New species and new records of Trechispora (Trechisporales, Basidiomycota) from Taiwan
Yi-Chung Lin, Yu-Ming Huang, Yi-Lun Huang, Shi-Liang Liu, Shuang-Hui He, Li-Wei Zhou, Che-Chih Chen

TL;DR
This study discovers and documents new species of the fungal genus Trechispora in Taiwan, expanding the known diversity and providing an identification key.
Contribution
The paper introduces four new species and seven new records of Trechispora in Taiwan through morphological and phylogenetic analyses.
Findings
Four new Trechispora species were described from Taiwan.
Seven species were newly recorded in Taiwan's fungal diversity.
The total number of Trechispora species in Taiwan was updated to 17.
Abstract
Trechispora (Hydnodontaceae) comprises a diverse group of wood- and soil-inhabiting fungi, primarily functioning as saprotrophs, with some species forming symbiotic associations with plants and animals. Despite the recognition of over 100 species worldwide, its diversity in Taiwan remains understudied. This study presents the first comprehensive taxonomic revision of Trechispora in Taiwan, integrating morphological and phylogenetic analyses based on sequence data from the nuc rDNA internal transcribed spacer ITS1-5.8S-ITS2 (ITS) region and the nuc 28S rDNA (28S). We describe four new species (Trechispora acerosa, T. floralis, T. formosana, and T. orchidophila) and report seven newly recorded species for Taiwan (T. crystallina, T. dentata, T. latehypha, T. mollusca, T. odontioidea, T. subsinensis, and T. wenshanensis). T. sinensis is synonymized under T. odontioidea. Morphological and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 10
Figure 10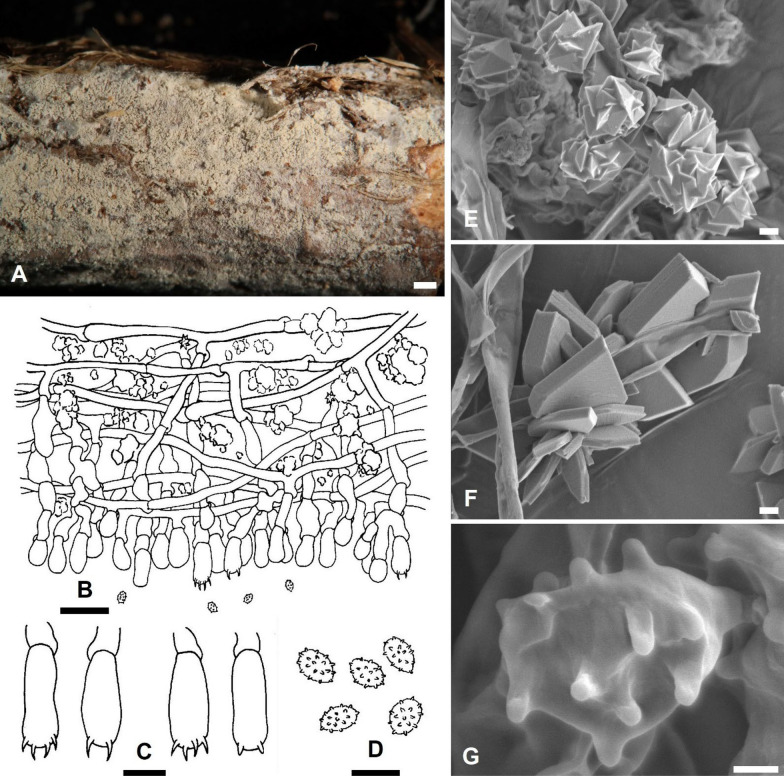 Figure 11
Figure 11 Figure 12
Figure 12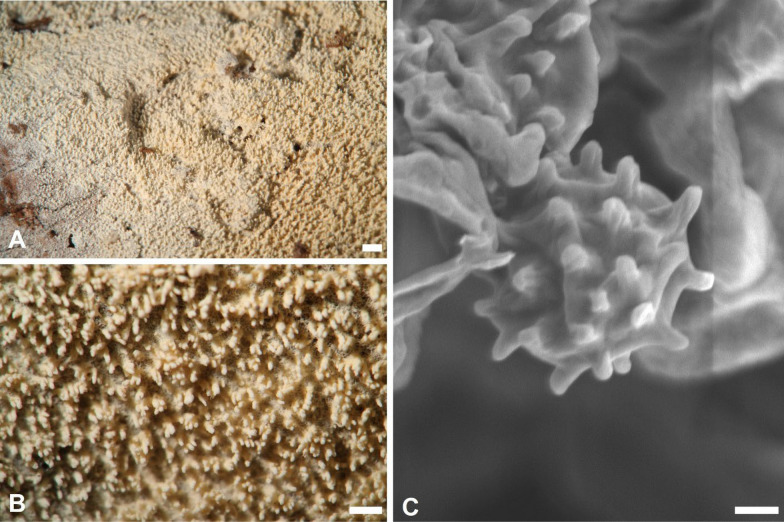 Figure 13
Figure 13 Figure 14
Figure 14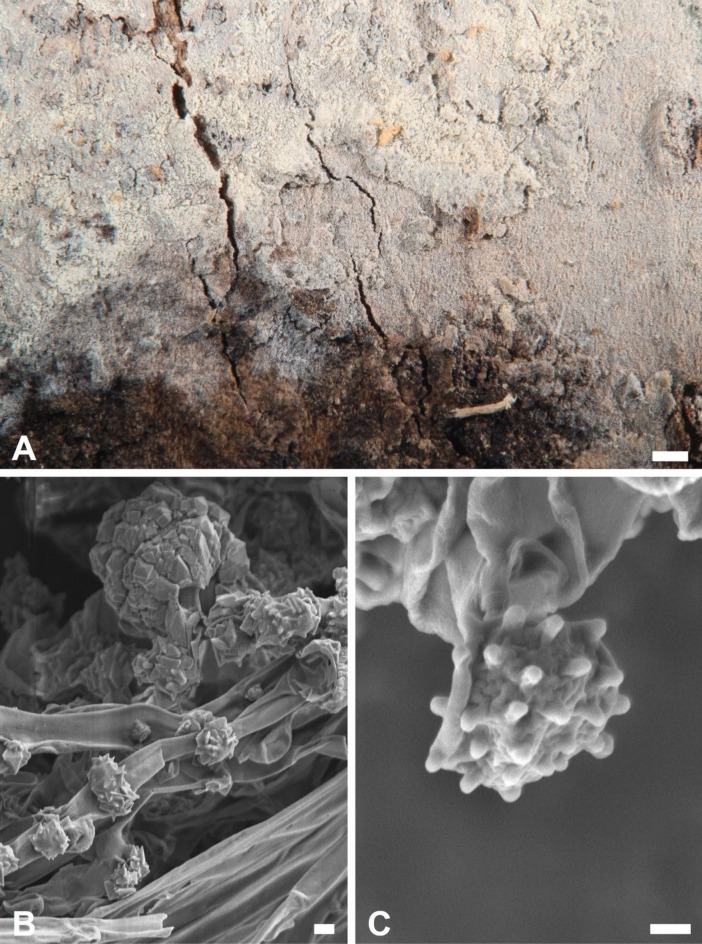 Figure 15
Figure 15 Figure 1
Figure 1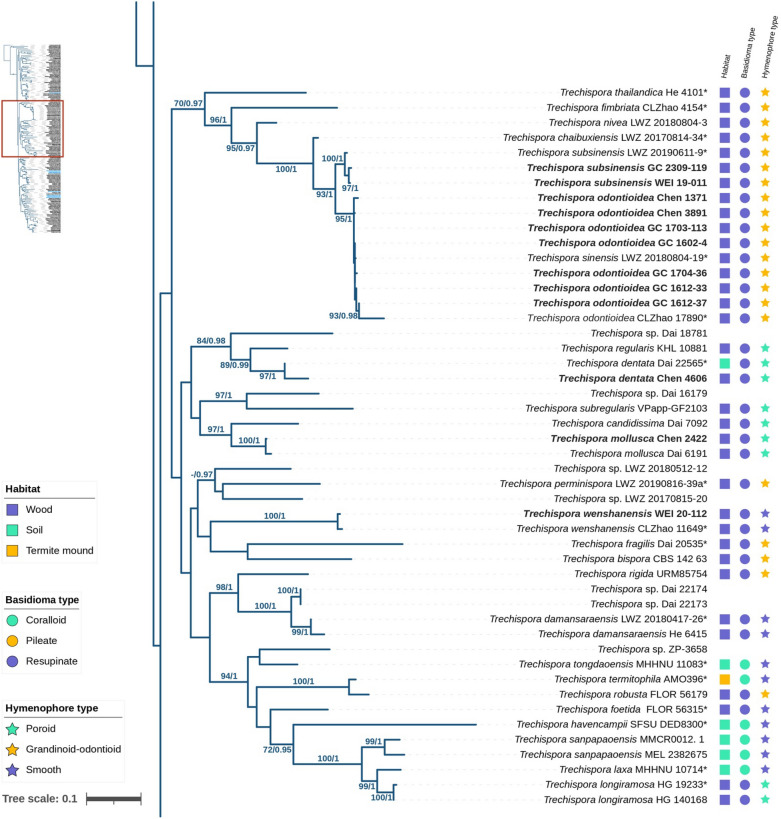 Figure 1
Figure 1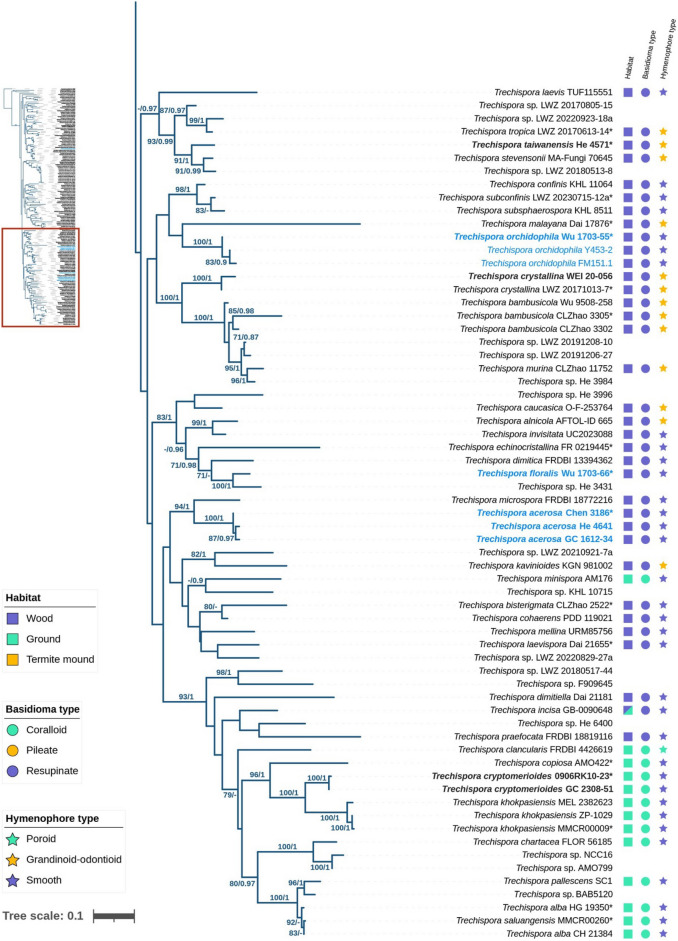 Figure 1
Figure 1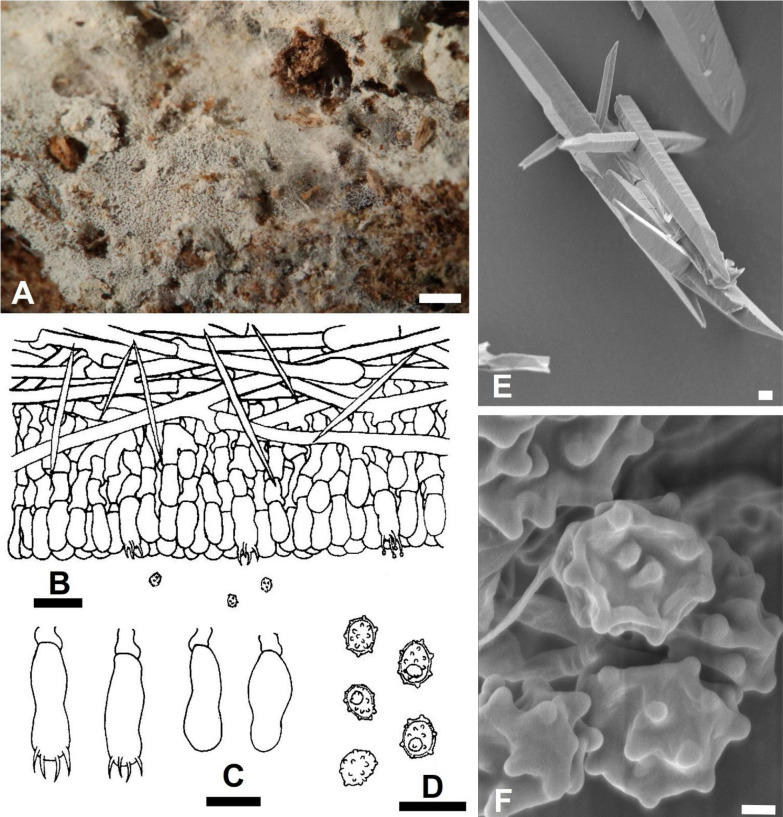 Figure 2
Figure 2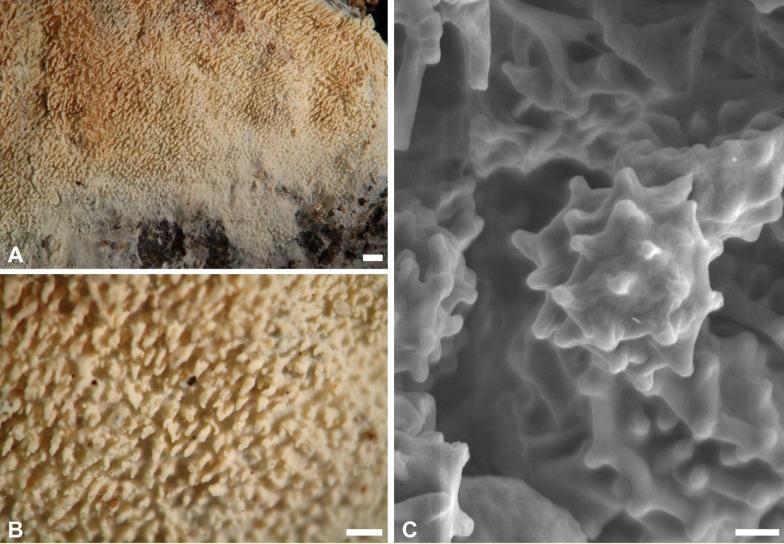 Figure 3
Figure 3 Figure 4
Figure 4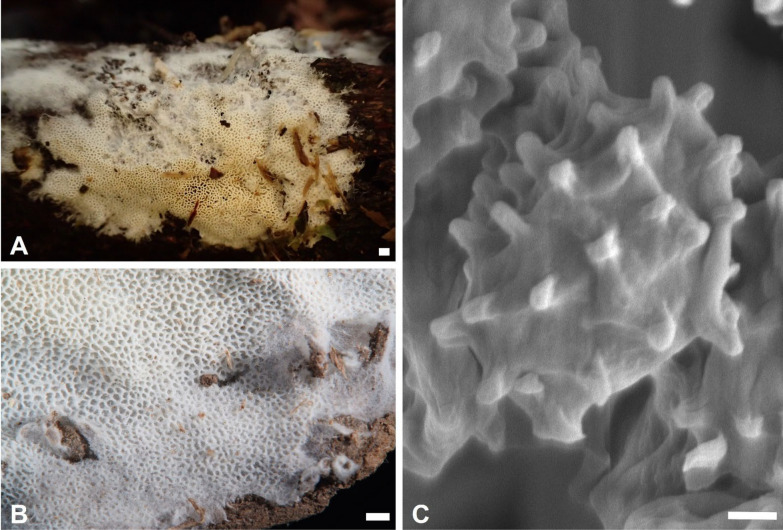 Figure 5
Figure 5 Figure 6
Figure 6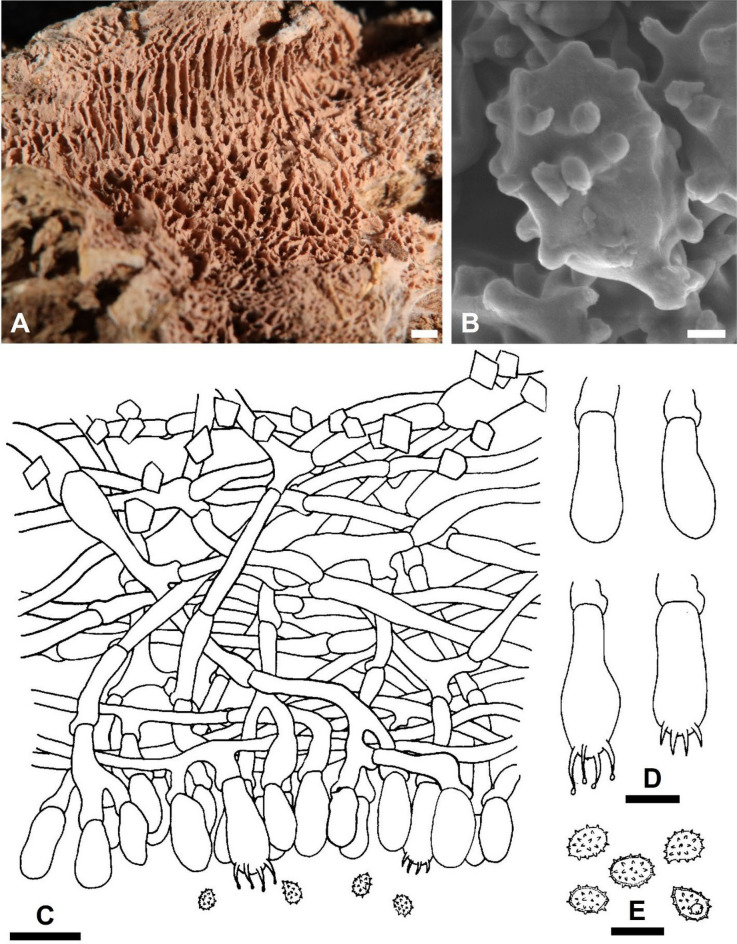 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9- —http://dx.doi.org/10.13039/100020595National Science and Technology Council
- —Forestry and Nature Conservation Agency of the Ministry of Agriculture (TW)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Lichen and fungal ecology · Plant Pathogens and Fungal Diseases
