The Drosophila FM7 - A Strain Contains a Fixed Translocation Involving the X and 4 Chromosomes
Keith Maggert, Selina Kindelay

TL;DR
This study reveals that the Drosophila FM7-A balancer chromosome is a translocation between chromosomes 1 and 4, which affects genetic experiments.
Contribution
The discovery of a new translocation structure in the FM7-A balancer chromosome provides insights into its genetic behavior.
Findings
FM7-A is a translocation involving chromosomes 1 and 4 with 5% of chromosome 1 material transferred.
The chromosome 4 segregant is viable and fertile without the chromosome 1 segregant.
The translocation structure may impact genetic experiments using this balancer chromosome.
Abstract
We discovered that a commonly used balancer chromosome – FM7-A, Tb 1 – exists as an uncharacterized translocation between the progenitor FM7a chromosome and a normal chromosome 4 . Genetic and cytological data evidence show that T ( 1 ; 4 ) FM7-A translocates about 5% of chromosome 1 material to an entire chromosome 4 , thus the large “chromosome 1 ” segregant is lethal without the small “chromosome 4 ” segregant, though the latter is viable and fertile without the former. We characterize this balancer chromosome to inform Drosophila molecular-geneticists of the structure as it may affect their use of this balancer chromosome and the interpretations of their results.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
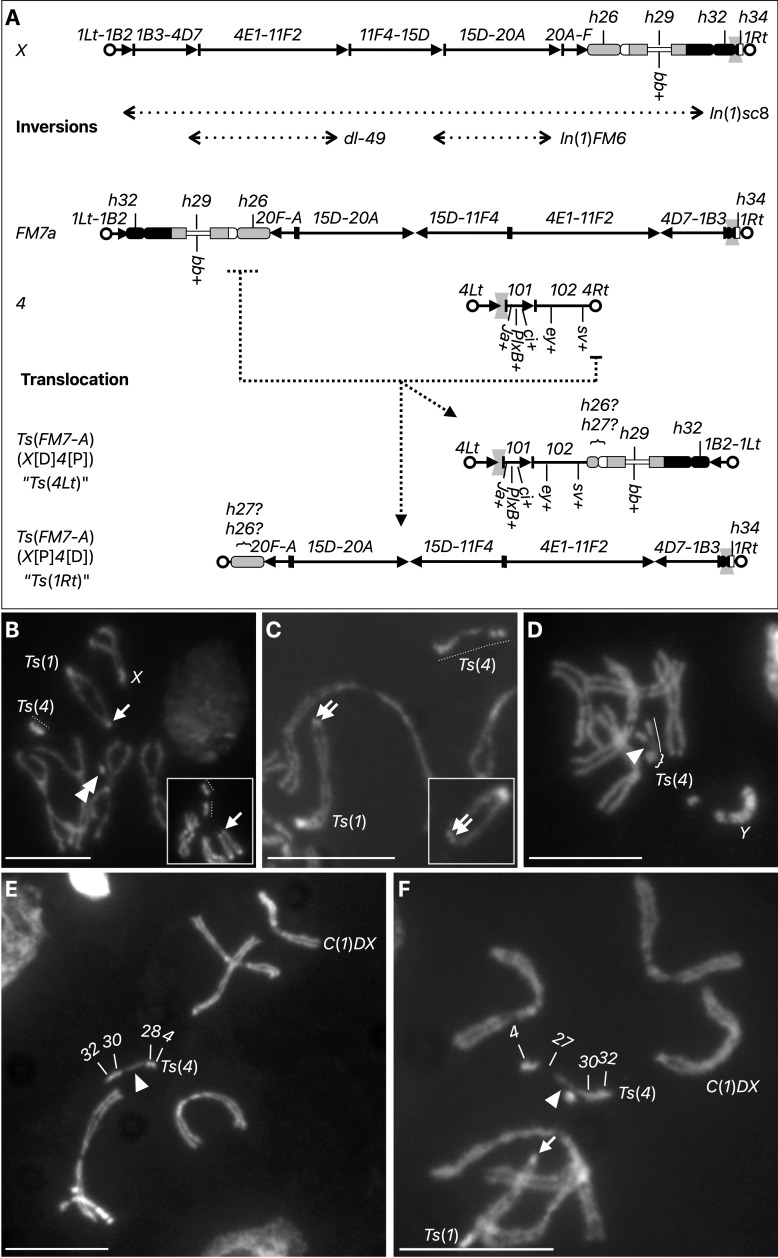 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomic variations and chromosomal abnormalities · Developmental Biology and Gene Regulation · Microtubule and mitosis dynamics
Description
In the course of examining the tempo and conditions of rDNA magnification (Kindelay and Maggert 2024), we found that the * FM7-A * chromosome strain de-linked the complementation of a lethal null allele of the rDNA ( * bb * , the bobbed locus) from the X chromosome (Gatti and Pimpinelli, 1992), leading us to discover an unknown and uncharacterized chromosome translocation between chromosome 1 (the “ * FM7-A * ” X ) and 4 in this strain. * FM7-A * was generated by introduction of a P -element containing dominant * white * ^+^ and * Tubby * ^1^ alleles to the existing X -chromosome * FM7a * balancer chromosome (Merriam, 1969) by Ramono Lattao and colleagues (Lattao et al., 2011), and is currently available from the Bloomington Drosophila Stock Center (as * FM7a * , P { * w * ^+mC^
