Claustrophilic oviposition: Oviposition performance depends on container size in a novel forced oviposition method for Culex quinquefasciatus and Aedes aegypti
Kendra A. Dagg, Alden S. Estep, Cason E. Bartz, Edwin R. Burgess I.V., Adly M.M. Abd-Alla, Adly M.M. Abd-Alla, Adly M.M. Abd-Alla

TL;DR
A new method for forcing mosquitoes to lay eggs in small containers is effective for two species, improving lab research and colony establishment.
Contribution
A modified forced oviposition method is introduced and tested for Aedes aegypti and Culex quinquefasciatus, showing species-specific efficacy and claustrophilic oviposition behavior.
Findings
Aedes aegypti showed higher oviposition success (>80%) compared to Culex quinquefasciatus (50–60%).
Both species preferred smaller oviposition tubes (1.5 and 5 mL) for laying eggs.
The method successfully produced F1 progeny from both lab and wild-caught mosquitoes.
Abstract
Mosquito vector control research relies heavily on the use of live organisms, with wild populations playing a critical role in surveillance, field product validation, and strengthening the overall efficacy and accuracy of studies. However, too often wild mosquito rearing and colony establishment in laboratory settings is consistently challenging. Here we present a modified version of a forced oviposition method for use against two different mosquito species, Aedes aegypti and Culex quinquefasciatus. The efficacy of the technique was tested with both laboratory and wild-caught strains placed in oviposition tubes consisting of a 1.5 mL tube containing moistened cotton and a strip of germination paper. To determine if size had an impact on oviposition rate, an additional test was conducted using four different size oviposition tubes; 1.5, 5, 15, and 50 mL. Overall, the forced oviposition…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
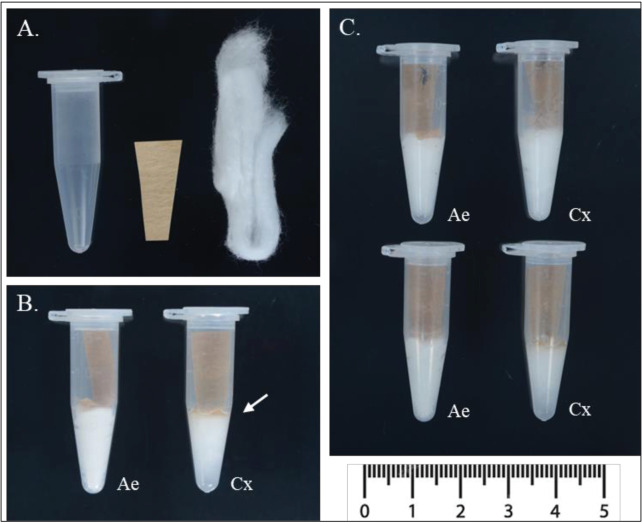 Fig 1
Fig 1 Fig 2
Fig 2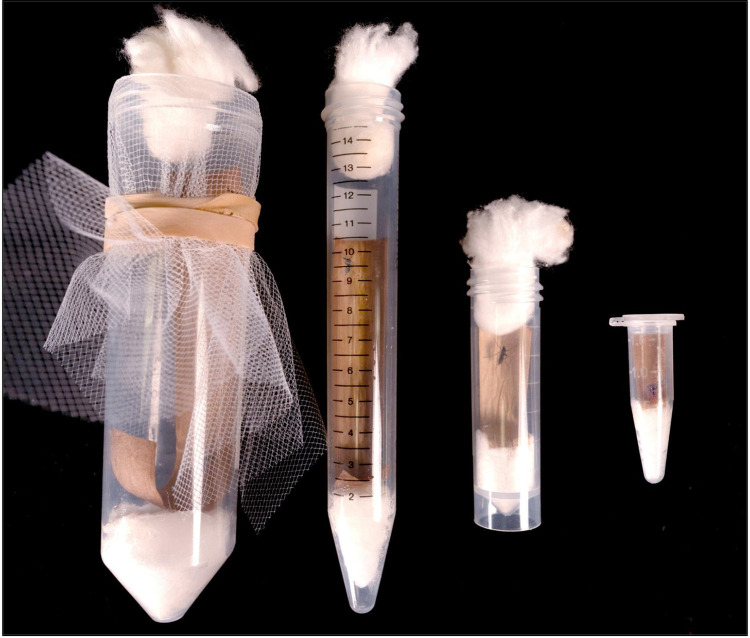 Fig 3