Background
Trechispora (Hydnodontaceae Jülich) is the largest genus in Trechisporales K.H. Larss (Hibbett et al. 2007), typified by T. onusta P. Karst. (Karsten 1890). It includes basidiomata of coralloid, pileate, and resupinate forms, with smooth, grandinioid, odontioid, hydnoid, or poroid hymenophores (Bernicchia and Gorjón 2010; de Meiras-Ottoni et al. 2021; Karsten 1890). Microscopically, Trechispora has monomitic or dimitic hyphal systems with nodose-septate generative hyphae, short basidia, and basidiospores ranging from smooth to verrucose or aculeate. Ampullate septa occur on subicular hyphae, especially in some mycelial cords (Larsson 1992). Calcium oxalate crystals on the subicular hyphae provide additional diagnostic features (Larsson 1992, 1994).
Most Trechispora species are saprotrophs found on decayed wood, bamboos, grasses, palms, tree ferns, mosses, or soil with litter (Chikowski et al. 2020; Larsson 1992; Lin et al. 2022; Liu et al. 2019). They play an important role as decomposers in forest ecosystems (de Meiras-Ottoni et al. 2021; Jülich 1976; Larsson 1992). Some species have been identified as potential ectomycorrhizal fungi (Dunham et al. 2007; Rosenthal et al. 2017; Vanegas-Leon et al. 2019), endophytes (Fulthorpe et al. 2020; Shen et al. 2022), or even pathogens (Wilkinson 1987). Additionally, Matsuura and Yashiro (2010) reported an unknown Trechispora species forming termite-mimicking sclerotia in Nasutitermes takasagoensis nests.
Currently, over 100 species of Trechispora are recognized worldwide (Chikowski et al. 2020; Drechsler-Santos et al. 2008; Hjortstam and Ryvarden 2007; Kirk et al. 2008; Larsson 1994, 1996; Liu et al. 2022a, 2022b; Luo and Zhao 2022; Ordynets et al. 2015; Phookamsak et al. 2019; Westphalen and Silveira 2012). Among these, only seven species have been recorded in Taiwan: T. cryptomerioides (W.-R. Lin & P.-H. Wang) S.L. Liu & L.W. Zhou, T. dimitica Hallenb., T. farinacea (Pers.) Liberta, T. lunata (Romell ex Bourdot & Galzin) Jülich, T. praefocata (Bourdot and Galzin) Liberta, T. rigida (Berk.) K.H. Larss., and T. taiwanensis S.L. Liu, S.H. He & L.W. Zhou (Lin et al. 2022; Liu et al. 2022a; Maekawa 1992; Wu 2003).
This study provides the first comprehensive taxonomic revision of Trechispora in Taiwan, integrating morphological and phylogenetic analyses. We describe and illustrate newly discovered species, clarify the taxonomic placement of previously recorded taxa, and present an identification key to accepted Trechispora species in Taiwan.