Tb * ^1^ } * FM7-A * ) (RRID:BDSC_36489). As we describe below, we believe this translocated chromosome set has breakpoints in h26 or h27 (of the original * FM7a * ) and distal 102 (of the original chromosome 4 ) ( Figure 1A ). Any such translocation is composed of two elements which can (in principle, though not necessarily in practice) be separated through independent segregation in meiosis: here the complete genotype ( T ( 1 ; 4 ) * FM7-A * , * FM7a * : 4 ) has a large translocation segregant ( Ts ( 1 ^P^ 4 ^D^ ) * FM7-A * , henceforth in this work, “ Ts ( 1 )”) and a small segregant ( Ts ( 1 ^D^ 4 ^P^ ) * FM7-A * , henceforth “ Ts ( 4 )”). Ts ( 1 ) possesses most of the X euchromatin and is driven by the chromosome X centromere ( 4Rt
102 || [ h26
h27 ] | 20F
20A | 15D
20A | 15D
11F4 | 4E1 - 11F2 | 4D7
1B3 | h32 • 1Rt — the brackets around h26
h27 in Figure 1A indicate ambiguity in the breakpoint, and the double vertical line is the “ * FM7-A * breakpoint” defined by the translocation described herein). Ts ( 4 ) possesses most of chromosome 4 driven by the 4 centromere, and is appended by the tip of the X ( 4Lt • 101
*102 * || [ h27
h32 ] | 1B2
1Lt ).
To map the breakpoints, we performed genetic crosses to ascertain if linkage of genes moved between * FM7a * and chromosome 4 in the * FM7-A * strain. We found that y w / Y ; Ts ( 4 ) males heterozygous for three alleles of * JY _ alpha _ * ** ** ( * JY alpha * ^CR70483-TG4.0^ , * JY alpha * ^I^ , and * JY alpha * ^P^ ) were fertile, indicating the breakpoint does not affect, and is distal to, * JY _ alpha _ * , the proximal-most mutable locus on chromosome 4 (Öztürk-Çolak et al., 2024). y w / Y ; Ts ( 4 ) males were viable when heterozygous with Df ( 4 ) * M101-63a * , Df ( 4 ) * M101-62f * , or two alleles of * Plexin B * ( PlexB ^KG00878^ and PlexB ^MI15559-DH.GT-TG4.1^ ), indicating that the chromosome *4 * breakpoint does not affect these other genes in the proximal region, and is therefore in distal 4 . Correspondingly, females and males of genotype y w ; Ts ( 4 )/ ci gvl ey sv were phenotypically normal for all four chromosome 4 -linked mutations, indicating that the chromosome 4 -linked breakpoint is distal to * cubitus interruptus * in cytological band * 101E/F, eyeless * in proximal 102 , and * shaven * in distal 102. Finally, we were able to generate y w ; Ts ( 4 )/ Ts ( 4 ) females and males and maintain them as a homozygous stock, indicating that the chromosome 4 breakpoint is distal to any lethal or fertile complementation groups, including the distal-most gene identified on the right arm of chromosome 4 (the gene Calcium-dependent secretion activator , * Cadps * , which is homozygous lethal (Renden et al. 2001)). We conclude from these data that the chromosome 4 breakpoint is between * Cadps * and the telomere, and does not involve the movement of any genes from chromosome 4 to the Ts ( 1 ) element of the translocation.
