A Transcriptional Variant of Anaplastic Lymphoma Kinase Promotes Apoptosis in Ovarian High‐Grade Serous Carcinoma
Ako Yokoi, Daigo Yoshimori, Yasuko Oguri, Miki Hashimura, Makoto Saegusa

TL;DR
A new variant of the ALK gene promotes cell death in ovarian cancer cells, possibly through interaction with a DNA repair protein.
Contribution
The study identifies a novel ALK transcript, ALKATI, and its role in inducing apoptosis in ovarian high-grade serous carcinoma.
Findings
ALKATI localizes to both the cytoplasm and nucleus in HGSC cells, unlike full-length ALK.
ALKATI increases sensitivity to cisplatin-induced apoptosis and interacts with PARP1.
ALKATI reduces epithelial-mesenchymal transition and cancer stem cell properties without affecting proliferation.
Abstract
The current study aims to delineate the role of a novel anaplastic lymphoma kinase (ALK) transcript, ALKATI, in ovarian high‐grade serous carcinoma (HGSC). Overexpressed ALKATI exhibited both cytoplasmic and nuclear localization in HGSC cells, whereas full‐length ALK was predominantly cytoplasmic. ALKATI interacts with the DNA repair protein, poly (ADP ribose) polymer 1 (PARP1), and cells stably overexpressing ALKATI (OE‐ ALKATI) were more sensitive to cisplatin‐induced apoptosis. Consistent with this, cleaved PARP1 levels were higher in HGSC tissue samples in areas with nuclear ALK immunoreactivity. The ratio of antiapoptotic BCL2 relative to proapoptotic BAX was significantly increased in OE‐ALKATI cells, despite the increase in apoptosis, suggesting that ALKATI‐mediated apoptosis is independent of mitochondrion‐driven cell death. OE‐ALKATI decreased epithelial‐mesenchymal…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
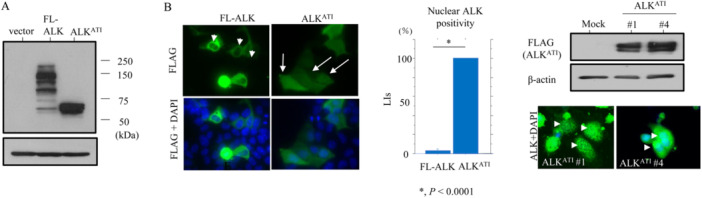 Figure 1
Figure 1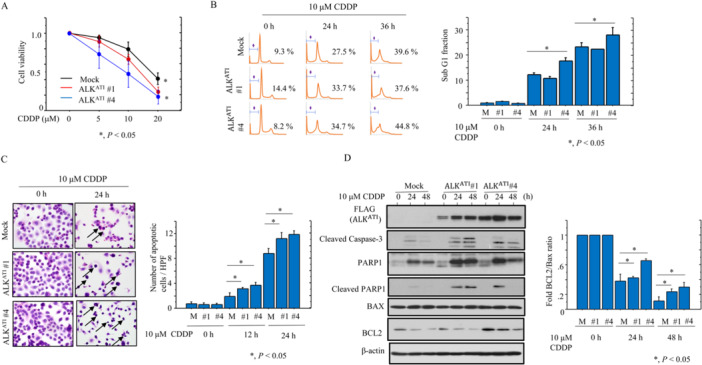 Figure 2
Figure 2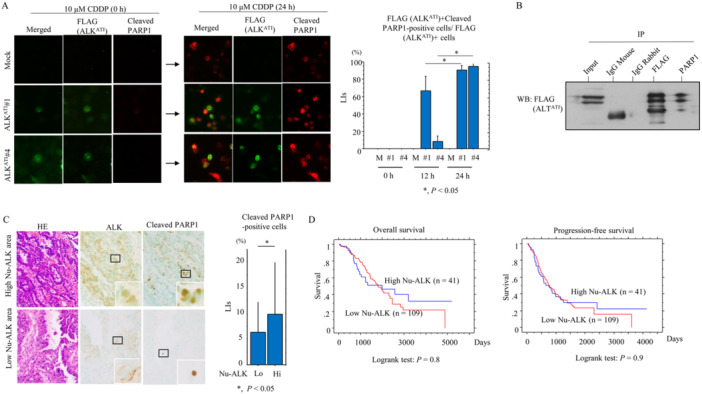 Figure 3
Figure 3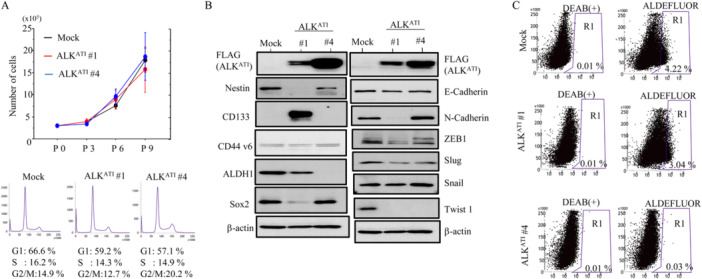 Figure 4
Figure 4- —This study was supported by a grant from JSPS KAKENHI (Grants 23K06471 [MS] and 23K14484 [AY].10.13039/501100001691
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPARP inhibition in cancer therapy · Ovarian cancer diagnosis and treatment · DNA Repair Mechanisms
Introduction
1