Materials and methods
Morphological studies
The studied specimens were deposited at the fungaria of Beijing Forestry University, Beijing (BJFC), and the National Museum of Natural Science, Taichung (TNM). The acronyms follow the Index Herbariorum (http://sweetgum.nybg.org). Basidiomata were photographed using an Olympus TG-4 or a Canon EOS R digital camera. The hymenial surface was examined under an Olympus SZ51 (Olympus, Tokyo, Japan) stereomicroscope at magnifications up to × 50. Free-hand thin sections of basidiomata were mounted in 5% potassium hydroxide (KOH) with 1% phloxine for observation and measurements and in Melzer’s reagent to test for amyloidity and dextrinoidity, using a ZEISS Axioscope 5 microscope (Carl Zeiss, Germany) at magnifications up to 1000 × . The following abbreviations are used: L = mean basidiospore length (arithmetical average for all basidiospores), W = mean basidiospore width (arithmetical average for all basidiospores), Q = L/W ratio, n = the number of basidiospores measured per specimen, m = meters above sea level. Microscopic measurements and morphological terminology followed the protocols established by Wu (1990).
For scanning electron microscopy (SEM) imaging, small fragments of dried basidiomata were directly mounted onto specimen stubs. The samples were then sputter-coated with platinum using a JEC-3000FC Auto Fine Coater (JEOL, Tokyo, Japan) and examined for basidiospore and crystal morphology using a JSM-7800F Schottky Field Emission Scanning Electron Microscope (JEOL).
DNA extraction, polymerase chain reaction (PCR), and DNA sequencing
DNA was extracted from either dried specimens or mycelia cultivated on 2% MEA. Tissue disruption and homogenization were performed before extraction with the aid of liquid nitrogen and Tissue Lyser II (Qiagen, Hilden, Germany). DNA was extracted using the NautiaZ Plant DNA Extraction Mini Kit (Nautia Gene, Taipei, Taiwan) following the manufacturer’s protocols. The study targeted two genetic regions: the nuc rDNA internal transcribed spacer ITS1-5.8S-ITS2 (ITS) region was amplified using the primer pair ITS1F/ITS4B (Gardes and Bruns 1993), while the nuc 28S rDNA (28S) was amplified using primer pair LR0R/LR5 (Vilgalys and Hester 1990). Amplifications were conducted in a Bio-Rad T100™ Thermal Cycler (Bio-Rad, Hercules, California, U.S.). The PCR conditions for the ITS and 28S regions were as follows: initial denaturation at 94 °C for 5 min, followed by 36 cycles at 94 °C for 30 s, 55 °C for 30 s, and 72 °C for 1 min, and a final extension at 72 °C for 7 min. PCR products were purified and sequenced using the Sanger method by Genomics BioSci & Tech (New Taipei, Taiwan). Newly generated sequences were manually assembled using BioEdit v. 7.2.5 (Hall 1999) or Geneious Prime 2024.0.5 (https://www.geneious.com), and their quality was checked based on five guidelines by Nilsson et al. (2012) before submission to GenBank (https://www.ncbi.nlm.nih.gov/genbank/). Consensus sequence accuracy and identity were verified by comparison with sequences in GenBank.
Alignment, phylogenetic analyses, and visualizing trees
Sequences of each single-gene dataset were aligned with MAFFT v. 7.409 (Katoh and Standley 2013), using the default algorithm and manually adjusted with MEGA v. 7 (Kumar et al. 2016) when necessary. Porpomyces mucidus (Pers.) Jülich was selected as the outgroup according to Liu et al. (2022a). The resulting alignments were deposited at Figshare (http://dx.doi.org/10.6084/m9.figshare.28407632). The Bayesian Inference (BI) and Maximum Likelihood (ML) methods were applied to the datasets using MrBayes v. 3.2.6 (Ronquist et al. 2012) and RAxML Black-Box (Stamatakis 2014), respectively. For the BI analyses, jModeltest 2.1.10 (Darriba et al. 2012) was first used to estimate separate models for each gene region in all datasets, based on Akaike information criterion (AIC). The Markov chain Monte Carlo (MCMC) search was run for ten million generations, with four chains and trees sampled every 1000 generations. The first 25% of trees (2500) were discarded as burn-in while the remaining trees were used to construct the 50% majority-rule consensus phylogram with posterior probabilities (PP). For the ML analysis, the best-scoring tree with proportional values of bootstrap (BS) was computed under a GTRGAMMA model with 1000 bootstrap replicates, followed by a thorough ML search. Partitioned BI and ML analyses were conducted (Table 1). Gaps were treated as missing data. Branches were regarded as having statistical support if values of PP and/or BS were ≥ 0.9 and ≥ 70%, respectively. Both BI and ML analyses were performed at the CIPRES Science Gateway (Miller et al. 2010; https://www.phylo.org/). The phylogenetic trees were visualized and edited in TreeGraph2 (Stöver and Müller 2010), Interactive Tree of Life (iTOL) v6 (Letunic and Bork 2024; https://itol.embl.de/), and Adobe Illustrator 27.9 (Adobe Systems, Inc). Table 1. Species and sequences used in the phylogenetic analysesSpeciesSample noLocationITS28SReferencesDextrinocystis calamicolaHe 5693ChinaMK204533MK204546Liu et al. (2019)Porpomyces mucidusDai 12692Czech RepublicKT157833KT157838Wu et al. (2015)Trechispora acerosaChen 3186Taiwan**PV085797PV085828****This studyT. acerosaGC 1612-34**Taiwan***PV085798PV085827**This studyT. acerosaHe 4641ChinaOM523513OM339319Liu et al. (2022a)T. albaCH21384ChinaOR557258–Liu et al. (2024)T. albaHG 19350ChinaOM523516–Liu et al. (2022a)T. albofarinosaCLZhao 4356ChinaOQ241383OQ282703Luo et al. (2024)T. alnicolaAFTOL-ID 665–DQ411529AY635768UnpublishedT. araneosaKHL 8570SwedenAF347084AF347084Larsson et al. (2004)T. bambusicolaCLZhao 3302ChinaMW544021MW520171Zhao and Zhao (2021)T. bambusicolaCLZhao 3305ChinaMW544022MW520172Zhao and Zhao (2021)T. bambusicolaWu 9508-258ChinaPV085818PV085820**This studyT. bisporaCBS:142.63AustraliaMH858241MH869842Vu et al. (2019)T. bisterigmataCLZhao 2522ChinaOQ241386–Luo et al. (2024)T. candidissimaDai 7092ChinaOM523407OM339229Liu et al. (2022a)T. caucasicaO-F-253764SwedenUDB038261–UnpublishedT. caulocystidiataFLOR 56314BrazilMK458772–Furtado et al. (2021)T. chaibuxiensisLWZ 20170814-34ChinaOM523409OM339231Liu et al. (2022a)T. chartaceaFLOR 56185BrazilMK458775–Liu et al. (2022a)T. clancularisFRDBI 4426619UKMW487976–UnpublishedT. cohaerensHHB 19445New ZealandMW740327–UnpublishedT. confinisKHL 11064SwedenAF347081AF347081Larsson et al. (2004)T. constrictaDai 10534ChinaOM523416–Liu et al. (2022a)T. constrictaHe 5899ChinaOM523417OM339236Liu et al. (2022a)T. copiosaAMO422*BrazilMN701013MN687971de Meiras-Ottoni et al. (2021)T. cryptomerioides0906RK10-23TaiwanKF679506OK422242Lin et al. (2022)T. cryptomerioidesGC 2308-51TaiwanPV085817–**This studyT. crystallinaLWZ 20171013-7*VietnamOM523420OM339239Liu et al. (2022a)T. crystallinaWEI 20-056TaiwanPV085799–**This studyT. cyatheaeFR 0219443FranchUDB024016UDB024017Ordynets et al. (2015)T. damansaraensisLWZ 20180417-26Malaysia–OM339241Liu et al. (2022a)T. damansaraensisHe 6415MalaysiaOM523421OM339240Liu et al. (2022a)T. dealbataFLOR 56183BrazilMK458777–Liu et al. (2022a)T. dentataChen 4606Taiwan–PV085826**This studyT. dentataDai 22565ChinaOK298491OM049408Liu et al. (2022b)T. dimiticaFRDBI 13394362UKMW487977OR892779UnpublishedT. dimitiellaDai 21181ChinaOK298493OK298949Liu et al. (2022b)T. echinocristallinaFR 0219445FranchUDB024018UDB024019Ordynets et al. (2015)T. echinosporaMA-Fungi 82485Equatorial GuineaJX392845JX392846Tellería et al. (2013)T. farinaceaKHL 8451–AF347082AF347082UnpublishedT. fimbriataCLZhao 4154ChinaMW544023MW520173Zhao and Zhao (2021)T. fissurataLWZ 20171015-35*VietnamOM523431OM339249Liu et al. (2022a)T. floralisWu 1703-66Taiwan****PV085813PV085822**This studyT. foetidaFLOR 56315*BrazilMK458769–Furtado et al. (2021)T. formosanaChen 3151Taiwan****PV085800PV085823**This studyT. fragilisDai 20535ChinaOK298494OK298950Liu et al. (2022b)T. gelatinosaAMO1139BrazilMN701021MN687978de Meiras-Ottoni et al. (2021)T. gracilisLWZ 20210626-5bChinaOM523436OM339254Liu et al. (2022a)T. havencampiiSFSU DED8300AfricaNR154418KT253947Desjardin and Perry (2015)T. hondurensisHONDURAS19-F016HondurasMT571523MT636540UnpublishedT. hymenocystisDai 2247FinlandOM523439–Liu et al. (2022a)T. hymenocystisKHL 8795SwedenAF347090AF347090Larsson et al. (2004)T. incisaGB-0090648SwedenKU747095KU747087UnpublishedT. invisitataUC2023088USAKP814425–UnpublishedT. kavinioidesKGN 981002NorwayAF347086AF347086Larsson et al. (2004)T. khokpasiensisMEL2382623AustraliaKP012986–Deng et al. (2023)T. khokpasiensisMMCR00009ThailandMZ687107MZ683197Deng et al. (2023)T. khokpasiensisZP-1029ChinaOM523532–Deng et al. (2023)T. laevisTUF115551EstoniaUDB016406–UnpublishedT. laevisporaDai 