Ts ( 4 ) dominantly ( i.e. , as a heterozygote) complemented strongly-expressive * bobbed * mutations induced on two separate Y chromosomes ( Y , *484 * and Y , 183 ) in C ( 1 ) DX / Y ; Ts ( 4 )/ 4 females (Paredes and Maggert 2009), which indicates that the X -linked breakpoint is proximal to the * bobbed * locus in h29 as it was inverted as part of the h26
h32 block to distal X in the progenitor * FM7a * ( Figure 1A ).
In diploid neuroblasts, Ts ( 4 ) appeared much larger than a normal chromosome 4 ( Figure 1B, dotted line and double arrowhead, respectively), indicating an acquisition of a significant amount of genomic material. Correspondingly, Ts ( 1 ) appeared truncated relative to the regular X chromosome, and notably ended with a small dot of heterochromatin ( Figure 1B, arrow, see also inset). We infer this block of heterochromatin to be h26 and perhaps up to h29 , the location of the rDNA (Gatti and Pimpinelli, 1992). First, the staining with DAPI is brighter, indicating at least some heterochromatin is present. Second, the bright ends of the chromosome exhibit sister cohesion even when clearly euchromatic regions do not. Finally, the loss of the ability of the Ts ( 1 ) element to rescue * bobbed * alleles (while the Ts ( 4 ) acquired it) means that little if any of h29 moved to chromosome 4 . We interpret the small size of this distal cohesing heterochromatin as h26 or h26 / h27 . This means that the Ts ( 4 ) element contains h26 / h27 through h32 , plus *1B2 *
1Lt , accounting for the larger-than-normal appearance plus the new heterochromatin/euchromatin of Ts ( 4 ). To better resolve the heterochromatic breakpoint, we altered the rDNA dose to alter the condensation of the rDNA .
The Ts ( 4 ) element appears condensed in Ts ( 1 )/ X ; Ts ( 4 )/ 4 females ( Figure 1B ), indicating relatively inactive rDNA . The rDNA were more active in Ts ( 1 )/ Y ; Ts ( 4 )/ 4 males ( Figure 1C ). The pronounced activity of the rDNA appears as a decondensed rDNA chromosomal region and the chromosome 4
- and
FM7a * -derived materials are clearly discriminable (bracket and line, respectively). This is the general trend we've described before: males show more biased X -linked rDNA expression than do females (Kindelay and Maggert, 2024). In C ( 1 ) DX / Y ; Ts ( 4 )/ 4 females, which have more Ts ( 4 )-linked rDNA expression, the decondensed rDNA locus was evident ( Figure 1D, arrowhead). The rDNA was extremely decondensed when the rDNA were removed from the Y in C ( 1 ) DX / Y , rDNA ^-^ ; Ts ( 4 )/ 4 females ( Figure 1E, arrowhead). In C ( 1 ) DX / Ts ( 1 ); Ts ( 4 )/ 4 metafemales, though the Ts ( 4 ) element shows pronounced decondensation at the rDNA , there is no sign of decondensation on the Ts ( 1 ) element ( Figure 1F, arrowhead and arrow, respectively). These data indicate that the entirety of the rDNA , which retains normal activity and regulatory properties, were transferred to Ts ( 4 ). Further, the enhanced resolution clearly shows the heterochromatic bands and euchromatic *1B2 *
1Lt sequence on the Ts ( 4 ) element (as indicated).