Ovarian carcinoma is the leading cause of mortality among all gynecological malignancies; this can be attributed to the presence of peritoneal metastases in 74% of cases at diagnosis [1, 2]. High‐grade serous carcinoma (HGSC) is by far the dominant subtype diagnosed clinically, and accounts for up to 80% of all ovarian cancer deaths [3, 4]. Delineating the molecular mechanisms involved in the initiation and progression of HGSC is therefore critical for the discovery of novel therapeutic agents to improve survival rates.
Anaplastic lymphoma kinase (ALK) is a 200‐kDa receptor tyrosine kinase encoded by the ALK gene on chromosome 2p23. The ALK protein consists of a large extracellular domain, a lipophilic transmembrane segment, and a cytoplasmic tyrosine kinase domain [5, 6]. Consistent with its role as an oncogenic driver, full‐length ALK and ALK translocations are found in primary solid tumors including neuroblastoma and lung carcinoma [7, 8].
An alternative transcription initiation (ATI) site located in intron 19 of the ALK gene leads to the expression of ALK isoform, ALK^ATI^, which encodes only the intracellular kinase domain [9, 10]. ALK^ATI^ is expressed in approximately 11% of melanoma cases, less frequency in other tumor types and is not present in normal tissues [9, 10].
We have shown that full‐length ALK is a critical driver of HGSC and engenders the aggressive phenotypic characteristics of HGSC [11]. Spurred by the discovery of ALK^ATI^ in other tumor types, we designed the current study to evaluate the role of this novel isoform in HGSC.
Materials and Methods
2
Plasmids and Cell Lines
2.1
ALK^ATI^ cDNA was amplified from a human full‐length ALK cDNA clone [11] by PCR using the specific primers, 5′‐ GCGAATTCATGCAGATGGAGCTGCAGAGCCC‐3′ and 5′‐ CGGGATCCGGGCCCAGGCTGGTTCATGC‐3′. The resulting PCR products, as well as full‐length ALK cDNA, were cloned into the p3xFLAG‐CMV‐14 vector (Sigma‐Aldrich Chemicals, St Louis, MO). Clones stably overexpressing ALK^ATI^ were established using the HGSC cell line, OVCAR‐3, as described previously [11].
Antibodies and Reagents
2.2
Antibodies are listed in Supplementary Table S1. Cisplatin (CDDP; P4394) and doxorubicin (Dox; D1515) were obtained from Sigma‐Aldrich Chemicals (St. Louis, MO, USA).
Western Blot and Co‐Immunoprecipitation (Co‐IP) Assays
2.3
Extraction of total cellular proteins, western blot, and co‐IP assays were carried out as described previously [11].
Flow Cytometry and Aldefluor Assay
2.4
Cell cycle analysis and ALDH1 enzyme activity using a fluorogenic dye‐based Aldefluor assay (Stem Cell Technologies, Grenoble, France) were performed as described previously [11].