21655ChinaOK298495OM108710Liu et al. (2022b)T. larssoniiLWZ 20190817-11aChinaOM523442OM339259Liu et al. (2022a)T. latehyphaChen 3372TaiwanPV085801–This studyT. latehyphaGC 2109-57TaiwanPV085802–**This studyT. latehyphaHe 3924ChinaOM523443OM339261Liu et al. (2022a)T. latehyphaHe 5438ChinaOM523445–Liu et al. (2022a)T. laxaMHHNU10714ChinaOP959650OP954661Deng et al. (2023)T. longiramosaCH 19233ChinaOM523449–Liu et al. (2022a)T. longiramosaHG 140168ChinaOM523448OM339264Liu et al. (2022a)T. malayanaDai 17876SingaporeOM523452OM339265Liu et al. (2022a)T. mellinaURM85756Brazil–MH280000Chikowski et al. (2020)T. microsporaFRDBI 18772216UKOL828778–UnpublishedT. minisporaAM176MexicoMK328886MK328895Yuan et al. (2020)T. mollisURM85884*BrazilMK514945MH280003Chikowski et al. (2020)T. molluscaChen 2422TaiwanPV085803–**This studyT. molluscaDai 6191ChinaOM523455OM339269Liu et al. (2022a)T. murinaCLZhao 11752*ChinaOL615004OL615009Luo and Zhao (2022)T. niveaLWZ 20180804-3ChinaOM523461OM339273Liu et al. (2022a)T. orchidophilaFM151.1Réunion, FranchJF691276–Martos et al. (2012)T. orchidophilaWu 1703-55Taiwan****PV085812PV085824**This studyT. orchidophilaY453-2Okinawa, JapanLC327027–Ogura‐Tsujita et al. (2018)T. odontioideaChen 1371TaiwanPV085804****–****This studyT. odontioideaChen 3891TaiwanPV085805–**This studyT. odontioideaCLZhao 17890*ChinaON417458OQ282713Luo and Zhao (2022)T. odontioideaGC 1602-4TaiwanPV085806****PV085829****This studyT. odontioideaGC 1612-33TaiwanPV085807****–****This studyT. odontioideaGC 1612-37TaiwanPV085808****PV085830****This studyT. odontioideaGC 1703-113TaiwanPV085809****PV085819****This studyT. odontioideaGC 1704-36TaiwanPV085810–**This studyT. olivaceaCLZhao 17826ChinaON417457OQ282714Luo and Zhao (2022)T. pallescensSC1IndiaMZ518207MZ518091UnpublishedT. papillosaAMO795BrazilMN701023MN687981de Meiras-Ottoni et al. (2021)T. patawaensisVPapp-GF1901FrenchOL314550OL314546Crous et al. (2021)T. perminisporaLWZ20190816-39aChinaOM523525OM339329Liu et al. (2022a)T. praefocataFRDBI 18819116UKOL828784–UnpublishedT. regularisKHL 10881JamaicaAF347087AF347087Larsson et al. (2004)T. rigidaURM85754BrazilMT406381MH279999Chikowski et al. (2020)T. robustaFLOR 56179BrazilMK458770–Furtado et al. (2021)T. saluangensisMMCR00260ThailandMZ687104MZ683201Sommai S et al. 2023T. sanpapaoensisMEL:2382675AustraliaKP013038–UnpublishedT. sanpapaoensisMMCR00124.1ThailandMZ687109MZ683200Sommai et al. (2023)T. scabraFLOR 56189BrazilMK458773–Furtado et al. (2021)T. sinensisLWZ 20180804-19ChinaOM523482OM339290Liu et al. (2022a)Trechispora sp.AMO799BrazilMN701008MN687969de Meiras-Ottoni et al. (2021)Trechispora sp.BAB5120IndiaKT804576–UnpublishedTrechispora sp.Dai 16179ChinaOM523506OM339313Liu et al. (2022a)Trechispora sp.Dai 18781AustraliaOM523508OM339315Liu et al. (2022a)Trechispora sp.Dai 22173ChinaOK298496OK298951Liu et al. (2022b)Trechispora sp.Dai 22174ChinaOK298497OK298952Liu et al. (2022b)Trechispora sp.DLL2010-077USAJQ673209–UnpublishedTrechispora sp.DLL2011-186USAKJ140681–UnpublishedTrechispora sp.F909645SwedenJX392817JX392818Telleria et al. (2013)Trechispora sp.He 3431ChinaOM523509OM339316Liu et al. (2022a)Trechispora sp.He 3984ChinaOM523510OM339317Liu et al. (2022a)Trechispora sp.He 3996ChinaOM523511–Liu et al. (2022a)Trechispora sp.He 4503ChinaOM523512OM339318Liu et al. (2022a)Trechispora sp.He 5812Sri LankaOM523514OM339320Liu et al. (2022a)Trechispora sp.He 6400MalaysiaOM523515OM339321Liu et al. (2022a)Trechispora sp.KHL 10715–AF347088AF347088Larsson et al. (2004)Trechispora sp.KHL 16968BrazilMH290763–Chikowski et al. (2020)Trechispora sp.LWZ 20170805-15ChinaOM523517–Liu et al. (2022a)Trechispora sp.LWZ 20170815-20ChinaOM523518OM339322Liu et al. (2022a)Trechispora sp.LWZ 20171015-17VietnamOM523519OM339323Liu et al. (2022a)Trechispora sp.LWZ 20180512-12AustraliaOM523520OM339324Liu et al. (2022a)Trechispora sp.LWZ 20180513-8AustraliaOM523521OM339325Liu et al. (2022a)Trechispora sp.LWZ 20180517-43AustraliaOM523522OM339326Liu et al. (2022a)Trechispora sp.LWZ 20180517-44AustraliaOM523523OM339327Liu et al. (2022a)Trechispora sp.LWZ 20180517-45AustraliaOM523524OM339328Liu et al. (2022a)Trechispora sp.LWZ 20191206-27MalaysiaOM523526OM339330Liu et al. (2022a)Trechispora sp.LWZ 20191208-10MalaysiaOM523527–Liu et al. (2022a)Trechispora sp.LWZ 20210921-7aChinaOM523530OM339333Liu et al. (2022a)Trechispora sp.LWZ 20220829-27aChina****PV085831****PV082941****This studyTrechispora***** sp.LWZ 20220922-4aChina****PV085833PV082940This studyTrechispora sp.LWZ 20220923-18aChinaPV085832PV082939This studyTrechispora sp.NCC16BrazilMN701007MN687968de Meiras-Ottoni et al. (2021)Trechispora sp.SP48BrazilMN701005MN687965de Meiras-Ottoni et al. (2021)Trechispora sp.WEI 16-317Taiwan****PV085811PV085821**This studyTrechispora sp.Yuan 6129ChinaOM523531–Liu et al. (2022a)Trechispora sp.ZP-3658ChinaOM523533–Liu et al. (2022a)T. stevensoniiMA-Fungi 70645–JX392843JX392844Telleria et al. (2013)T. subaraneosaLWZ 20210918-10aChinaOM523529OM339332Liu et al. (2024)T. subconfinisLWZ 20230715-12aChinaPP959670–Wang et al. (2024)T. subfarinaceaLWZ 20200921-33aChinaOM523528OM339331Phookamsak et al. (2024)T. subfissurataHe 3907ChinaOM523490OM339298Liu et al. (2022a)T. subhymenocystisLWZ 20190818-29bChinaOM523492OM339299Liu et al. (2022a)T. subhymenocystisLWZ 20190818-32bChina–OM339300Liu et al. (2022a)T. subregularisVPapp-GF2103FrenchOL331097OL314548UnpublishedT. subsinensis*****GC 2309-119Taiwan**PV085815–**This studyT. subsinensis*LWZ 20190611-9ChinaOM523497OM339304Liu et al. (2022a)T. subsinensisWEI 19-011TaiwanPV085814–**This studyT. subsphaerosporaKHL 8511SwedenAF347080AF347080Larsson et al. (2004)T. taiwanensisHe 4571TaiwanOM523498OM339305Liu et al. (2022a)T. termitophilaAMO396BrazilMN701025MN687983de Meiras-Ottoni et al. (2021)T. thailandicaHe 4101 ThailandOM523499OM339307Liu et al. (2022a)T. thelephoraURM85758Brazil–MH280002Chikowski et al. (2020)T. tongdaoensisMHHNU11083ChinaOP959651OP954662Deng et al. (2023)T. torrendiiURM85886BrazilMK515148MH280004Chikowski et al. (2020)T. tropicaLWZ 20170613-14ChinaOM523502OM339310Liu et al. (2022a)T. tuberculataDai 17433BrazilOM523507OM339314Liu et al. (2022a)T. wenshanensisCLZhao 11649*ChinaOQ241389OQ282716Luo et al. (2024)T. wenshanensisWEI 20-112TaiwanPV085816**PV085825**This studyTubulicium raphidisporumHe 3191ChinaOM523534OM339334Liu et al. (2022a)Asterisks () indicate holotype sequences. Newly generated sequences are shown in bold
Results
Molecular phylogeny
The ITS + 28S dataset, comprising 162 fungal specimens, was analyzed using Bayesian Inference (BI) and Maximum Likelihood (ML) methods. The alignment contained a total of 2755 sites, including gaps, with 1247 sites for ITS and 1508 for 28S. The General Time Reversible model with gamma rate heterogeneity and invariant sites (GTR + G + I) was identified as the best-fit model for BI analyses based on the Akaike Information Criterion (AIC). The BI and ML analyses yielded similar topologies, supporting the placement of four newly described species (Fig. 1): Trechispora acerosa, T. floralis, T. formosana, and T. orchidophila in distinct lineages within the Trechispora clade (PP = 1, BS = 100%). T. acerosa and T. orchidophila formed independent lineages (both PP = 1, BS = 100%) unrelated to other species, while T. floralis and T. formosana were closely related to Trechispora sp. He 3431 and T. subhymenocystis, respectively (PP = 1, BS = 100%; PP = 1, BS = 94%).Fig. 1. The phylogram of Trechispora inferred from ML analyses using ITS + 28S dataset. Branches are labelled with BS ≥ 70% from ML and PP ≥ 0.9 from Bayesian analyses. Blue text indicates new species described in this study, while bold text represents species collected from Taiwan. Asterisks (*) indicate holotype specimens
Taxonomy
Trechispora acerosa Yi C. Lin and C. Chih Chen, sp. nov. Figure 2.Fig. 2Trechispora acerosa (from holotype). A Basidioma. B Part of vertical section through basidioma. C Basidia and basidioles. D Basidiospores. E–F Scanning electron micrographs of needle-like crystals (E) and basidiospores (F). Scale bars: A = 1 mm; B = 10 μm; C–D = 5 μm; E = 1 μm; F = 0.5 μm
MycoBank MB857631.
Etymology. acerosa (Lat.), referring to the needle-like crystals.
Diagnosis. Trechispora acerosa is distinguished by its white basidiomata with smooth hymenophore, verrucose, thick-walled basidiospores, and needle-like crystals.
Typification. TAIWAN. Nantou County, Jenai township, trailhead of Southern Tungyenshan, 24°2'N, 121°6'E, 1550 m, on rotten angiosperm trunk, 28 Mar 2016, leg. S.-Z. Chen, C.-C. Chen & C.-L. Wei, Chen 3186 (holotype TNM F0030009). GenBank: ITS = PV085797; 28S = PV085828.