We could not generate Ts ( 1 )/ Y ; ci gvl ey sv males, confirming that Ts ( 4 ) contains genes that were originally X -linked that are necessary for viability. It is likely these genes correspond to cytological region 1Lt - 1B2 ( Figure 1A ), which is distalmost in * FM7a * , and contains approximately 25-30 genes (Öztürk-Çolak et al., 2024). First, the Y in males of this genotype supported viability of C ( 1 ) DX / Y females, indicating that it is not the translocation of the bobbed locus away from * FM7-A * that is responsible for the lethality in Ts ( 1 )/ Y ; +/+ males. Second, we reasoned that if our inability to recover Ts ( 1 )/ Y ; +/+ flies was due to loss of the now Ts ( 4 )-linked X genes, we could rescue the lethality by reintroduction of 1Lt
- 1B2* from a different source. We therefore sought to create Ts ( 1 )/ Y ; +/+ flies bearing the free duplication
Dp(1;f)1187 * , which shares the scute-8 breakpoint with * FM7a * and * FM7-A * (Sun et al. 2003). Addition of this chromosome restored viability, demonstrating that the euchromatin distal to the distal heterochromatic block on * FM7a * is missing in Ts ( 1 )/ Y ; +/+ males, and its absence is responsible for the lethality.
We cannot be certain of the location of the fusion points between chromosome 4 and the piece of * FM7a * that was translocated. The appended chromosome * FM7a * material could be in either orientation, and on either arm of chromosome 4. Also, we cannot tell whether the telomere of the Ts ( 1 ) element is the bona fide 1L telomere, the 4R telomere, another telomere, or the acquisition of telomeric activity by other DNA. These ambiguities do not affect our overall assessment of the structure of * FM7-A * , and our best guess is shown in the Figure 1A .
Our finding of a novel rearranged X and 4 chromosome pair in the * FM7-A * stock may be important in a number of situations. First, whole genome sequencing of mutations (or heterochromatic polymorphisms) balanced with * FM7-A * may yield erroneous results or assemblies. Second, “Basc” tests (Ashburner et al. 2011) using * FM7-A * are expected to uncover mutations on both the X and 4 components, altering the distribution, identity, frequency, and linkage of such alleles. Third, the use of * FM7-A * cannot be used to measure either X or 4 nondisjunction (or, of course, co-nondisjunction). Fourth, progeny counts of phenotypes linked to either the X or 4 may be altered when segregation products are present in the genotype. Fifth, and relevant to our original purpose, experiments to alter, score, or balance bobbed alleles are not reliable with an unmarked bobbed ^+^ chromosome *4 * in the background. On the brighter side, our ability to generate X / X ; Ts ( 4 )/ Ts ( 4 ) flies indicates that the Ts ( 4 ) element may be used to increase the ploidy of the tip of the X chromosome, or allow the use of the element as a bobbed ^+^ chromosome (or a Dp ( *1Lt *
1B2 )) whose segregation is independent of the X or Y .
Methods
General husbandry and genetic crosses
Flies were fed standard molasses agar food and reared at 25°C. During crosses, parents were transferred on day 6 after introducing females to males and dumped from the transfer vial on day 11; virgins were collected starting day 9 and males were collected on day 16. For fly counts, data were collected on days 14 and 18. No flies were taken or counted after day 18.
For crosses testing the linkage of chromosome
- 4* genes, virgin females of the desired mutants were crossed to
FM7-A * males, and F1 progeny scored for the phenotype of the mutation in question. For * JY * _ alpha _ , whose phenotype is sterility, males were crossed to C ( 1 ) DX virgins and presence/absence of any first instar larvae was scored after 5 days.
To determine if Ts ( 4 ) could be homozygosed in the absence of Ts ( 1 ), * ci * ^D^ virgin females were crossed to * FM7-A * males and the female and male offspring bred inter se . The non-Bar non-Tubby non-cubitus-interruptus progeny were then transferred and scored for three subsequent generations to assure the absence of the * ci * ^D^ and Ts ( 1 ) chromosomes.
To rescue Y -linked * bobbed * ( rDNA ) mutations, * FM7-A * virgin females were crossed to males bearing either Y , 484 or Y , 183 . Male offspring were then outcrossed to C ( 1 ) DX females. Rescue was determined by measuring the penetrance and expressivity of the * bobbed * alleles on the *Y * chromosomes.