Fig 3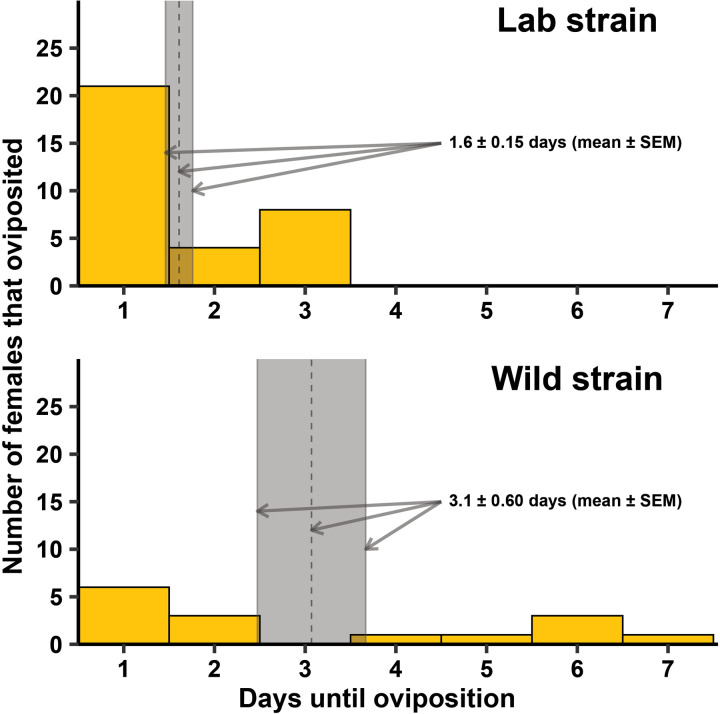 Fig 4
Fig 4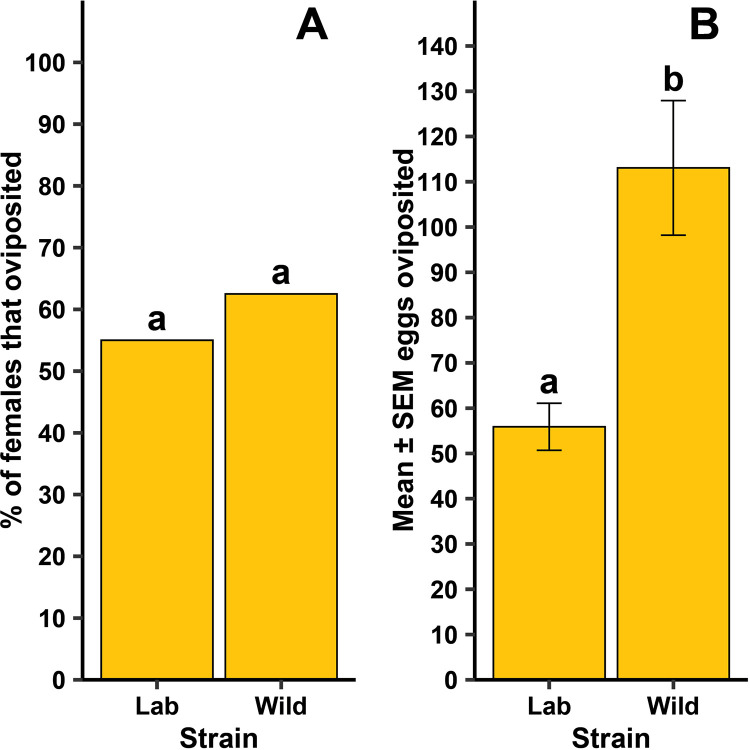 Fig 5
Fig 5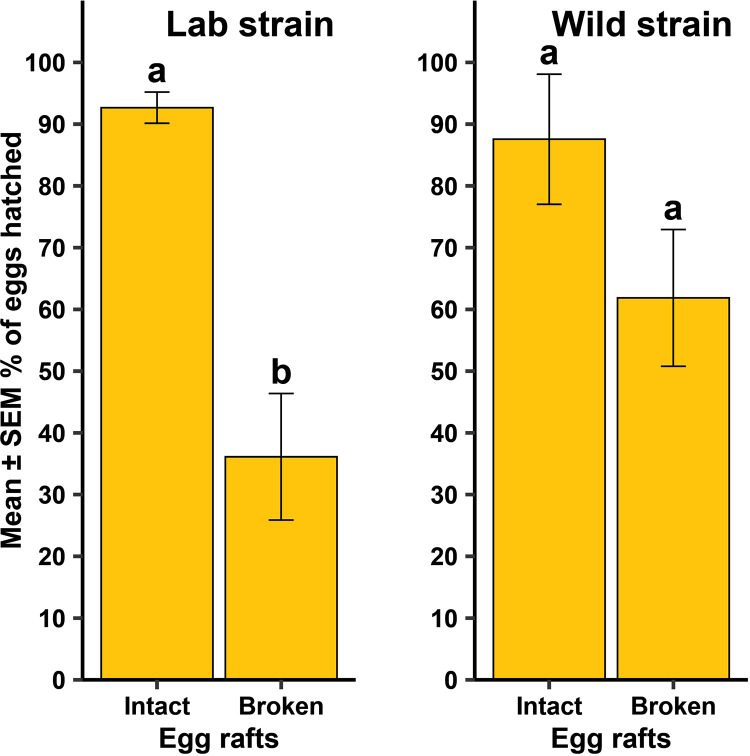 Fig 6
Fig 6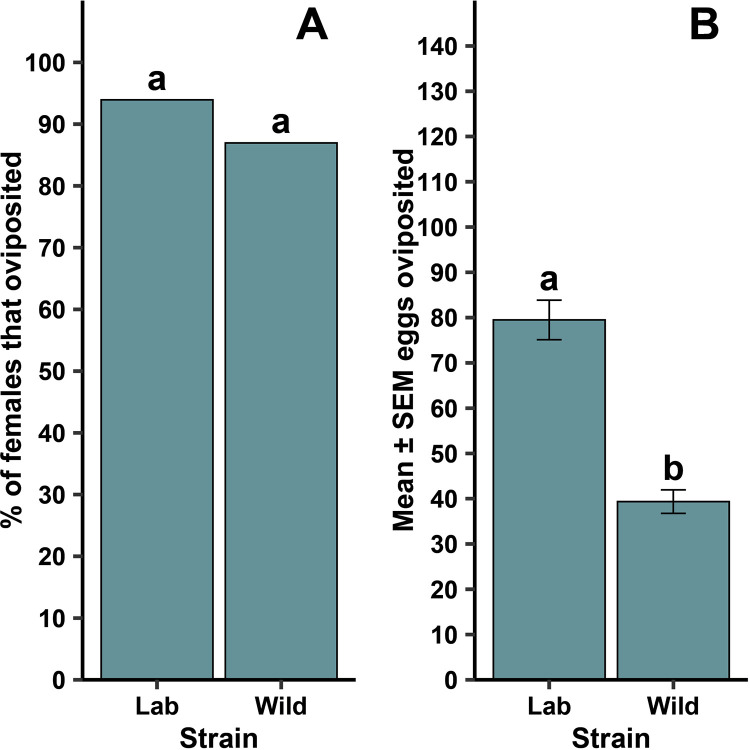 Fig 7
Fig 7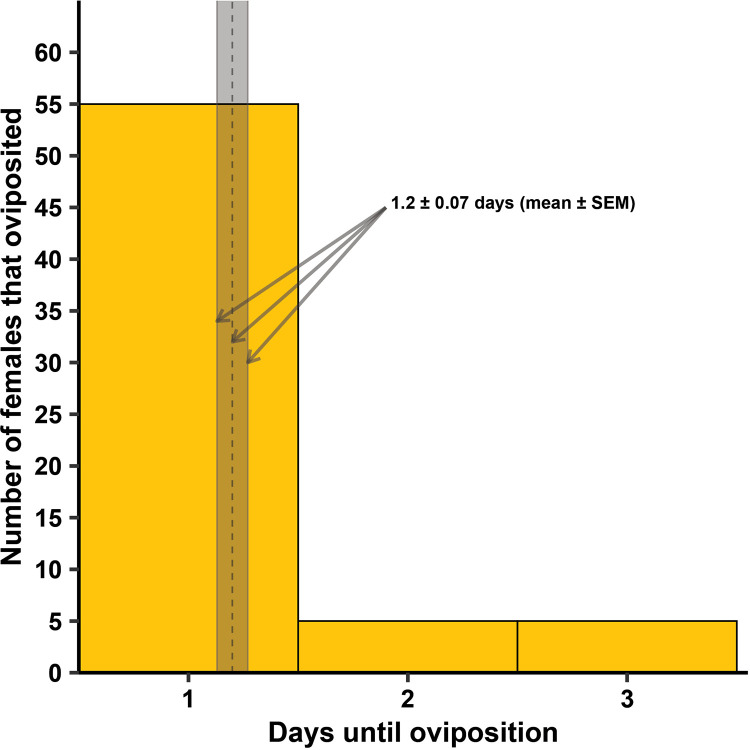 Fig 8
Fig 8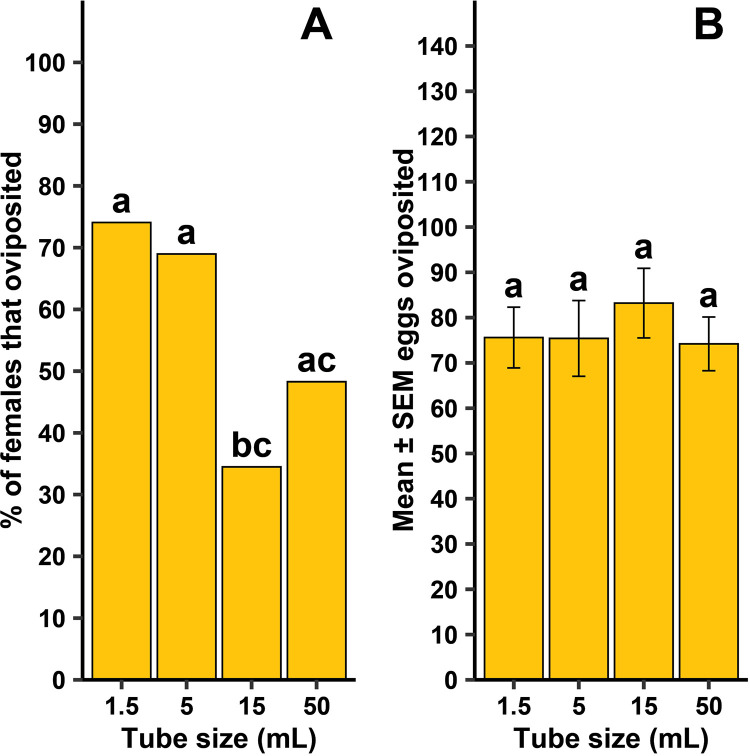 Fig 9
Fig 9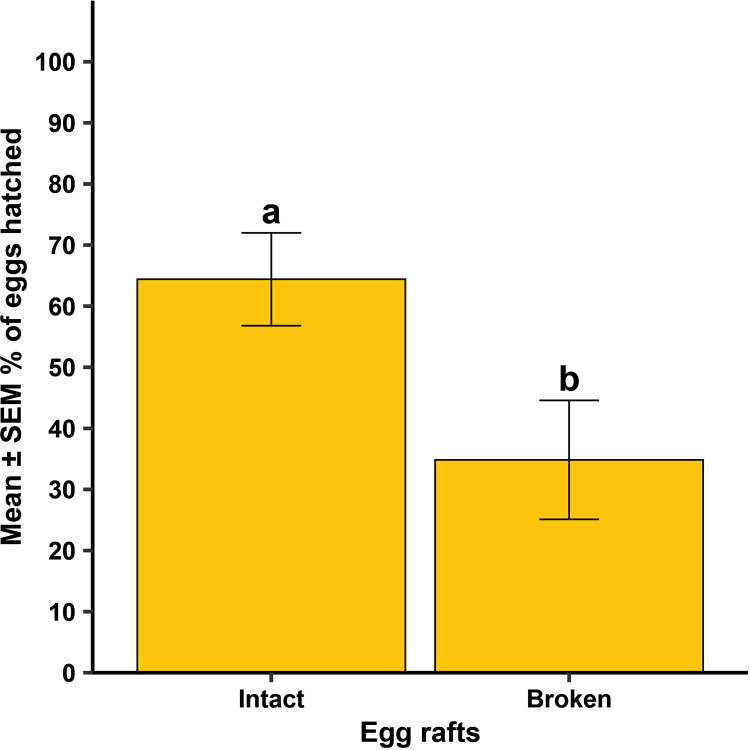 Fig 10
Fig 10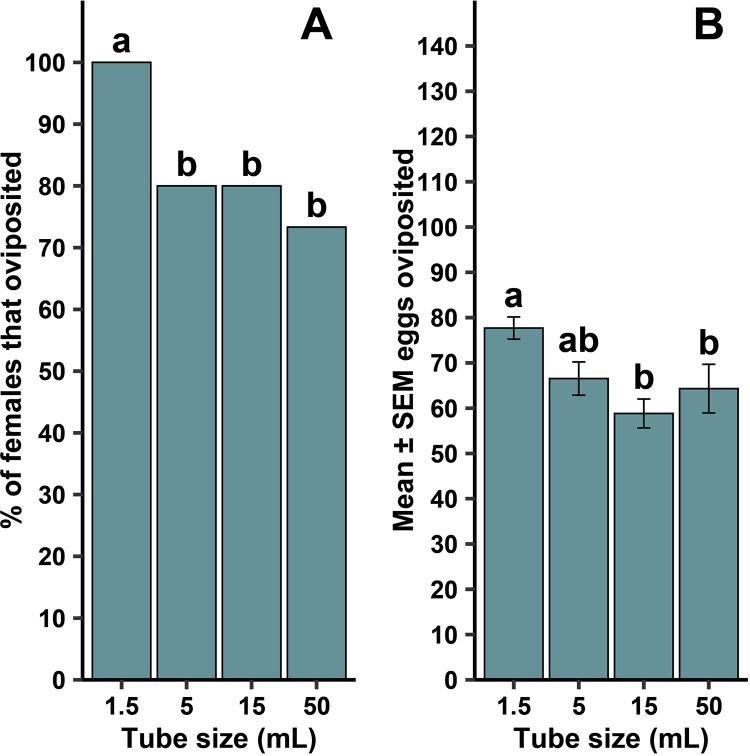 Fig 11
Fig 11- —http://dx.doi.org/10.13039/100011508Florida Department of Agriculture and Consumer Services
- —http://dx.doi.org/10.13039/100000005U.S. Department of Defense
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMosquito-borne diseases and control · Malaria Research and Control · Insect Pest Control Strategies
Introduction
Vector-borne disease (VBD), and particularly mosquito-borne disease, such as malaria, dengue virus, and West Nile virus, are major causes of morbidity and mortality [1]. In 2023, the World Health Organization (WHO) estimated that VBDs accounted for 17% of all reported global infectious disease cases [2]. Despite continual efforts to reduce this global disease burden, an increase in disease incidence is thought to be due to a combination of factors, such as shifts in climate, land-use practices, international travel and trade, and urbanization, all of which affect both the pathogen and their associated vector [1]. The number of reported malaria cases in 2023 surpassed that of 2022 by nearly 11 million cases [3]. By April 2024, over 7.6 million dengue cases were reported from roughly 86 countries/territories, which was three times the number reported in 2023 [4]. While substantial medical advances (e.g. vaccine development) have contributed to case management, costs of medical care, supply limitations, and constraints on eligibility for medical treatment (e.g. age or medical status) restrict access to care [5]. To date, vector control remains the principal method of any vector-borne disease control program, and in some cases, vector control is currently the only method available for disease prevention [6,7].