Description. Basidiomata annual, resupinate, thin, soft, fragile, easily separated from substratum, up to 2 cm long, 1 cm wide. Hymenophore smooth, arachnoid to farinose, white to cream. Margin white to cream, fimbriate. Hyphal system monomitic; generative hyphae with clamp connections. Subicular hyphae long-celled, colorless, thin-walled, moderately branched and septate, subparallel to interwoven, 2–5.5 μm in diam, ampullate septa usually present in the hyphae, up to 6 μm wide. Generative hyphae distinct, colorless, thin or slightly thick-walled, short-celled moderately branched, smooth, subparallel, 2.5–4 μm in diam. Needle-like crystals frequently present in subiculum. Cystidia absent. Basidia cylindrical with a slight median constriction, colorless, thin-walled, with four sterigmata and a basal clamp connection, 11–14 × 4–4.6 μm. Basidioles similar in shape to basidia, but smaller. Basidiospores ovoid to ellipsoid, colorless, slightly thick-walled, becoming thick-walled when aged, verrucose, inamyloid, indextrinoid, usually with one oil drop, (2.9–)3.3–3.9(–4.3) × (2.2–)2.4–2.9(–3.2) μm, L = 3.6 μm, W = 2.7 μm, Q = 1.2–1.5 (n = 30).
Additional specimens examined. TAIWAN. Nantou County, Luku township, Hsitou, 23°41'N, 120°48'E, 1250 m, on fallen gymnosperm trunk, 11 Dec 2016, leg. S.-Z. Chen & C.-C. Chen, GC 1612-34 (TNM F0031438); ibid., on rotten Cunninghamia lanceolata, 11 Dec 2016, leg. S.-H. He, He 4641 (BJFC024084). Kaohsiung City, Taoyuan District, Shihshan Forestry Road, 23°04'N, 120°46'E, 1620 m, on rotten angiosperm trunk, 30 Jun 2017, leg. C.-C. Chen, C.-L. Wei, W.-C. Chen and Y.-P. Chen, WEI 17-439 (TNM F0032508).
Ecology and distribution. On rotten angiosperm or gymnosperm (e.g., Cunninghamia) trunks in Taiwan. Mar, Jun, Dec.
Notes. Trechispora acerosa resembles T. microspora (P. Karst.) Liberta and T. praefocata by having smooth hymenophore, needle-like crystals, and small basidiospores. However, T. acerosa differs from T. microspora by having thicker and narrower basidiospores (T. acerosa: 2.4–2.9 μm; T. microspora: 3–3.5 µm) (Bernicchia and Gorjón 2010). A key diagnostic feature of T. microspora is the absence of warts near the apiculus of basidiospores (Larsson 1992), whereas in T. acerosa, the warts are uniformly distributed (Figs. 2D, F). Compared with T. praefocata, T. acerosa has narrower basidia (T. acerosa: 4–4.6 μm; T. praefocata: 5–6 µm) and smaller basidiospores (T. praefocata: 5.5–6.5 × 4.5–5 µm) (Liberta 1966).
Trechispora crystallina S.L. Liu and L.W. Zhou, Mycosphere 13:906. 2022. Figures 3.Fig. 3Trechispora crystallina (from WEI 20–056). A–B Basidiomata. C Scanning electron micrograph of basidiospores. Scale bars: A = 1 mm; B = 0.5 mm;** C** = 1 μm
Description and illustration: See Liu et al. (2022a).
Specimens examined. CHINA. Jilin Province, Jilin City, Zuojia Town, 44°4'N, 126°6'E, 270 m, on angiosperm branch, 11 Aug 2016, leg. S.-H. Wu, Wu 1608-215 (TNM F0030631). TAIWAN. Taoyuan City, West Peak of Bajiawanshan, 24°38′08"N, 121°23′25"E, 1690 m, on rotten angiosperm branch, 13 Jul 2020, leg. C.-L. Wei, WEI 20-056 (TNM F0036784).
Ecology and distribution. On living or dead angiosperm wood in China (Inner Mongolia, Jilin), Vietnam, and Taiwan (Liu et al. 2022a; this study). Jul in Taiwan.
Notes. Trechispora crystallina is characterized by white to cream basidiomata with grandinioid hymenophore, presence of crystals in the subiculum and trama, and verrucose basidiospores (3.5–4.2 × 3–3.6 μm) (Liu et al. 2022a). The examined specimens share these features in most aspects, but WEI 20-056 has odontioid hymenophore (Figs. 3A–B), and Wu 1608–215 has basidiospores ranging from verrucose to aculeate. Additional specimens confirm variations in hymenophore configuration (grandinioid to odontioid) and basidiospore ornamentation (verrucose to aculeate) in this species. Trechispora crystallina is newly recorded from Taiwan.
Trechispora cryptomerioides (W.R. Lin & P.H. Wang) S.L. Liu & L.W. Zhou in Wang et al., Mycology 16:116. 2025. Figure 4.Fig. 4. Basidiomata of Trechispora cryptomerioides (from GC 2308–51) in situ. Scale bars: A = 1 cm; B = 5 mm. Photographed by Ms. Li-Hung Chen
Description and illustration. See Lin et al. (2022, as Scytinopogon cryptomerioides W.R. Lin & P.H. Wang).
Specimens examined. TAIWAN. Hsinchu County, Jianshi township, Lidong Villa to Mount Lidong Fort, 24°41′29"N, 121°18′23"E, 1500–1900 m, on the ground, 9 Sep 2021, leg. C.-C. Chen, GC 2109-44 (TNM F0038416); ibid., Mount Lidong Fort, 1900 m, on the ground, 22 Aug 2023, leg. L.-H. Chen, GC 2308–51 (TNM F0038419); ibid., Syakaro Historic Trail, 24°35′18"N, 121°15′10"E, 1300 m, on the ground, 9 Aug 2023, leg. C.-C. Chen & C.-C. Chang, GC 2308-12 (TNM F0038418). Nantou County, Hsinyi township, Jenlun Forest Road, on soil, 5 Jul 2009, leg. W.-R. Lin, 0906RK10-23 (TNM F0028829, holotype).
Ecology and distribution. On the ground in forests dominated by Cryptomeria japonica at elevations of 600–2100 m in Taiwan. Jun to Oct. The mycelium is associated with the roots of C. japonica (Lin et al. 2022; this study).
Notes. Trechispora cryptomerioides was originally placed in Scytinopogon and was recently transferred to Trechispora by Wang et al. (2024) based on morphological and phylogenetic evidence. This species is characterized by white to cream, coralloid basidiomata with smooth hymenophore, and aculeate basidiospores measured as 4–6 × 3–3.5 μm by Lin et al. (2022). Morphology of our specimens agrees with the protologue.
Trechispora dentata Z.B. Liu & Yuan Yuan, Frontiers in Microbiology 13:9, 2022. Figure 5.Fig. 5Trechispora dentata (from Chen 4606 except for A, which is from GC 2405–27). A–B Basidiomata when fresh (A) and dry (B).** C** Scanning electron micrograph of basidiospores. Scale bars: A–B = 1 mm; C = 0.5 μm
Description and illustration. See Liu et al. (2022b).
Specimens examined. TAIWAN. Nautou County, Jenai township, Meifeng, 24°05′29''N, 121°10′34''E, 2000–2100 m, on rotten wood, 5 May 2024, leg. C.-C. Chen, GC 2405-27 (TNM F0038421). Taichung City, Heping District, near Mt. Baimao trailhead, 24°10′10"N, 120°55′10"E, 960 m, on rotten angiosperm wood, 12 May 2021, leg. S.-Z. Chen & C.-L. Wei, Chen 4606 (TNM F0038415).
Ecology and distribution. On soil or rotten angiosperm wood in SW China (Yunnan) and Taiwan (Liu et al. 2022b; this study). May in Taiwan.
Notes. Trechispora dentata was recently described from Yunnan, China, based on a specimen collected from soil. It is characterized by white to cream basidiomata with distinctly irpicoid to dentate hymenophore (pores or aculei 3–4 per mm), and thick-walled, aculeate basidiospores measured as 4.1–5 × 3.2–4 μm by Liu et al. (2022b). Taiwanese specimens found on rotten wood agree with the protologue, except for having poroid hymenophore with more entire dissepiments (Figs. 5A–B). T. dentata is newly recorded from Taiwan.
Trechispora floralis Yi C. Lin & C. Chih Chen, sp. nov. Figure 6.Fig. 6Trechispora floralis (from holotype). A Basidioma. B Part of vertical section through basidioma. C Basidia. D Basidiospores. E–F Scanning electron micrographs of flower-like crystals (E) and basidiospores (F). Scale bars: A = 1 mm; B = 10 μm; C–D = 5 μm; E = 1 μm; F = 0.5 μm
MycoBank MB857633.
Etymology. floralis (Lat.), referring to the flower-like crystals.
Diagnosis. Trechispora floralis is distinguished by its white basidiomata with smooth hymenophore, aculeate basidiospores, and flower-like crystals.
Typification***.***** TAIWAN**. Nantou County, Jenai township, Huisun Forestry Station, 24°5'N, 121°2'E, 750 m, on rotten wood, 24 Mar 2017, leg. S.-H. Wu, Wu 1703-66 (holotype TNM F0031286). GenBank: ITS = PV085813; 28S = PV085822.
Description. Basidiomata annual, resupinate, thin, soft, fragile, easily separated from substratum, up to 1.5 cm long, 1 cm wide. Hymenophore smooth to grandinioid, farinaceous, white. Margin white, fimbriate. Hyphal system monomitic; generative hyphae with clamp connections. Subicular hyphae long-celled, colorless, thin-walled, moderately branched and septate, subparallel to interwoven, 1.5–2.5 μm in diam, ampullate septa usually present in the hyphae, up to 4.5 μm wide. Subhymenial hyphae distinct, colorless, tortuous, thin-walled, much branched, smooth, 3.5–7.5 μm in diam. Crystals frequently occurring in subiculum as aggregated, flower-like forms. Cystidia absent. Basidia subclavate to cylindrical, colorless, thin-walled, with four sterigmata and a basal clamp connection, 11–15 × 4.7–5.4 μm. Basidioles similar in shape to basidia, but smaller. Basidiospores ellipsoid, colorless, thin-walled, aculeate, inamyloid, indextrinoid, (3.9–)4.1–4.8(–5.4) × (2.7–)3.1–3.8(–4.2) μm, L = 4.5 μm, W = 3.6 μm, Q = 1.2–1.6 (n = 30).
Additional specimen examined. TAIWAN. Nantou County, Jenai township, Aowanda National Forest Recreation Area, 23°57'N, 121°11'E, 1250 m, 28 Aug 2017, on angiosperm branch, leg. C.-L. Wei & Y.-L. Huang, WEI 17-633 (TNM F0032638).
Ecology and distribution. On angiosperm wood in Taiwan. Mar, Aug.
Notes. Trechispora floralis resembles T. minima K.H. Larss. and T. damansaraensis S.L. Liu & L.W. Zhou having white basidiomata with smooth hymenophore, and aculeate basidiospores. However, T. floralis differs by having flower-like crystals and longer basidiospores than T. minima and T. damansaraensis, respectively (T. minima: 3.5–4 × 3.2–3.7 μm; T. damansaraensis: 3–3.8 × 2.3–3 µm) (Larsson 1992; Liu et al. 2022a).