Cell Counting Kit‐8 Assay
2.5
Cell viability after CDDP treatment was evaluated using the Cell Counting Kit‐8 (CCK‐8; Dojindo Lab, Kumamoto, Japan), according to the manufacturer's instructions.
Apoptotic Index
2.6
The number of apoptotic cells identified in HE‐stained sections was calculated by counting the mean number of apoptotic figures per field as described previously [12].
Immunofluorescence
2.7
After 10 μM CDDP treatment, cells were incubated with primary antibodies. Alexa 488 and 570 (Thermo Fisher Scientific, Waltham, MA, USA) were used as secondary antibodies. Cells that were double‐immunopositive for FLAG‐ALK^ATI^ and cleaved PARP1 were counted in five randomly selected high‐power fields (HPFs). The labeling indices (LIs) were then calculated per the number of FLAG‐ALK^ATI^ immunopositive cells.
Clinical Cases
2.8
We selected 150 cases of surgically resected HGSC (Supplementary Table S2) that were carried out between 2005 and 2019 at Kitasato University Hospital. Cases were chosen, according to the criteria of the 2014 World Health Organization classification, the TNM, and the International Federation of Gynecology and Obstetrics (FIGO) classification [13, 14, 15]. Thirty‐three cases received neoadjuvant paclitaxel/carboplatin‐based chemotherapy (NAC) before surgical treatment (Supplementary Table S2). NAC‐treated HGSC cases were subdivided into three groups on the basis of chemotherapy response score (CRS) for HGSC [16], as follows: CRS 1, no or minimal tumor response; CRS 2, partial response; CRS3, total or near‐total response. All tissues were routinely fixed in 10% formalin and processed for embedding in paraffin wax. Approval for this study was given by the Ethics Committee of the Kitasato University School of Medicine (B18‐048).
Immunohistochemistry (IHC)
2.9
IHC was performed using a combination of the microwave oven heating and polymer immunocomplex (Envision, Dako) methods as described previously [11, 12]. For evaluation of IHC findings, scoring of nuclear, cytoplasmic, or membranous immunoreactivities was performed [11, 12]. Briefly, the proportion of immunopositive cells among the total number of counted cells was subdivided into five categories as follows: 0, all negative; 1, < 10%; 2, 10–30%; 3, 30–50%; and 4, > 50% positive cells. The immunointensity was also subclassificed into four groups: 0, negative; 1, weak; 2, moderate; and 3, strong. IHC scores were generated by multiplication of the values of the two parameters. We subdivided the nuclear ALK scores into high and low categories by cutoff value (= 3, as well as 1, 2, 4, 6, and 8) on the basis of the mean value (= 2.3) (Supplementary Figure S1). Cleaved PARP1‐immunopositive cells were also counted in five HPFs that were randomly selected from nuclear ALK‐high and ALK‐low areas, respectively. We then calculated cleaved PARP1 LIs in these regions. In addition, p53 immunoreactivity was subdivided into mutant (mt, score 0 and 6‐12) and wild‐type (wt, score 1‐5) classes [17, 18].
Statistics
2.10
Comparative data were analyzed using the Mann–Whitney U‐test and Chi‐square test. Overall survival (OS) and progression‐free survival (PFS) were calculated as described previously [11]. The cutoff for statistical significance was set as p < 0.05.