The development and testing of mosquito vector control tools and strategies relies heavily on research conducted on the target mosquito species itself. The use of live organisms is vital for research on mosquito behavior, biology, physiology, product testing, insecticide resistance, vector incrimination, and vector competency [7–9]. Additionally, wild collected populations play a pivotal role by providing insight into the field applicability of methods and strengthening the overall efficacy and accuracy of studies. In order to properly conduct mosquito research, large numbers of individual organisms are required to ensure the robustness of the dataset. While most strains of mosquitoes are hardy and resilient in their natural habitat, wild mosquito rearing and colony establishment in laboratory settings is consistently challenging [8]. Quite often, the most common barriers to insectary rearing of wild mosquitoes are insufficient numbers of females collected from the field, and a general difficulty to induce oviposition from field blood-fed or gravid females [9].
To address these rearing challenges, we sought to modify an oviposition encouragement technique originally developed by Morgan et al. [10] for establishing laboratory colonies of field-caught Anopheles spp., for use with two different mosquito species of public health importance, Culex quinquefasciatus and Aedes aegypti. Both vector species have distinct oviposition habitat preferences and oviposition behaviors. The Morgan et. al [10] method has been successfully used to establish wild laboratory colonies for multiple Anopheles spp. in Thailand, Ethiopia, Madagascar, and Uganda [9–12]. However, there is no literature on similar techniques available used to establish colonies of other mosquito vector species. Here we demonstrated that