Trechispora formosana Yi C. Lin & C. Chih Chen, sp. nov. Figure 7.Fig. 7Trechispora formosana (from holotype except for A, which is from Chen 3228). A Basidioma. B Scanning electron micrograph of basidiospores.** C** Part of vertical section through basidioma. D Basidia and basidioles. E Basidiospores. Scale bars: A = 1 mm; B = 0.5 μm; C = 10 μm; D–E = 5 μm
MycoBank MB857634.
Etymology. formosana (Lat.), referring to the Formosa, the historical name of Taiwan, the type locality.
Diagnosis. Trechispora formosana is distinguished by its brick-red basidiomata with poroid hymenophore, and aculeate basidiospores.
Typification. TAIWAN. Nantou County, Jenai township, trailhead of Southern Tungyenshan, 24°2'N, 121°6'E, 1550 m, on angiosperm wood, 28 Mar 2016, leg. S.-Z. Chen, C.-C. Chen & C.-L. Wei, Chen 3151 (holotype TNM F0029980). GenBank: ITS = PV085800; 28S = PV085823.
Description. Basidiomata annual, resupinate, soft, fragile, easily separated from substrates, up to 3 cm long, 1.5 cm wide. Hymenophore poroid, brick red, with round, angular to irregular pores, 2–3 per mm; dissepiments thin, entire. Subiculum white, soft corky, thin, about 0.1 mm thick. Tubes clay-pink, soft, up to 1 mm long. Margin white, fimbriate, rhizomorphic. Hyphal system monomitic; generative hyphae with clamp connections. Subicular hyphae long-celled, colorless, thin-walled, moderately branched and septate, subparallel to interwoven, 2–5.5 μm in diam, ampullate septa usually present in the hyphae, up to 8.5 μm wide. Generative hyphae distinct, colorless, thin or slightly thick-walled, moderately branched, smooth, more or less parallel, 3–5 μm in diam. Crystals often present as single rhomboidal plates in subiculum. Cystidia absent. Basidia cylindrical with a slight median constriction, colorless, thin-walled, with four sterigmata and a basal clamp connection, 13–22 × 5–6 μm. Basidioles similar in shape to basidia, but smaller. Basidiospores ovoid to ellipsoid, colorless, slightly thick-walled, thick-walled when aged, aculeate, inamyloid, indextrinoid, occasional with one oil drop, (3.8–)4–4.5(–5) × (3.1–)3.2–3.7(–4.4) μm, L = 4.2 μm, W = 3.5 μm, Q = 1.1–1.4 (n = 30).
Additional specimen examined. TAIWAN. Nantou County, Jenai township, trailhead of Southern Tungyenshan, 24°2'N, 121°6'E, 1550 m, on rotten wood, 28 Mar 2016, leg. S.-Z. Chen, C.-C. Chen & C.-L. Wei, Chen 3228 (TNM F0030028).
Ecology and distribution. On angiosperm wood in Taiwan. Mar.
Notes. Trechispora formosana is similar to T. hondurensis Schoutteten & Haelew., T. hymenocystis (Berk. & Broome) K.H. Larss., and T. subhymenocystis S.-L. Liu, H.-S. Yuan & L.-W. Zhou in having poroid hymenophore and aculeate basidiospores. However, T. hondurensis differs by having smaller basidiospores (3.7–3.8 × 2.8–2.9 μm) (Haelewaters et al. 2020), while T. hymenocystis has sphaerocysts in cords and larger basidiospores (4.5–5.5 × 3.5–4.5 μm) (Larsson 1994). Additionally, T. subhymenocystis differs by having white and slightly dentate pore surface, and growing on gymnosperms (Liu et al. 2022a).
Trechispora latehypha S.L. Liu, S.H. He & L.W. Zhou, Mycosphere 13:906. 2022. Figure 8.Fig. 8. Basidioma of Trechispora latehypha (from Chen 3372). Scale bar = 1 mm
Description and illustration. See Liu et al. (2022a).
Specimens examined. TAIWAN. Nautou County, Jenai township, trailhead of Southern Tungyenshan, 24°02'N, 121°06'E, 1568 m, on rotten bamboo culm, 7 Dec 2016, leg. S.-Z. Chen & C.-C. Chen, Chen 3372 (TNM F0030862). Yilan County, Yuanshan township, Caopi wetland, 24°46′03''N, 121°36′28''E, 800 m, on angiosperm branch, 28 Sep 2021, leg. C.-C. Chen, GC 2109-57 (TNM F0038417).
Ecology and distribution. On angiosperm and gymnosperm wood or bamboo culms in China (Fujian, Guangdong, Hainan) and Taiwan. Jun, Aug, Sep, Dec (Liu et al. 2022a; this study).
Notes. Trechispora latehypha is characterized by white to cream basidiomata with smooth hymenophore, rows of short-celled, wide hyphae in the subhymenium, thick-walled generative hyphae in the subiculum, and aculeate basidiospores measuring 3–3.5 × 2.4–2.9 μm (Liu et al. 2022a). It resembles T. wenshanensis and T. orchidophila in having a smooth hymenophore and aculeate basidiospores and may be confused with T. bambusicola due to its occurrence on bamboo. However, T. latehypha differs from T. wenshanensis and T. orchidophila by its thick-walled generative hyphae and lack of crystals, while T. bambusicola has an odontioid hymenophore. The morphology of our specimens agrees with the protologue. Trechispora latehypha is newly recorded from Taiwan.
Trechispora mollusca (Pers.) Liberta, Canadian Journal of Botany 51:1878. 1973. Figure 9.Fig. 9. Basidioma of Trechispora mollusca (from Chen 2422). Scale bar = 1 mm
Descriptions and illustrations. See Larsson (1992) and Bernicchia and Gorjón (2010).
Specimens examined. CHINA. Jilin Province, Antu County, Changbai Mountain Forest Ecosystem Research Station, 1300 m, 15 Sep 1995, on rotten wood of Abies, leg. Y.-C. Dai, Dai 2154 (TNM F0007320); ibid., 16 Aug 1997, on rotten wood of Abies, leg. Y.-C. Dai, Dai 2565 (TNM F0008167). Liaoning Province, Kuandian County, Baishilazi National Nature Reserve, 1 Sep 2004, on fallen trunk of Cerasus, leg. B.-K. Cui, Cui 1089 (TNM F0017524). FINLAND. Kittilän Lappi. Kittila, W of Akäskero, 31 Aug 1999, on fallen trunk of Betula, leg. Y.-C. Dai, Dai 3197 (TNM F0014047). TAIWAN. Taichung City, Hoping District, Anma Villa, 24°15'N, 121°0'E, 2150 m, 29 Jul 2014, on rotten angiosperm wood, leg. S.-Z. Chen & W.-C. Chen, Chen 2422 (TNM F0028196); ibid., Dasyueshan Forestry Road, 1900 m, 19 Aug 2008, on rotten angiosperm branch, leg. S.-H. Wu, S.-Z. Chen & Y.-T. Wang, Wu 0808-79 (TNM F0022702).
Ecology and distribution. On rotten gymnosperm or angiosperm wood in the forest regions of Northern Hemisphere (Larsson 1992; Bernicchia and Gorjón 2010). Jul, Aug in Taiwan.
Notes. Trechispora mollusca is characterized by white to cream basidiomata with poroid hymenophore (pores 2.5–5 per mm), thick-walled subicular hyphae, and aculeate, subglobose basidiospores measuring 3.5–5 × 2.5–3 μm by Liberta (1973). It resembles T. dentata and T. formosana in having resupinate basidiomata with poroid hymenophore; however, T. formosana has brick-red basidiomata, while T. dentata has thin-walled subicular hyphae (Liu et al. 2022b; this study). Trechispora mollusca is newly recorded from Taiwan.
Trechispora odontioidea K.Y. Luo & C.L. Zhao, Journal of Fungi 8:7. 2022. Figure 10.Fig. 10Trechispora odontioidea (from WEI 19–458). A–B Basidiomata. C Scanning electron micrograph of basidiospores. Scale bars: A–B = 1 mm; C = 0.5 μm
= Trechispora sinensis S.L. Liu, L.W. Zhou & S.H. He, Mycosphere 13:924. 2022. (syn. nov.)
Descriptions and illustrations. Liu et al. (2022a, as T. sinensis) and Luo and Zhao (2022).
Specimens examined. CHINA. Jilin Province, Antu County, Erdaobaihe Town, Changbaishan, 42°23'N, 128°6'E, 730 m, on rotten stump, 7 Sep 2011, leg. S.-H. Wu, Wu 1109-59 (TNM F0025606). Liaoning Province, Dalian City, Jinzhou District, Daheishan, 39°5'N, 121°47'E, 200 m, on rotten angiosperm trunk, 2 Aug 2017, leg. S.-H. Wu, Wu 1708-78 (TNM F0031640). TAIWAN. Kaohsiung City, Maolin District, Tona Nursery, 22°54'N, 120°44'E, 850 m, on angiosperm branch, 31 May 2005, leg. S.-Z. Chen, Chen 1371 (TNM F0018762). Miaoli County, Taian township, Shei-Pa National Park, East line of Talu Forest Road, 24°30'N, 121°7'E, 2040 m, on rotten trunk, 22 Apr 2017, leg. C.-C. Chen, GC 1704-36 (TNM F0031482). Nantou County, Hsinyi township, Tungpu, Tsaihung Waterfalls, 23°33'N, 120°56'E, 1400 m, on angiosperm branch, 1 Feb 2016, leg. C.-C. Chen & C.-L. Wei, GC 1602-4 (TNM F0030309); ibid., Yushan National Park, Japanese Occupation Era Batongguan Historical Trail, 23°33'N, 120°56'E, 1390 m, on fallen angiosperm trunk, 18 Mar 2018, leg. C.-C. Chen, GC 1803–12 (TNM F0033466); ibid., Jenai township, Aowanda National Forest Recreation Area, 23°57'N, 121°10'E, 1320 m, on angiosperm branch, 13 Sep 2018, leg. C.-L. Wei, WEI 18–439 (TNM F0034239); ibid., Luku township, Hsitou, 23°41'N, 120°48'E, 1250 m, on angiosperm branch, 11 Dec 2016, leg. S.-Z. Chen & C.-C. Chen, GC 1612-33 (TNM F0031437); ibid., GC 1612-37 (TNM F0031441). New Taipei City, Pinglin District, Taweishan Trail, 24°56'N, 121°43'E, 270 m, on angiosperm branch, 23 Mar 2018, leg. S.-Z. Chen, C.-C. Chen & C.-L. Wei, Chen 3891 (TNM F0033945); ibid., Wanli District, Laoliaohu, 25°10'N, 121°37'E, 470 m, on angiosperm branch, 23 May 2017, leg. Y.-L. Huang, C.-L. Wei & C.-C. Chen, WEI 17–204 (TNM F0031926); ibid., Wulai District, Neidong National Forest Recreation Area, 24°50'N, 121°32'E, 400 m, on fallen branch, 23 Jun 2011, leg. E.O. Yurchenko, EYu 110623-28b (TNM F0024881); ibid., Yangminshan, 25°9'N, 121°33'E, 600 m, on angiosperm branch, 10 Apr 1994, leg. S.H. Wu, Wu 9404-3 (TNM F0002377). Taichung City, Hoping District, Dasyueshan Forestry Road, 24°15'N, 120°55'E, 1250 m, on bamboo culm, 30 Mar 2017, leg. C.-C. Chen, GC 1703-113 (TNM F0031212). Taipei City, Peitou District, Tinghushan Hiking Trail, 25°9'N, 121°32'E, 410 m, on angiosperm branch, 2 Dec 2019, leg. C.-L. Wei, WEI 19-458 (TNM F0036289).