Results
3
Transcripts from the ATI site in intron 19 of the ALK gene result in three isoforms that encode proteins with predicted molecular weights of 61.1, 60.8, and 58 kDa [9, 10]. Consistent with this, we observed a major band of 60 kDa following transfection of ALK^ATI^ into OVCAR‐3 cells, whereas full‐length ALK migrated at around 200 kDa (Figure 1A). ALK^ATI^ exhibited both nuclear and cytoplasmic localization both when transiently overexpressed and when stably overexpressed in OVCAR‐3 cells (OE‐ALK^ATI^). In contrast, full‐length ALK was predominantly cytoplasmic (Figure 1B). OE‐ALK^ATI^ cells were significantly more sensitive to CDDP‐induced apoptosis when compared to mock cells (Figure 2A). Consistent with this, we observed an increased sub‐G1 fraction (Figure 2B) and more apoptotic cells (Figure 2C), as well as increased expression of ALK^ATI^, cleaved caspase‐3, PARP1, cleavedPARP1, and a higher BCL2: BAX ratio (Figure 2D). Similar findings were also observed in the Dox‐treated OE‐ALK^ATI^ cells (Supplementary Figure S2A). Co‐immunolocalization of nuclear ALK^ATI^ and cleaved PARP1 was significantly higher in OE‐ALK^ATI^ cells at 24 h post‐CDDP treatment when compared to 12 h (Figure 3A). This was consistent with the increased interaction between ALK^ATI^ and PARP1 observed following co‐IP experiments (Figure 3B). Heterogenous high nuclear ALK immunoreactivity (IHC score ≧ 3) was observed in 40 (26.6%) of 150 clinical HGSC tissue samples (Figure 3C). Of 33 cases (CRS1, 13 cases; CRS2, 20; CRS3, 0) receiving NAC, nuclear ALK scores appeared higher in patients with CRS 2 as compared to those of CRC1, although the difference was not statistically significant, probably due to the small number of cases investigated (Supplementary Figure S2B). Moreover, cleaved PARP1 immunoreactivity was significantly higher in ALK‐high nuclear immunopositive lesions when compared to the ALK‐low regions (Figure 3C). In contrast, nuclear ALK scores did not correlate with clinicopathological factors, OS, or PFS in HGSC, even when subdivisions were further made on the basis of five cutoff values (Figure 3D and Supplementary Table S2).
Both nuclear and cytoplasmic localization of ALKATI in HGSC cells. (A) Western blot analysis for the indicated proteins in total lysates from OVCAR‐3 cells transfected with empty vector, full‐length (FL) ALK, or ALKATI. (B) Left: immunofluorescence of OVCAR‐3 cells transfected with full‐length (FL) ALK or ALKATI. Note the lack of nuclear FL‐ALK staining (indicated by arrowheads), in contrast to distinct nuclear ALKATI staining (indicated by arrows). FL‐ALK and ALKATI were detected using anti‐FLAG antibody. Middle: the LIs of nuclear immunopositivity for FL‐ALK and ALKATI are shown as mean ± SD. Upper right: western blot analysis for the indicated proteins in total cell lysates from OE‐ALKATI and mock cells. Lower right: immunofluorescence of OE‐ALKATI cells. Note the distinct nuclear ALKATI staining (indicated by arrow‐heads).
Changes in apoptosis following ALKATI overexpression in HGSC cells. (A) Treatment of OE‐ALKATI and mock cells with 10 μM CDDP for 24 h. Cell viability was measured using the CCK‐8 Viability kit. Viability for untreated cells is set as 1. (B) Left: flow cytometric cell cycle analysis for OE‐ALKATI and mock cells (M) after 10 μM CDDP treatment for the time shown. Daggers indicate sub‐G1 fractions. Right: the percentage of cells undergoing apoptosis (sub‐G1) was calculated and the data shown are as mean ± SD. The value in untreated cells is set as 1. (C) Left: arrows show OE‐ALKATI and mock cells (M) undergoing apoptosis after 10 μM CDDP treatment. Original magnification, x400. Right: numbers of apoptotic cells are shown as mean ± SD. (D) Left: western blot analysis of the indicated proteins in total lysates from OE‐ALKATI and mock cells with 10 μM CDDP treatment for the time shown. Right: the BCL2: BAX ratio was calculated following normalization to β‐actin using ImageJ version 1.41 (NIH, Bethesda, MD; http//imagej.gov/ij) and the data are shown as mean ± SD. Expression levels in untreated cells (0 h) were set as 1.