with slight modifications, this method was effective at producing high numbers of viable Cx. quinquefasciatus and Ae. aegypti eggs from both established laboratory strains as well as F_1_ progeny from field-caught female mosquitoes. Additionally, Morgan et al. (2010) [10] hypothesized that the success of the method could be related to the confined space the mosquitoes are put in, which may suggest an affinity for tight spaces (i.e., claustrophilic) when ovipositing. To test this hypothesis, we investigated if holding container size had an impact on oviposition rate and egg production in both species through the use of four different sized oviposition containers that are commonly available in laboratories and mosquito control districts.
Materials and methods
Mosquito strains and rearing
Laboratory reared strains and locally collected wild populations of Cx. quinquefasciatus and Ae. aegypti were used in this study. All laboratory strains were provided by the Center for Medical, Agricultural & Veterinary Entomology (CMAVE), U.S. Department of Agriculture (Gainesville, FL, USA). Laboratory strains used for this study were the Cx. quinquefasciatus CMAVE strain and the Ae. aegypti ‘Orlando’ (ORL) strain, each referred to hereafter as the “lab strain” for their respective species. Laboratory strains were reared using standardized protocols previously described [13]. Wild caught mosquitoes were supplied as adult blood engorged or gravid females for both species by Gainesville Public Works Department, Mosquito Control (Gainesville, FL, USA). Mosquitoes were collected by the city Mosquito Control Department in July 2024 within the Gainesville, FL city area and were morphologically identified using keys from Darsie et al. [14]. Gravid females were held for 24 hrs under standard insectary conditions (27°C, 70% relative humidity, light:dark cycle 12:12 hrs), and blood engorged females were held for 96 hrs before use in any experimentation.