Ecology and distribution. On angiosperm branches or trunks, rotten stumps, or bamboo culms in China (Beijing, Chongqing, Fujian, Guangdong, Guangxi, Guizhou, Henan, Hubei, Hunan, Jiangsu, Jiangxi, Jilin, Liaoning, Yunnan) and Taiwan (Liu et al. 2022a; Luo and Zhao 2022; this study). Fruiting year-round and found at elevations ranging from 270 to 2040 m in Taiwan.
Notes. Trechispora sinensis, recently described from mainland China, is synonymized under T. odontioidea based on morphological and phylogenetic evidence (Fig. 1). T. odontioidea is characterized by cream to buff basidiomata with odontioid hymenophore, presence of bipyramidal crystals, and thin-walled, verrucose basidiospores measuring 2.8–3.3 × 2.5–2.9 μm (Liu et al. 2022a, b). In Taiwan, T. odontioidea is one of the most common Trechispora species. It may be confused with T. bambusicola C.L. Zhao, which also occurs on bamboo, and T. subsinensis. However, T. bambusicola differs by having thick-walled basidiospores and lacking crystals (Zhao and Zhao 2021), while T. subsinensis differs by having predominantly thick-walled tramal hyphae and aculeate basidiospores (Liu et al. 2022a, b).
Trechispora orchidophila Yi C. Lin & C. Chih Chen, sp. nov. Figure 11.Fig. 11Trechispora orchidophila (from holotype). A Basidioma. B Vertical section through basidioma. C Basidia. D Basidiospores.** E**–G Scanning electron micrographs of crystals in polytetrahedral forms (E) and rhomboidal flakes (F), as well as basidiospores (G). Scale bars: A = 1 mm; B = 10 μm; C–D = 5 μm; E–F = 1 μm; G = 0.5 μm
MycoBank MB857632.
Etymology. orchidophila (Lat.), referring to its association with orchids.
Diagnosis. Trechispora orchidophila is characterized by white to cream basidiomata with smooth hymenophore, presence of crystals in subiculum, and thin-walled, aculeate basidiospores.
Typification. TAIWAN. Nantou County, Jenai township, Huisun Forestry Station, 24°5'N, 121°2'E, 750 m, on rotten branch, 24 Mar 2017, leg. S.-H. Wu, Wu 1703–55 (holotype TNM F0031278). GenBank: ITS = PV085812; 28S = PV085824.
Description. Basidiomata annual, resupinate, thin, soft, fragile, easily separated from substratum, up to 8 cm long, 1 cm wide. Hymenophore smooth, farinaceous, white to cream. Margin white to cream, fimbriate. Hyphal system monomitic; generative hyphae with clamp connections. Subicular hyphae long-celled, colorless, thin-walled, moderately branched and septate, subparallel to interwoven, 2.5–5 μm in diam, ampullate septa usually present in the hyphae, up to 4.5 μm wide. Subhymenium hyphae distinct, colorless, thin-walled, much branched, smooth, 2.5–5 μm in diam. Crystals frequently occurring in subiculum as small, aggregated, polytetrahedral forms or rhomboidal flakes. Cystidia absent. Basidia subclavate to cylindrical, colorless, thin-walled, with two to four sterigmata and a basal clamp connection, 11.4–14.6 × 4.4–4.8 μm. Basidioles similar in shape to basidia, but smaller. Basidiospores ellipsoid, colorless, thin-walled, thick-walled when aged, aculeate, inamyloid, indextrinoid, (3.7–)3.9–4.7(–5.3) × (2.6–)2.9–3.5(–3.8) μm, L = 4.3 μm, W = 3.2 μm, Q = 1.2–1.6 (n = 30).
Ecology and distribution. On rotten branch in Taiwan. Mar. Also associated with orchid roots, including Orchidaceae sp. in Réunion (France), and the mycoheterotrophic orchid Erythrorchis altissima in Okinawa (Japan) (Martos et al. 2012; Ogura-Tsujita et al. 2018).
Notes. The ITS sequences of the holotype Trechispora orchidophila (Wu 1703-55) has 99% similarity with FM151.1 (GenBank: JF691276; 573/580) and Y453-2 (GenBank: LC327027; 574/580), both uncultured environmental samples from orchid roots. FM151.1 was detected in Orchidaceae sp. in Réunion of France, while Y453-2 was recovered from Erythrorchis altissima in Okinawa of Japan (Martos et al. 2012; Ogura-Tsujita et al. 2018). Phylogenetically, T orchidophila is closely related to T. malayana S.L. Liu, S.H. He & L.W. Zhou (Fig. 1). However, T. malayana has odontioid hymenophore and aculeate basidiospores.
Morphologically, Trechispora orchidophila resembles T. larssonii S.L. Liu, L.W. Zhou & S.H. He and T. latehypha S.L. Liu, S.H. He & L.W. Zhou in having white to cream basidiomata with smooth to grandinioid hymenophore, presence of rhomboidal crystals, and ellipsoid basidiospores. However, T. orchidophila differs by having longer basidiospores (T. larssonii: 2.8–3.3 μm*; T. cyatheae*: 3–3.5 μm) (Liu et al. 2022a; Ordynets et al. 2015).
Trechispora rigida (Berk.) K.H. Larss., Nordic J. Bot. 16:92. 1996. Figure 12.Fig. 12. Basidioma of Trechispora rigida (from Wu 0008–5). Scale bar = 1 mm
Description and illustration. Larsson (1996).
Specimens examined. CHINA. Liaoning Province, Fushun City, Qingyuan Manchu Autonomous County, Qingyuan ecological experiment station, 41°51'N, 124°56'E, 590 m, on angiosperm branch, 30 Jul 2017, S.-H. Wu, Wu 1707-127 (TNM F0031564). Yunnan Province, Hsishuangpanna, Green Stone Forest Park, 21°55'N, 101°17'E, 650 m, on branch of angiosperm, 18 Aug 1997, S.-H. Wu & S.-Z. Chen, Wu 9708–313 (TNM F0007941); ibid., Chuxiong Yi Autonomous Prefecture, Nanhua County, Tujie Town, 24°49'N, 100°46'E, 1915 m, on gymnosperm branch, 15 Aug 2017, C.-C. Chen, GC 1708–314 (TNM F0032823). TAIWAN. Nantou County, Huisun Forestry Station, 700 m, on angiosperm branch, 4 Jun 1999, S.-H. Wu, Wu 9901–20 (TNM F0010251); ibid., Tungpu village, 23°33'N, 120°56'E, on angiosperm trunk, 20 Sep 1995, 1600 m, S.-H. Wu & S.-Z. Chen, Wu 9509–26 (TNM F0004256); ibid., Shalihsienhsi Forest Road, 23°32'N, 120°55'E, 1350 m, on angiosperm branch, 24 Nov 1993, leg. S.-H. Wu & S.-Z. Chen, Wu 9311–78 (TNM F0001504). Pingtung County, Manchou township, Chufengshan, 22°2'N, 120°51'E, 90 m, on angiosperm branch, 29 Nov 2018, C.-C. Chen & C.-L. Wei, WEI 18–606 (TNM F0034700); ibid., Wanliteshan, 22°4′33"N, 120°50′12"E, 230 m, on angiosperm branch, 29 Nov 2018, C.-C. Chen & C.-L. Wei, WEI 18–679 (TNM F0034741). Taitung County, Orchid Island, Chungai Bridge, on rotten angiosperm branch, 50 m, 30 Apr 1997, S.-H. Wu & J.Y. Tseng, Wu 9704-128 (TNM F0008677); ibid., Taimali, 850 m, on angiosperm branch, S.-H. Wu, Wu 890519-27 (TNM F0014986). Taipei City, Yangminshan National Park, Luchiaokenghsi Ecological Reserve, 25°11'N, 121°34'E, 350 m, on rotten angiosperm branch, 21 Oct 2001, S.-H. Wu et al., Wu 0008-5 (TNM F0012765).
Ecology and distribution. On angiosperm and gymnosperm wood in Argentina, Brazil, China (Liaoning, Yunnan), and Taiwan (Larsson 1996; Hjortstam and Ryvarden 2007; Silveira 2016; this study). Jun, Apr, Sep to Nov in Taiwan.
Notes. Trechispora rigida is characterized by white to cream basidiomata with grandinioid hymenophore and aculeate, concave basidiospores, originally measured as 4.5–5.5 × 4 μm by Larsson (1996). The examined specimens from mainland China and Taiwan share these morphological features, though their basidiospores were slightly smaller (4–4.5 × 3–3.5 μm). This species was first reported from Taiwan by Wu (2003).
Trechispora subsinensis S.L. Liu, S.H. He & L.W. Zhou, Mycosphere 13:931. 2022. Figure 13.Fig. 13Trechispora subsinensis (from GC 2309–119). A–B Basidiomata. C Scanning electron micrograph of basidiospores. Scale bars: A–B = 1 mm; C = 0.5 μm
Description and illustration. See Liu et al. (2022a).