Relationship between ALKATI and PARP1 in HGSC. (A) Left: immunofluorescence of ALKATI and cleaved PARP1 in OE‐ALKATI and mock cells treated with 10 μM CDDP for the indicated times. Note the co‐immunolocalization of ALKATI and cleaved PARP1 in nuclei of cells treated with CDDP. Immunoreactivity for ALKATI is detected using anti‐FLAG antibody. Right: percentages of ALKATI‐positive cells that are also positive for cleaved PARP1 are shown as mean ± SD. (B) After immunoprecipitation (IP) with the indicated antibodies using OE‐ALKATI cell lysates, we performed western blot (WB) using anti‐FLAG antibody. Input was 5% of the total cell extract. Normal rabbit and mouse IgGs were used as a negative control. (C) Left: staining with HE and IHC for the indicated proteins in samples with high and low nuclear (Nu)‐ALKATI‐immunopositive (upper and lower panels, respectively). The closed boxes are magnified in the insets. Original magnification, x200 and x400 (insets). Right: cleaved PARP1 LIs in the high and low (Hi and Lo) Nu‐ALKATI ‐immunopositive categories. The LIs shown are mean ± SD. (D) OS (right) and PFS (left) relative to nuclear (Nu)‐ALKATI (low vs. high expression) in HGSC. n, number of cases.
Since data from the TCGA Research Network indicate that HGSC is characterized by TP53 mutations in up to 96% of the cases [19] and TP53 wt HGSC are rare (< 1%) [20], we further examined whether p53 abnormality affects nuclear ALK immunoreactivity in HGSC. Although mt and wt p53 were found in 47 (81%) and 11 (18.9%) of 58 HGSC cases investigated, respectively, there was no association between nuclear ALK scores and p53 status (Supplementary Figure S3); this may be due to extremely high p53 abnormality in HGSC, in contrast to the nuclear ALK immunopositivity.
OE‐ALK^ATI^ cells had similar proliferative rates and cell cycle profiles as mock cells (Figure 4A). In contrast, levels of several epithelial‐mesenchymal transition (EMT)‐ and cancer stem cell (CSC)‐related markers including Nestin, ALDH1, ZEB1, Slug, and Twist 1 were lower in OE‐ALK^ATI^ cells when compared to mock cells (Figure 4B). Consistent with a lower level of CSC‐like properties, the frequency of ALDH1^high^ cells was low in the OE‐ALK^ATI^ cell population compared to mock cells (Figure 4C). In contrast, there were no associations between nuclear ALK scores and expression of several CSC markers in HGSC (Supplementary Figure S4); this may be due to the heterogenous distribution of nuclear ALK‐immunopositive cells within the tumors.
Relationships between ALKATI, proliferation, and epithelial‐mesenchymal transition/cancer stem cell properties in HGSC cells. (A) Upper: OE‐ALKATI and mock cells were seeded at low density. P0, P3, P6, and P9 are 0, 3, 6, and 9 days after seeding, respectively. Lower: flow cytometry analysis of OE‐ALKATI and mock cells 3 days after seeding (P3). (B) Western blot analysis for the indicated proteins in total lysates from OE‐ALKATI and mock cells. (C) The percentage of live single‐cell populations, as well as the values of R1 in Aldefluor relative to R1 in DEAB(+) (indicated by parentheses), contained in each gate in OE‐ALKATI and mock cells are shown.
Discussion and Conclusions
4
We found that overexpression of ALK^ATI^ sensitizes HGSC cells to CDDP and is associated with the response to CDDP in clinical HGSC samples. Based on the colocalization and physical interaction of ALK^ATI^ and PARP1, we speculate that ALK^ATI^ may perturb PARP1‐dependent DNA repair pathways that are activated by genotoxins such as CDDP [21]. This would be consistent with the known regulation of DNA damage response, gene expression, and nuclear action structure formation by other tyrosine kinases [22, 23]. Since the BCL2:BAX ratio actually increases during CDDP‐ and Dox‐induced apoptosis in OE‐ALK^ATI^ cells, we suggest that ALK^ATI^‐mediated apoptosis is independent of ‘classical’ mitochondrion‐driven cell death.