Experiment 1: Forced oviposition performance with recently collected field strains
Eggs were generated using a modified version of the forced-oviposition method originally designed for field caught Anopheles spp. described by Morgan et al. [10]. Modifications to the Anopheles spp. method were based on genus oviposition behavior and ease of equipment accessibility in an insectary. Each oviposition container consisted of a 1.5 mL Eppendorf tube, moistened cotton, and a strip of germination paper (Fig 1A). A small hole was made in the cap of the tube to allow air flow during the holding period. Cotton balls were broken into smaller pieces and were moistened using Gainesville city tap water. At the bottom of the 1.5 mL tube, a single small piece of moistened cotton was pushed down into the conical tip of the tube. Once pushed down, the cotton filled no more than the conical tip of the 1.5 mL tube and all excess water was removed. A roughly 2 x 1 cm strip of germination paper, with a tapered end (forming a triangle like shape) was inserted tip first into the tube. The inserted end of the germination paper was pushed down to fold over the surface of the moisten cotton (Fig 1B). For both laboratory and wild Ae. aegypti, excess water was completely removed from the bottom of the tube, however, for Cx. quinquefasciatus a thin layer of water was left at the bottom of the tube (approx. 1mm at the meniscus) (Fig 1B-C). Individual females that had previously been blood fed were gently aspirated into a prepared 1.5 mL Eppendorf tube and the cap was secured. Oviposition tubes were maintained in standard insectary conditions (27°C, 70% relative humidity, light:dark cycle 12:12 hrs) for the designated holding period. Aedes aegypti strains were checked at 24 hrs and 48 hrs, while the Cx quinquefasciatus strains were checked every 24 hrs for 168 hrs (7 days).
Layout of equipment and examples of complete set up of modified forced oviposition technique for Aedes aegypti (Ae) and Culex quinquefasciatus (Cx).(A) Oviposition containers consist of a 1.5 mL Eppendorf tube, a small piece of germination paper cut with a small taper at the end, and a piece of cotton. (B) Moistened cotton is pushed to the bottom of the tube and germination paper placed on top with only the tapered end covering the cotton. Excess water is removed from Ae tubes, with a ~ 1 mm water layer remaining (white arrow) only for the Cx tube set up. (C) A single female is aspirated into each container (Photo credit: Kendra A. Dagg).
Variables collected included: 1) did the female oviposit (yes/no) (Fig 2A-B), 2) number of eggs laid, and 3) number of days between transfer to oviposition tube to laid eggs. Oviposition was determined through visual examination of germination papers and cotton under a dissecting microscope (Stemi 2000, Zeiss International, Germany). If females did not oviposit within the time period, they were dissected to check for gravidity (yes/no), i.e., to determine if they had eggs but did not oviposit. To check the viability of eggs, 20–25 randomly selected eggs from each female were transferred to a 35 mm diameter plastic Petri dish. If a female laid <20 eggs, all eggs available were hatched. Each dish was filled with 5 mL of tap water and 20 µL of a larval food slurry (approx. 150 mg of ground fish food flakes, mixed with 1.5 mL of tap water). Dishes were held in standard insectary conditions and checked every 24 hrs for 72 hrs. The total number of eggs hatched was recorded for each dish at each timepoint. Any hatched larvae were reared up to the L2 stage for ease of visualization.
Example egg papers collected from oviposition tubes.Eggs were collected between 24 – 168 hr after female transferred into oviposition tube for Culex quinquefasciatus (A) and 24 hr for Aedes aegypti (B) (Photo credit: Kendra A. Dagg).
For Cx. quinquefasciatus, the lab strain was replicated 13–18 times across each of four generational cohorts (N = 60) and the wild strain was replicated 11–13 times across two generational cohorts (N = 24). For Ae. aegypti, the lab strain was replicated 9 – 13 times across three generational cohorts (N = 33) and the wild strain was replicated 10 – 13 times across two generational cohorts (N = 23). This replication structure represents all individual females that met three conditions (in order): 1) blood fed, 2) alive at the end of the assay, 3) gravid. Any females that did not meet all three conditions were excluded from the analysis.
Experiment 2: Tube size effects on forced oviposition performance
To determine if oviposition tube size influenced oviposition rate and egg numbers, oviposition tubes were prepared in four different sizes: 1.5 mL Eppendorf tubes, 5 mL polypropylene centrifuge tube, 15 mL polypropylene centrifuge tube, and 50 mL polypropylene centrifuge tube. The additional tube sizes were set up using the same equipment as previously described and capped with either a piece of cotton (5 mL tubes) or a piece of mesh secured by a rubber band (15 mL and 50 mL tubes) (Fig 3). Moistened cotton pieces and germination paper size was prepared proportional to the oviposition tube size. Gravid lab strain females were individually aspirated into an oviposition tube and stored in standard insectary conditions (27°C, 70% relative humidity, light:dark cycle 12:12 hrs). Oviposition was observed for the Ae. aegypti lab strain and the Cx. quinquefasciatus lab strain every 24 hrs for 48 and 96 hrs, respectively. The same data variables were recorded as described in the forced oviposition assay (Experiment 1).
Impact of tube size on oviposition performance.Four different tube sizes were used to determine if tube size affected oviposition performance; (from left to right) 50 mL, 15 mL, 5 mL, and 1.5 mL (Photo credit: Kendra A. Dagg).
For Cx. quinquefasciatus, all four tube sizes were replicated 8 – 10 times across each of three generational cohorts (N = 114). For Ae. aegypti, all four tube sizes were replicated 15 times across two generational cohorts (N = 120). This replication structure represents the same criteria for inclusion in the analysis as described in Experiment 1.
