. In contrast, Y.hainanensis sp. nov. forms a distinct branch clearly separated from other Yamadazyma species; however, a more robust phylogenetic placement for this species awaits further validation. The sequence divergences of the LSU D1/D2 and ITS regions between the new species and their closest relatives are below the similarity thresholds typically used to define species boundaries in ascomycetous yeasts (Kurtzman and Robnett 1998; Vu et al. 2016). Phenotypically, the two species share traits such as colony morphology, color, and cell shape, but can be differentiated from their closest relatives by distinct physiological and biochemical characteristics (Table 2). The integration of genetic, phylogenetic, and morphological data strengthens the reliability of species delimitation.
Yeast classification has traditionally relied on their ability to ferment sugars, typically measured by CO_2_ production (Kurtzman et al. 2011). Most ascomycetous yeasts, with few exceptions, are capable of sugar fermentation, whereas basidiomycetous yeasts generally are not. Interestingly, the three Chinese strains identified as Y.hainanensis sp. nov. were negative for sugar fermentation (Table 2), a rare feature within Yamadazyma. Only two other species, Y.paraaseri and Y.andamanensis, have been reported to lack this ability (Am-In et al. 2011; Gao et al. 2021; Avesani et al. 2024).
To date, 68 species of the genus Yamadazyma have been formally described, including the two introduced in this study (Avesani et al. 2024; Seike et al. 2024). Of these, 12 species have been recorded in China (e.g., Y.diospyri, Y.paraphyllophila, Y.insectorum, Y.akitaensis, Y.olivae, Y.dushanensis, Y.luoyangensis, Y.ovata, Y.paraaseri, Y.triangularis, Y.akebiae, and Y.hainanensis) (Gao et al. 2021; Cai et al. 2022). As one of the most biodiverse countries globally, China is likely home to many more undescribed Yamadazyma species. Ongoing surveys and taxonomic efforts are essential for a more complete understanding of species richness within the genus.
Numerous Yamadazyma species have been studied extensively, and some strains are used in agriculture and biotechnology (González-Gutiérrez et al. 2024; Lopes et al. 2018; Cai et al. 2022). While these yeasts are known to inhabit diverse ecological niches, environmental changes—especially those driven by climate change—are affecting both terrestrial and aquatic habitats (Jain et al. 2020). Such shifts may influence the global distribution and diversity of Yamadazyma species. Future research should therefore not only continue to document novel species and their distribution but also assess how climate variability may impact this ecologically and industrially important genus.
Supplementary Material
XML Treatment for Yamadazyma akebiae
XML Treatment for Yamadazyma hainanensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Al-Sweih N Ahmad S Khan S Khan Z Joseph L Vayalil S Chandy R (2017) Persistent Candidaconglobata bloodstream infection in a preterm neonate successfully treated by combination therapy with amphotericin B and caspofungin.Journal de Mycologie Médicale 27(2): 71–276. 10.1016/j.mycmed.2017.01.01028189376 · doi ↗ · pubmed ↗
- 2Am-In S Limtong S Yongmanitchai W Jindamorakot S (2011) Candidaandamanensis sp. nov., Candidalaemsonensis sp. nov. and Candidaranongensis sp. nov., anamorphic yeast species isolated from estuarine waters in a Thai mangrove forest.International Journal of Systematic and Evolutionary Microbiology 61(2): 454–461. 10.1099/ijs.0.022038-020207807 · doi ↗ · pubmed ↗
- 3Andrew R (2016) Fig Tree: Tree figure drawing tool Version 1.4.3. Institute of Evolutionary Biology, University of Edinburgh Press, United Kingdom.
- 4Avesani M Zapparoli G Jindamorakot S Limtong S (2024) Yamadazymaoleae f.a. sp. nov. and Yamadazymamolendinolei f.a. sp. nov., two novel ascomycetous yeast species isolated from olive oil mills in Italy, and reassignment of 11 Candida species to the genus Yamadazyma. International Journal of Systematic and Evolutionary Microbiology 74(12): ее006592. 10.1099/ijsem.0.00659239621539 · doi ↗ · pubmed ↗
- 5Billon-Grand GA (1989) A new ascosporogenous yeast genus: Yamadazyma gen. nov.Mycotaxon 35: 201–204.
- 6Burgaud G Arzur D Sampaio JP Barbier G (2011) Candidaoceani sp. nov., a novel yeast isolated from a Mid-Atlantic Ridge hydrothermal vent (-2300 meters).Antonie van Leeuwenhoek 100(1): 75–82. 10.1007/s 10482-011-9566-121359849 · doi ↗ · pubmed ↗
- 7Burgaud G Coton M Jacques N Debaets S Maciel NOP Rosa CA Gadanho M Sampaio JP Casaregola S (2016) Yamadazymabarbieri f.a. sp. nov., an ascomycetous anamorphic yeast isolated from a Mid-Atlantic Ridge hydrothermal site (-2300 m) and marine coastal waters.International Journal of Systematic and Evolutionary Microbiology 66(9): 3600–3606. 10.1099/ijsem.0.00123927306608 · doi ↗ · pubmed ↗
- 8Cai J Xing L Zhang W Fu L Zhang J (2022) Selection of Potential Probiotic Yeasts from Dry-Cured Xuanwei Ham and Identification of Yeast-Derived Antioxidant Peptides. Antioxidants 11(10): е1970. 10.3390/antiox 11101970 PMC 959875836290693 · doi ↗ · pubmed ↗