For cytology, Ts ( 1 )/ X ; Ts ( 4 )/ 4 females and Ts ( 1 )/ Y ; Ts ( 4 )/ 4 males (in panels B and C, respectively) were generated by crossing * FM7-A * virgin females to y w/Y , B ^S^ males. C ( 1 ) DX / Y ; Ts ( 4 )/ 4 females and C ( 1 ) DX / Ts ( 1 ); Ts ( 4 )/ 4 metafemales (in panels D and F, respectively) were generated by crossing C ( 1 ) DX virgin females to * FM7-A * males. Generation of C ( 1 ) DX / Y ; Ts ( 4 )/ 4 females (in panel E) was done as described in the preceding paragraph, using the chromosome Y , rDNA ^SK19-bb-l^ (Kindelay and Maggert 2024).
To determine if Ts ( 1 ) could be homozygosed in the absence of Ts ( 4 ), * FM7-A * virgin females were crossed to ci gvl ey sv . Female progeny were each backcrossed to ci gvl ey sv males. cubitus-interruptus grooveless eyeless shaven Bar Tubby females and cubitus-interruptus grooveless eyeless shaven non-Bar non-Tubby males were bred inter se for 6 generations to look for cubitus-interruptus grooveless eyeless shaven Bar Tubby males; none were found (despite 79 cubitus-interruptus grooveless eyeless shaven Bar Tubby female, 147 cubitus-interruptus grooveless eyeless shaven non-Bar non-Tubby female, and 159 cubitus-interruptus grooveless eyeless shaven non-Bar non-Tubby male siblings).
To ascertain rescue by * Dp(1;f)1187 * , cubitus-interruptus grooveless eyeless shaven Bar Tubby females from the above cross were crossed to * Dp(1;f)1187 * males. Progeny were scored for the presence of non-yellow cubitus-interruptus grooveless eyeless shaven Bar Tubby males.
Neuroblast squashes for mitotic chromosomes
Neuroblasts were done as described (Sullivan et al. 2008). Slides were visualized on a Zeiss AxioSkop-II mot using 100X oil optics (Zeiss Plan-NEOFLUAR, 100X oil, NA 1.3). Images were taken with an AxioCam and post-processed for bright/contrast using GIMP 2.10.34 (rev 3) on a MacBook Pro (2023).
Reagents
Drosophila strain genotypes (and their respective Bloomington Drosophila Stock Center numbers, at https://bdsc.indiana.edu, as of May 24, 2025):
C ( 1 ) DX : * C * ( * 1 * ) * DX * , * y * ^1^ * f * ^1^ * bb * ^0^ / * w * ^1118^ * shi * ^1^ / * Dp * ( * 1 * ; * Y * ) * B * ^ S ^ * Y * (lab strain)
ci gvl ey sv : * ci * ^1^ * gvl * ^1^ * ey * ^R^ * sv * ^n^ (RRID:BDSC_641)
ci ^D^ : * bt * ^D^ / In ( 4 ) * ci * ^D^ , * ci * ^D^ * pan * ^ciD^ (RRID:BDSC_638)
Dp(1;f)1187 : Df ( 1 ) * sc * ^8^ , * y * ^1^ * sc * ^8^ * w * ^a^ ; * Dp * ( * 1 * ; * f * ) * 1187 * , P { * ry * ^+t7.2^
PZ } 0801 P { * ry * ^+t7.2^
PZ } 8-23 y ^+^ ; * ry * ^506^ (RRID:BDSC_3940)
FM7-A : * FM7a * , P { * w * ^+mC^
Tb * ^1^ } * FM7-A * (RRID:BDSC_36489)
FM7a : * FM7a * (isolated from RRID:BDSC_3717)
JY alpha : * y * ^1^ * w * ^*^ ; TI { GFP ^3xP3.cLa^