Statistical analyses
All analyses were conducted in R version 4.4.2 (R Core Team 2024) and statistical significance was set to α = 0.05. For Experiment 1, analysis was done in the following order for Cx. quinquefasciatus: 1) a generalized linear model (quasipoisson) was used to assess if there was a difference in the time to oviposit (dependent variable) between the lab and wild strains (independent variable). 2) A 2x2 Chi-square test of independence with Yates correction for continuity was done to assess differences in frequency of oviposition between the two strains. 3) To determine if there was a strain effect (independent variable) on the number of eggs laid by females (dependent variable), a general linear model was used. 4) To determine if there was a difference in hatch rate (dependent variable) by strain (independent variable), a generalized linear model (quasibinomial) was used. 5) Finally, the oneway.test function, which controls for heterogeneity of variance, was used to determine if there was an effect of egg rafts being broken up vs. intact (independent variable) on the hatch rate (dependent variable). Because this test is a one-way ANOVA, strains had to be analyzed separately. For Experiment 1, analysis of Ae. aegypti was identical to the Cx. quinquefasciatus analysis except that all Ae. aegypti females oviposited within 1 day, and they lay eggs singly rather than in rafts, thus only analyses 2 – 4 were done on Ae. aegypti. For Experiment 2, all analyses were identical to Experiment 1 except that the independent variable of ‘strain’ was replaced with ‘tube size’ and Chi-squares used Yates correction for continuity (unnecessary for 4x2 tables). All models were diagnosed for departures from statistical assumptions, included Q-Q plots of residuals, and tests for homogeneity of variances. When Chi-square expected cell means were less than 5, Fisher’s exact test was used instead [15]. Where applicable, in the event the omnibus Chi-square test of independence or Fisher’s exact test was positive, pairwise comparisons were done using Fisher’s exact test. For generalized and general linear models, if overall significance was observed, and where applicable, posthoc pairwise comparisons were done with the ‘emmeans’ package [16].
Results
Experiment 1: Forced oviposition performance with recently collected field strains
For Cx. quinquefasciatus, there was a significant difference between the strains in the time it took for females to oviposit (Fig 4; F = 11.68, df = 1, 446, P = 0.001), but whether females oviposited at all was independent of strain (Fig 5A; P = 0.629). When females did oviposit, there was a significant difference between the strains in the mean number of eggs oviposited (Fig 5B; F = 20.72, df = 1, 46, P < 0.001). There was no difference between the two strains in the number of eggs that hatched (Table 1; F = 0.37, df = 1, 46, P = 0.544). For the lab strain, there was a significant difference in the percentage of eggs hatching between intact and broken up egg rafts (Fig 6A; F = 28.73, df = 1, 13.48, P < 0.001), while no difference was observed in the wild strain (Fig 6B; F = 2.82, df = 1, 12.79, P = 0.117).
Table 1: Mean ± standard error of the mean (SEM) number and percentage of eggs hatched relative to number tested in two strains of Culex quinquefasciatus (CMAVE – lab strain; wild – field collected strain) and two strains of Aedes aegypti (ORL – lab strain; wild – field collected strain).
Number of female lab strain Cx. quinquefasciatus that oviposited between 1 and 7 days in Experiment 1, utilizing a lab strain and a wild strain.There was a statistically significant difference between the strains at α = 0.05, so mean ± standard error of the mean (SEM) was calculated for each strain separately. The mean days to oviposition was 1.6 ± 0.15 for the lab strain and 3.1 ± 0.60 for the wild strain (gray columns in the figures represent SEMs, with the dashed lines in the middle representing the means). The lab strain was replicated 13-18 times across each of four generational cohorts (N = 60) and the wild strain was replicated 11-13 times across two generational cohorts (N = 24).
Differences for forced oviposition success frequency (A) and mean ± standard error of the mean (SEM) eggs when lab and wild strain female Cx. quinquefasciatus oviposited (B).Different lowercase letters above bars indicate statistical significance of pairwise comparisons between strains, with significance set at α = 0.05. The lab strain was replicated 13-18 times across each of four generational cohorts (N = 60) and the wild strain was replicated 11-13 times across each of two generational cohorts (N = 24).
Mean ± standard error of the mean (SEM) % eggs hatched when egg rafts from a lab strain and a wild strain of Culex quinquefasciatus were observed broken up versus being a single, intact egg raft.Different lowercase letters above error bars indicate statistical significance of pairwise comparisons between the egg raft status, with significance set at α = 0.05. The lab strain was replicated 13-18 times across each of four generational cohorts (N = 60) and the wild strain was replicated 11-13 times across two generational cohorts (N = 24).
For the lab and wild Ae. aegypti, all females oviposited within 1 day of being added to the tube (Fig 3). Whether females laid eggs at all was independent of strain (Fig 7A; P = 0.392). When oviposition occurred, there was a significant difference in the mean number of eggs oviposited between the lab and wild strains (Fig 7B; F = 47.48, df = 1, 49, P < 0.001). There was no difference between strains in the number of eggs that hatched (Table 1; F = 0.14, df = 1, 49, P = 0.707). All females, regardless of strain, oviposited within 1 day of being added to the tube.
Differences for forced oviposition success frequency (A) and mean ± standard error of the mean (SEM) eggs when lab and wild strain female Ae. aegypti oviposited (B).Different lowercase letters above bars indicate statistical significance of pairwise comparisons between strains, with significance set at α = 0.05. The lab strain was replicated 9 – 13 times across three generational cohorts (N = 33) and the wild strain was replicated 10 – 13 across two generational cohorts (N = 23).
Experiment 2: Tube size effects on forced oviposition performance
For lab strain Cx. quinquefasciatus there was no difference in the number of days until oviposition occurred among the tube sizes (Fig 8; F = 0.25, df = 3, 61, P = 0.864). The lab strain Cx. quinquefasciatus took 1.2 ± 0.07 (mean ± SEM) days to oviposit, regardless of tube size. The frequency of females that oviposited was dependent on the container size (Fig 9A; χ^2 ^= 11.2, df = 3, P = 0.011). There were pairwise significant differences where oviposition was dependent on tube size, including between 1.5-mL and 15-mL tubes (P = 0.004), and 5-mL and 15-mL tubes (P = 0.017). No other pairwise comparisons were significant. There was no significant difference of the mean number of eggs oviposited among the tube sizes when females did oviposit (Fig 9B; F = 0.20, df = 3, 61, P = 0.893), nor was there a significant difference among the tube sizes in the number of eggs that hatched (Table 2; F = 0.68, df = 3, 61, P = 0.565).