There are conflicting results regarding the functional role of ALK^ATI^ during tumor progression. For example, ALK^ATI^ reportedly stimulates multiple oncogenic signaling pathways, mediates growth‐factor‐independent cell proliferation and promotes tumorigenesis [9]. ALK^ATI^ also acts as a potential prognostic factor and therapeutic target in soft tissue sarcoma [24]. In contrast, others have found that ALK^ATI^ expression is insufficient for cellular transformation or growth, and does not predict single agent therapeutic activity in melanoma cells [25]. In our results, cells overexpressing ALK^ATI^ had a similar proliferative rate to mock cells, but did display reduced EMT/CSC properties, although OE‐full‐length ALK cells enhanced cancer stem cell (CSC) features, increased cell proliferation, and accelerated cell mobility [11]. Moreover, nuclear ALK‐immunopositivity correlated neither with clinicopathological factors nor with prognosis in HGSC. Together, these data lead us to conclude that while ALK^ATI^ expression may sensitize HGSC cells to genotoxin‐induced apoptosis, it may play a relatively minor overall role in tumor progression.
Author Contributions
Ako Yokoi, RO, and Makoto Saegusa carried out the majority of the experiments, analyzed the data, and wrote the manuscript. They were helped by Yasuko Oguri and Miki Hashimura. All authors reviewed and approved the final manuscript.
Ethics Statement
This study was approved by the Kitasato University Medical Ethics Committee (B20‐181).
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Sup Figure S1.
Sup Figure S2.
Sup Figure S3 v2.
Sup Figure S4.
Supplementary Table S1.
Supplementary Table S2.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1A. Jemal , R. Siegel , E. Ward , T. Murray , J. Xu , and M. J. Thun , “Cancer Statistics, 2007,” CA: A Cancer Journal for Clinicians 57 (2007): 43–66.17237035 10.3322/canjclin.57.1.43 · doi ↗ · pubmed ↗
- 2Y. Wang , Å. Helland , R. Holm , et al., “TP 53 Mutations in Early‐Stage Ovarian Carcinoma, Relation to Long‐Term Survival,” British Journal of Cancer 90 (2004): 678–685.14760384 10.1038/sj.bjc.6601537 PMC 2410156 · doi ↗ · pubmed ↗
- 3R. J. Kurman and I. M. Shih , “The Dualistic Model of Ovarian Carcinogenesis,” American Journal of Pathology 186 (2016): 733–747.27012190 10.1016/j.ajpath.2015.11.011PMC 5808151 · doi ↗ · pubmed ↗
- 4D. D. Bowtell , S. Böhm , A. A. Ahmed , et al., “Rethinking Ovarian Cancer II: Reducing Mortality From High‐Grade Serous Ovarian Cancer,” Nature Reviews Cancer 15 (2015): 668–679.26493647 10.1038/nrc 4019 PMC 4892184 · doi ↗ · pubmed ↗
- 5T. Iwahara , J. Fujimoto , D. Wen , et al., “Molecular Characterization of ALK, a Receptor Tyrosine Kinase Expressed Specifically in the Nervous System,” Oncogene 14 (1997): 439–449.9053841 10.1038/sj.onc.1200849 · doi ↗ · pubmed ↗
- 6S. W. Morris , C. Naeve , P. Mathew , et al., “ALK, the Chromosome 2 Gene Locus Altered by the t(2;5) in Non‐Hodgkin's Lymphoma, Encodes a Novel Neural Receptor Tyrosine Kinase That Is Highly Related to Leukocyte Tyrosine Kinase (LTK),” Oncogene 14 (1997): 2175–2188.9174053 10.1038/sj.onc.1201062 · doi ↗ · pubmed ↗
- 7L. Lamant , K. Pulford , D. Bischof , et al., “Expression of the ALK Tyrosine Kinase Gene in Neuroblastoma,” American Journal of Pathology 156 (2000): 1711–1721.10793082 10.1016/S 0002-9440(10)65042-0PMC 1876934 · doi ↗ · pubmed ↗
- 8F. Guo , X. Liu , Q. Qing , et al., “EML 4‐ALK Induces Epithelial–Mesenchymal Transition Consistent With Cancer Stem Cell Properties in H 1299 Non‐Small Cell Lung Cancer Cells,” Biochemical and Biophysical Research Communications 459 (2015): 398–404.25735977 10.1016/j.bbrc.2015.02.114 · doi ↗ · pubmed ↗