Specimens examined. CHINA. Guangxi Province, Dayaoshan Nature Reserve, Hekou Protection Station, 24°8'N, 110°5'E, 620 m, on angiosperm trunk, 9 Sep 2018, leg. S.-H. Wu, Wu 1809-21 (TNM F0032915). TAIWAN. Kaohsiung City, Maolin District, Shan-Ping Forest Ecological Garden, 22°58'N, 120°41'E, 730 m, 29 Jun 2017, on rotten wood, leg. C.-C. Chen, C.-L. Wei, W.-C. Chen & Y.-P. Chen, WEI 17-359 (TNM F0032470); ibid., on angiosperm branch, leg. C.-C. Chen, C.-L. Wei, W.-C. Chen, & Y.-P. Chen, WEI 17-401 (TNM F0032489). New Taipei City, Pinghsi District, Lingjiaoliao Mountain Trail, 25°2'N, 121°45'E, 210 m, on stem of Arenga engleri, 23 Aug 2018, leg. S.-Z. Chen, C.-C. Chen, & C.-L. Wei, Chen 3821 (TNM F0033911); ibid., Wulai Distric, Kalamoji Trail, 24°47'N, 121°30'E, 380 m, on angiosperm branch, 23 Jan 2019, leg. C.-C. Chen & C.-L. Wei, WEI 19-011 (TNM F0034764). Pingtung county, Manchou township, Chufengshan, 22°4'N, 120°51'E, 120 m, on rotten angiosperm trunk, 25 Apr, 2007, leg. S.-H. Wu, S.-Z. Chen & D.-M. Wang, Wu 0704–14 (TNM F0020980); ibid., Nanjenshan, 22°5'N, 120°52'E, 320 m, on angiosperm branch, 28 Nov 2018, leg. C.-C. Chen & C.-L. Wei, WEI 18-531 (TNM F0034466). Taichung City, Hoping District, Wuwoweishan, 24°14'N, 120°58'E, 1960 m, on angiosperm branch, 26 Oct 2017, leg. C.-L. Wei & Y.-L. Huang, WEI 17-717 (TNM F0032718). Taitung county, Orchid Island, 22°00'N, 121°34'E, 87 m, on angiosperm branch, leg. C.-C. Chen, S.-Z. Chen & S.-W. Chou, 22 Sep 2023, GC 2309-119 (TNM F0038420).
Ecology and distribution. On angiosperm (e.g., Arenga) branches or trunks in China (Guangdong, Guangxi), Thailand (Chiang Mai), and Taiwan (Liu et al. 2022a; this study). Occurs at elevations from 87 to 1960 m, recorded in Jan, Apr, Jun to Nov in Taiwan.
Notes. Trechispora subsinensis is newly recorded from Taiwan and is characterized by white to cream basidiomata with an odontioid hymenophore, thick-walled tramal hyphae, and aculeate basidiospores measuring 3–4 × 3–3.5 μm. The basidiospores of the examined specimens in this study are more globose and larger than those of the holotype (2.7–3.5 × 2.3–2.8 μm) (Liu et al. 2022a). Morphologically, T. subsinensis resembles T. crystallina and T. odontioidea due to its grandinioid to odontioid hymenophore and similar basidiospore size. However, T. crystallina differs by its thin-walled tramal hyphae and verrucose basidiospores, while T. odontioidea is distinguished by its verrucose basidiospores (Liu et al. 2022a).
Trechispora taiwanensis S.L. Liu, S.H. He & L.W. Zhou, Mycosphere 13:934. 2022. Figure 14.Fig. 14. Basidioma of Trechispora taiwanensis (from GC 1703–113). Scale bar = 1 mm
Description and illustration. See Liu et al. (2022a).
Specimens examined. TAIWAN. Nantou County, Lianhuachi, on dead bamboo, 6 Dec 2016, leg. S.H. He, He 4571 (BJFC 024012, holotype). Taichung City, Hoping District, Dasyueshan Forestry Road. 24°15'N, 120°55'E, 1250 m, on bamboo culm, 30 Mar 2017, leg. C.-C. Chen, GC 1703-113 (TNM F0031212).
Ecology and distribution. On bamboo culm in Taiwan. Mar, Dec (Liu et al. 2022a; this study).
Notes. Trechispora taiwanensis is characterized by white to cream basidiomata with smooth to grandinioid hymenophore, ellipsoid, aculeate basidiospores measuring 3–4 × 2–2.8 μm (Liu et al. 2022a). It may be confused with T. latehypha and T. odontioidea, which also occur on bamboo. However, T. latehypha differs by its smooth hymenophore and thick-walled generative hyphae, while T. odontioidea has an odontioid hymenophore and verrucose basidiospores (Liu et al. 2022a).
Trechispora wenshanensis K.Y. Luo & C.L. Zhao, MycoKeys 105:167. 2024. Figure 15.Fig. 15Trechispora wenshanensis (from WEI 20–112). A Basidioma. B–C Scanning electron micrographs of irregular crystals (A) and basidiospores (B). Scale bars: A = 1 mm; B = 1 μm C = 0.5 μm
Description and illustration. See Luo et al. (2024).
Specimen examined. TAIWAN. Taoyuan City, Fuxing District, on the way to Sileng Hot Spring, 24°38'N, 121°26'E, 1140 m, on angiosperm trunk, 12 Jul 2020, leg. C.-L. Wei, WEI 20-112 (TNM F0038414).
Ecology and distribution. On angiosperm branches and trunks in SW China (Yunnan) and Taiwan. Jul in Taiwan.
Notes. Trechispora wenshanensis is characterized by cream to buff basidiomata with smooth hymenophore, narrow generative hyphae (1–2 μm in diam), and aculeate basidiospores measuring 2.5–3.7 × 2–3 μm (Luo et al. 2024). The Taiwanese specimen has longer basidiospores (3.6–4.2 μm) and small, aggregated, irregular crystals in subhymenium (Fig. 15B). Trechispora wenshanensis is newly recorded from Taiwan.
Key to known species of Trechispora from Taiwan
- Basidiomata clavarioid, on ground…*Trechispora cryptomerioides *Basidiomata resupinate, on wood…2
- Hymenophore poroid…3 Hymenophore non-poroid…5
- Hymenial surface white ivory pale yellow, subicular hyphae slightly thick-walled, up to 3 μm wide…4 Hymenial surface brick-red, subicular hyphae thin-walled, up to 6 μm wide…T. formosana
- Basidiospores subglobose, < 4 μm long…*T. mollusca *Basidiospores ellipsoid, > 4 μm long…T. dentata
- Hymenophore smooth…6 Hymenophore non-smooth…12
- Hyphal system dimitic…*T. dimitica *Hyphal system monomitic…7
- Crystals in subiculum acicular…8 Crystals in subiculum differently shaped…9
- Basidiospore including spines > 5 μm long, > 4 μm wide…*T. praefocata *Basidiospore < 4 μm long, < 3 μm wide…T acerosa
- Crystals petaliform, basidiospores 4.1–4.8 × 3.1–3.8 μm…*T. floralis *Crystals in shape of rhomboidal flakes or absent…10
- Subhymenial hyphae thick-walled…*T. latehypha *Subhymenial hyphae thin-walled…11
- Subhymenial hyphae < 2 μm wide…*T. wenshanensis *Subhymenial hyphae > 2 μm wide…T. orchidophila
- Hymenophore colliculose or grandinioid; tramal hyphae thin-walled or slightly thick-walled…13Hymenophore odontoid to hydnoid; tramal hyphae distinctly thick-walled…16
- On bamboo…*T. taiwanensis *On other substrata…14
- Basidiospores ventrally concave…*T. rigida. *Basidiospores ventrally straight or convex…15
- Basidiospores verrucose…*T. crystallina *Basidiospores aculeate…T. farinacea
- Basidiospores verrucose…T. odontioideaBasidiospores aculeate…T. subsinensis
Discussion
Taxonomy and diversity of Trechispora in Taiwan
Taxonomic studies on the genus Trechispora have traditionally focused on Europe and the Americas (Karsten 1890; Larsson 1992, 1996; Chikowski et al. 2020), with more recent research expanding into the Indo-Pacific region (Ordynets et al. 2015; Liu et al. 2022a; Sommai et al. 2023). This study provides the first comprehensive revision of Trechispora in Taiwan, integrating morphological and molecular data.
Prior to this study, only seven species had been recorded in Taiwan (Lin et al. 2022; Liu et al. 2022a; Maekawa 1992; Wu 2003), but molecular data were unavailable for some of them, leaving their phylogenetic placements uncertain. We confirmed the taxonomic status of T. cryptomerioides, T. rigida, and T. taiwanensis based on morphological re-examinations, while T. lunata was previously transferred to Sidera as S. lunata (Romell ex Bourdot & Galzin) K.H. Larss. (Miettinen and Larsson 2011). However, Taiwanese T. rigida remains phylogenetically unresolved due to the lack of molecular data. T. dimitica, T. farinacea, and T. praefocata, originally reported from Orchid Island, were not recollected in this study, and their current distribution requires further investigation.
This study expands the known diversity of Trechispora in Taiwan, increasing the number of recognized species to 17. Four new species (T. acerosa, T. floralis, T. formosana, and T. orchidophila) are described, and seven species are newly recorded for Taiwan (T. crystallina, T. dentata, T. latehypha, T. mollusca, T. odontioidea, T. subsinensis, and T. wenshanensis). These findings highlight Taiwan as a previously underexplored region for Trechispora and suggest that additional undiscovered species may be present.
Ecological significance of Trechispora in Taiwan
Our findings indicate that Trechispora species in Taiwan inhabit a broad range of substrates, including dead or living angiosperm and gymnosperm wood, as well as soil. Among them, T. odontioidea and T. subsinensis are the most frequently encountered species, occurring year-round on angiosperm wood and bamboo culms from lowland areas to around 2000 m. The ecological role of T. dentata, originally described from soil in Yunnan (Liu et al. 2022b) but found in Taiwan on highly decomposed wood, remains unclear. A similar pattern is observed in T. incisa (Larsson 1992), suggesting that these species may not function as true wood-decay fungi but rather as soil- and wood-inhabiting fungi with alternative ecological strategies.
A unique case is T. cryptomerioides, the only known coralloid Trechispora species in Taiwan. Unlike other species that primarily inhabit wood, T. cryptomerioides is found on the ground in Cryptomeria japonica-dominated forests at 1300–1950 m. DNA of T. cryptomerioides has been detected in C. japonica root systems (Lin et al., unpublished data), and its fruiting is influenced by stand thinning (Lin et al. 2015). These findings suggest a close association between T. cryptomerioides and C. japonica, raising the possibility of a mycorrhizal relationship, though further studies are required to confirm this interaction (Lin et al. 2022).
Another noteworthy species is T. orchidophila, which was collected from rotten wood but is phylogenetically linked to environmental samples from orchid roots in Réunion (France) and Okinawa (Japan) (Martos et al. 2012; Ogura-Tsujita et al. 2018). This suggests that T. orchidophila may function as a saprotroph or root endophyte, potentially forming symbiotic associations with orchids, although additional studies are needed to clarify its ecological role.
Although Trechispora species are known to associate with termites, no termite-associated species have been recorded in Taiwan. In other regions, T. polygonospora Ryvarden and T. termitophila Meiras-Ottoni & Gibertoni have been found in termite mounds (de Meiras-Ottoni et al. 2021; Larsson 1992), and certain species produce termite egg-mimicking structures (termite balls) within termite nests (Matsuura and Yashiro 2010). Future surveys of termite nests and decomposing wood in Taiwan’s tropical and subtropical forests may reveal new termite-associated Trechispora species.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Larsson KH (1992) The genus Trechispora (Corticiaceae, Basidiomycetes). University of Gothenburg, Department of Systematic Botany, Dissertation
- 2Silveira RMB (2016) Trechisporales in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB 95394