Table 2: Mean ± standard error of the mean (SEM) number and percentage of eggs hatched relative to number tested in four centrifuge tube sizes for lab strains of Culex quinquefasciatus (i.e., CMAVE) and Aedes aegypti (i.e., ORL).
Number of female lab strain Cx. quinquefasciatus that oviposited between 1 and 3 days in Experiment 2, utilizing different tube sizes.No statistical difference was seen among the tube sizes at α = 0.05, so mean ± standard error of the mean (SEM) was calculated across all tube sizes. The mean days to oviposition was 1.2 ± 0.07 (gray column in the figure represents SEM, with the dashed line in the middle representing the mean). All four tube sizes were replicated 8 – 10 times across each of three generational cohorts (N = 114).
Tube size effects on forced oviposition success frequency (A) and mean ± standard error of the mean (SEM) eggs when females oviposited (B) by female lab strain Culex quinquefasciatus.Different lowercase letters above error bars indicate statistical significance of pairwise comparisons between different tube sizes, with significance set at α = 0.05. All four tube sizes were replicated 8 – 10 times across each of three generational cohorts (N = 114).
However, there was a significant difference in the number of eggs that hatched when egg rafts were oviposited broken up rather than intact (Fig 10; F = 5.73, df = 1, 32.74, P = 0.023). Broken up egg rafts were generally in small clusters or singly laid eggs. Of the females that oviposited, 50 had intact egg rafts, with 15 females producing broken up egg rafts.
Mean ± standard error of the mean (SEM) % eggs hatched when lab strain Culex quinquefasciatus rafts were observed broken up versus being a single, intact egg raft.Different lowercase letters above error bars indicate statistical significance of pairwise comparisons of percent egg hatching across all tube sizes, with significance set at α = 0.05. All four tube sizes were replicated 8 – 10 times across each of three generational cohorts (N = 114).
For lab strain Ae. aegypti, all females oviposited within 1 day*.* The frequency of females that oviposited at all was dependent on container size (Fig 11A; χ^2 ^= 8.64, df = 3, P = 0.034). There were significant pairwise comparisons where oviposition was dependent on tube size between 1.5-mL and 5-mL tubes (P = 0.024), 1.5-mL and 15-mL tubes (P = 0.024), and 1.5-mL and 50-mL tubes (P = 0.005). No other pairwise comparisons were significant. When oviposition occurred, there was also a significant effect of tube size on the mean number of eggs oviposited (Fig 11B; F = 5.21, df = 3, 96, P = 0.002). Pairwise comparisons of tube size revealed significant differences between 1.5-mL tubes and 15-mL tubes (P = 0.002), as well as 1.5-mL and 50-mL tubes (P = 0.049). There was no significant difference among the tube sizes in the number of eggs that hatched (Table 2; F = 0.10, df = 3, 96, P = 0.961).
Tube size effects on forced oviposition success frequency (A) and mean ± standard error of the mean (SEM) eggs when females oviposited (B) by gravid female lab strain Aedes aegypti.Different lowercase letters above error bars indicate statistical significance of pairwise comparisons between different tube sizes, with significance set at α = 0.05. All four tube sizes were replicated 15 times across two generational cohorts (N = 120).
Discussion
Our study demonstrated that the modified forced oviposition method is a highly effective approach in generating progeny for both Cx. quinquefasciatus and Ae. aegypti laboratory strains and F_1_ progeny for field-caught individuals. Forced oviposition has been successfully used to establish wild colonies of multiple Anopheles spp. [9–12]. However, this study represents the first report that the technique can be adapted for Ae. aegypti and Cx. quinquefasciatus. Methods developed for Culex spp. and Ae. aegypti have been previously reported in the literature but other techniques used larger containers, provided little instruction, required an excessive number of gravid females to ensure colony success, or recommended more specialized equipment, longer setup, and continual upkeep while holding mosquitoes [17,18]. Our results show that the percentage of females that oviposited was higher in Ae. aegypti compared to Cx. quinqufasciatus, suggesting this method was more effective for Ae. aegypti. That being said, > 50% of Cx. quinquefasciatus lab and field-caught females successfully oviposited (Fig 4A). Despite differences in performance, this technique, while using a minimal number of female mosquitoes and limited equipment, successfully generated numbers of eggs for both species and strains needed for initial colony establishment and subsequent vector research.
In Experiment 1, both laboratory and field-caught Cx. quinquefasciatus females consistently displayed a pattern of longer oviposition times compared to Ae. aegypti. Where all Ae. aegypti strains oviposited within 24 hrs, Cx. quinquefasciatus strains required more time to oviposit, with this pattern dependent on strain displayed by a maximum of 3–7 days to oviposit in laboratory and wild strains, respectively (Fig 4). However, this timeline is not outside the normal observed ranges, as most Culex spp. rearing protocols recommend monitoring for egg rafts up to seven days post blood feeding in laboratory colonies [19,20]. In both Cx. quinquefasciatus and Ae. aegypti, there was no statistical difference in the percentage of laboratory reared and field-caught females that oviposited. All larvae that hatched were reared to the L2 stage at a growth rate commonly observed in both laboratory and field-caught strains of these species [19,20]. Wild gravid Cx. quinquefasciatus females on average laid more eggs or larger egg rafts compared to laboratory strains. Interestingly, the hatch rate of broken egg rafts compared to whole intact egg rafts was significantly reduced in lab strains, while there was no statistical difference in wild Cx. quinquefasciatus hatch rate between broken and intact egg rafts. The higher numbers of oviposited eggs and hatch rate of broken egg rafts in the wild strain may be related to the overall robustness of wild strains compared to inbred lab strains. Laboratory environments themselves impose selective pressures on the population. Standard rearing protocols control all aspects of the mosquito’s environment, which, in this case, may lead to a bottleneck effect that selects for whole rafts or increased egg numbers under strictly ideal conditions [21]. The generational exposure to a standardized environment may lead to the selection of characteristics that would be disadvantageous once placed outside their highly regulated laboratory environment [22,23]. On the other hand, in the wild, it may be an adaptive advantage to produce hardier eggs, that are still viable even if the egg raft breaks. Further research needs to be done to investigate the difference in egg viability and hatch rate between intact and broken Cx. quinquefasciatus egg rafts.