CRIMIC.TG4.0 } * JYalpha * ^CR70483-TG4.0^ / In ( 4 ) * ci * ^D^ , * ci * ^D^ * pan * ^ciD^ , * y * ^1^ * w * ^^ ; TI { TI } FRT101F * JYalpha * ^l^ / In ( 4 ) * ci * ^D^ , * ci * ^D^ * pan * ^ciD^ , and * y * ^1^ * w * ^^ ; TI { TI } FRT101F * JYalpha * ^P^ / TI { GMR
HMS04515 } * Gat * ^eya^ (RRID:BDSC_97185, RRID:BDSC_605322, and RRID:BDSC_605323)
M101 : Df ( 4 ) * M101-63a * / In ( 4 ) * ci * ^D^ , * ci * ^D^ * pan * ^ciD^ and Df ( 4 ) * M101-62f /Dp * ( 2 ; 4 ) * ey * ^D^ , * Ablp * ^eyD^ : * ey * ^D^ (RRID:BDSC_1082 and RRID:BDSC_9433)
PlexB : * y * ^1^ ; * ry * ^506^ ; Df ( 4 ) * M101-62f * , P {y ^+mDint2^ w ^BR.E.BR^
SUPor-P } * PlexB * ^KG00878^ / In ( 4 ) * ci * ^D^ , * ci * ^D^ * pan * ^ciD^ , and * y * ^1^ * w * ^1118^ ; Mi { DH.1 } * PlexB * ^MI15559-DH.GT-TG4.1^ / In ( 4 ) * ci * ^D^ , * ci * ^D^ * pan * ^ciD^ (RRID:BDSC_14579 and RRID:BDSC_93694)
y w : * y * ^1^ * w * ^67c23^ (lab strain)
Y , 183 : * y * ^1^ / * y * ^+^ Y , P { * w * ^-^
RSw } 10B , rDNA ^bb-183^ (from Paredes and Maggert 2009)
Y , 484 : * y * ^1^ / * y * ^+^ Y , P { * w * ^-^
RSw } 10B , rDNA ^bb-l-484^ (from Paredes and Maggert 2009)
Y , rDNA ^SK19-bb-l^ : * y * ^1^ * w * ^67c23^ / Y , P { * w * ^+^ * y * ^+^
SUPorP } 590 , rDNA ^SK19-bb-l^ (from Kindelay and Maggert 2024)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gatti M Pimpinelli S 1992 Functional elements in Drosophila melanogaster heterochromatin.Annu Rev Genet 260066-419723927510.1146/annurev.ge.26.120192.0013231482113 · doi ↗ · pubmed ↗
- 2Kindelay SM Maggert KA 202478 Insights into ribosomal DNA dominance and magnification through characterization of isogenic deletion alleles.Genetics 22730016-673110.1093/genetics/iyae 06338797870 · doi ↗ · pubmed ↗
- 3Lattao R Bonaccorsi S Guan X Wasserman SA Gatti M 2011725 Tubby-tagged balancers for the Drosophila X and second chromosomes.Fly (Austin)541933-693436937010.4161/fly.5.4.1728321785267 PMC 3266079 · doi ↗ · pubmed ↗
- 4Öztürk-Çolak A Marygold SJ Antonazzo G Attrill H Goutte-Gattat D Jenkins VK Matthews BB Millburn G Dos Santos G Tabone CJ Fly Base Consortium 202421 Fly Base: updates to the Drosophila genes and genomes database.Genetics 0016-673110.1093/genetics/iyad 21138301657 PMC 11075543 · doi ↗ · pubmed ↗
- 5Paredes S Maggert KA 2009126 Expression of I-Cre I endonuclease generates deletions within the r DNA of Drosophila.Genetics 18140016-67311661167110.1534/genetics.108.09909319171942 PMC 2666528 · doi ↗ · pubmed ↗
- 6Renden R Berwin B Davis W Ann K Chin CT Kreber R Ganetzky B Martin TF Broadie K 2001816 Drosophila CAPS is an essential gene that regulates dense-core vesicle release and synaptic vesicle fusion.Neuron 3130896-627342143710.1016/s 0896-6273(01)00382-811516399 · doi ↗ · pubmed ↗
- 7Sun X Le HD Wahlstrom JM Karpen GH 200321 Sequence analysis of a functional Drosophila centromere.Genome Res 1321088-905118219410.1101/gr.68170312566396 PMC 420369 · doi ↗ · pubmed ↗