Tube size did affect oviposition rates, with a pattern of reduced oviposition frequency with an increase in container size. Both species appeared to demonstrate a claustrophilic behavior in oviposition (i.e., displaying an oviposition preference and higher frequency of ovipositing in smaller volume tubes by gravid females), with more females laying in 1.5 and 5 mL tubes for Cx. quinquefasciatus and 1.5 mL tubes for Ae. aegypti. Additionally, the mean number of eggs laid by Ae. aegypti decreased as tube size increased. There was no change in mean egg numbers for Cx. quinquefasciatus across all tube sizes. The change in egg numbers across tube size seen in Ae. aegypti could be attributed to their ‘skip-oviposition’ behavior, where females distribute eggs across multiple oviposition sites within a single gonotrophic cycle. This means eggs are laid intermittently and in many cases Ae. aegypti females only aggregate a small percentage of their eggs in a single container [24–26]. By comparison, Cx. quinquefasciatus typically lay their eggs as a clutch in a single raft, which could be interpreted as an ‘all or nothing’ approach [27]. To our knowledge, there are no reports of an intermittent style oviposition behavior observed in Cx. quinquefasciatus. One limitation in the present study for the claim that mosquitoes exhibit claustrophilic oviposition is that tubes were not selected based on a consistent ratio of increasing size. Instead, tube sizes were selected based on their similar material (i.e, plastic) and common availability in laboratories and mosquito control districts. Thus, with the data generated in the current study, we cannot rule out that tube dimensions may have played a role in oviposition behaviors. Further study, using containers proportionally scaled in size, would strengthen the definition of claustrophilic oviposition.
Based on our data, we recommend the use of 1.5 mL tubes for Cx. quinquefasciatus and Ae. aegypti egg collection, regardless of the strains being laboratory reared, or field caught. Tube setup should follow the methods outlined in Experiment 1, with the key difference between the two species being the level of excess water left in the tubes (a thin layer of water above the cotton, approx. 1 mm at the meniscus for Cx. quinquefasciatus, and complete removal of excess water for Ae. aegypti). A key consideration for adopting this method is the quality and type of water that is available. While tap water was used in this study, if a particular water source is deemed unacceptable for rearing purposes, then alternative waters sources may be necessary. Additionally, we recommend leaving Cx. quinquefasciatus females to oviposit for 3–4 days for lab strains and up to 7 days for wild strains. Aedes aegypti only required 24 hrs post transfer before they oviposited. It is worth noting that the age and degree of blood engorgement of laboratory and field-caught mosquitoes was not controlled for, nor was the longevity of eggs examined in this study. Adult age and level of blood engorgement are key factors that influence oviposition, however, it is difficult to control for these in field caught populations [27]. Further study will need to be conducted to determine if age and blood engorgement significantly impact the success of this forced oviposition method. All Ae. aegypti egg papers were hatched within 48 hrs, and Cx. quinquefasciatus egg rafts were not held in oviposition tubes past 24 hrs. For the purposes of this paper, we recommend readers adhere to the same timeline for egg hatching, i.e., Ae. aegypti papers dried for 48 hrs before hatching and Cx. quinquefasciatus egg rafts placed in water within 24 hrs.
The present study demonstrates that the forced oviposition method can be adapted to other mosquito vector species besides Anopheles spp. Present challenges in mosquito vector research start initially with ensuring teams have access to sufficient numbers of mosquitoes and successful production of F_1_ progeny required for colony establishment of field strains. This technique was shown to be highly effective in generating viable eggs using a minimal number of female mosquitoes and readily available items found in most laboratories and mosquito control districts. Proper utilization of the forced oviposition method expands our ability to conduct a variety of vector research with multiple key mosquito vector species, such as single female sex ratio determination, genetic isoline production by controlling for specific genetic backgrounds, and heritability studies focused on selective mating.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Franklinos LHV, Jones KE, Redding DW, Abubakar I. The effect of global change on mosquito-borne disease. Lancet Infect Dis. 2019;19(9):e 302–12. doi: 10.1016/S 1473-3099(19)30161-6 31227327 · doi ↗ · pubmed ↗
- 2WHO. Vector-borne diseases [Internet]. World Health Oganization. 2024 [cited 2025 Mar 15]. https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases
- 3Venkatesan P. WHO world malaria report 2024. Lancet Microbe. 2025;101073.10.1016/j.lanmic.2025.10107339923782 · doi ↗ · pubmed ↗
- 4WHO. Dengue - Global situation [Internet]. World Health Oganization. 2024 [cited 2025 Mar 10]. https://www.who.int/emergencies/disease-outbreak-news/item/2024-DON 518
- 5Idris F, Ting DHR, Alonso S. An update on dengue vaccine development, challenges, and future perspectives. Expert Opin Drug Discov. 2021;16(1):47–58. doi: 10.1080/17460441.2020.1811675 32838577 · doi ↗ · pubmed ↗
- 6Karunamoorthi K. Vector control: a cornerstone in the malaria elimination campaign. Clin Microbiol Infect. 2011;17(11):1608–16. doi: 10.1111/j.1469-0691.2011.03664.x 21996100 · doi ↗ · pubmed ↗
- 7Wilson AL, Courtenay O, Kelly-Hope LA, Scott TW, Takken W, Torr SJ, et al. The importance of vector control for the control and elimination of vector-borne diseases. P Lo S Negl Trop Dis. 2020;14(1):e 0007831. doi: 10.1371/journal.pntd.0007831 31945061 PMC 6964823 · doi ↗ · pubmed ↗
- 8Masters SW, Knapek KJ, Kendall LV. Rearing Aedes aegypti Mosquitoes in a Laboratory Setting. 2020.PMC 758528333106772 · pubmed ↗