Gracilariopsisgrevogerungii (Gracilariales, Rhodophyta), a new species of marine algae from Indonesia

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
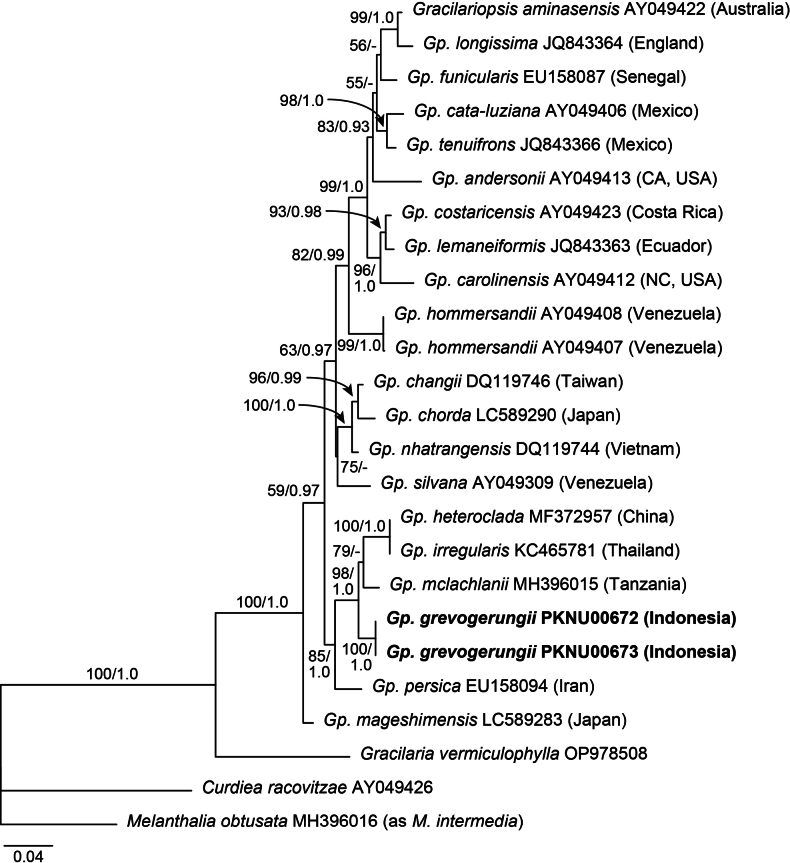 Figure 1
Figure 1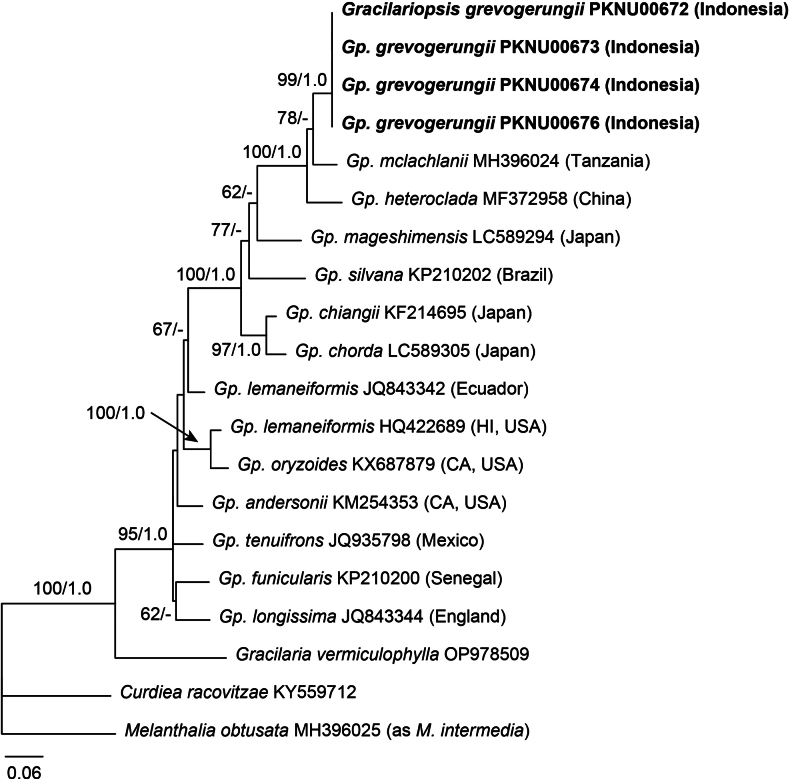 Figure 2
Figure 2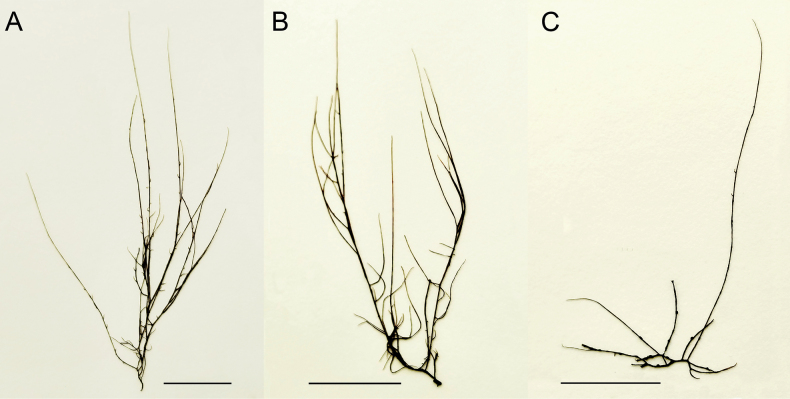 Figure 3
Figure 3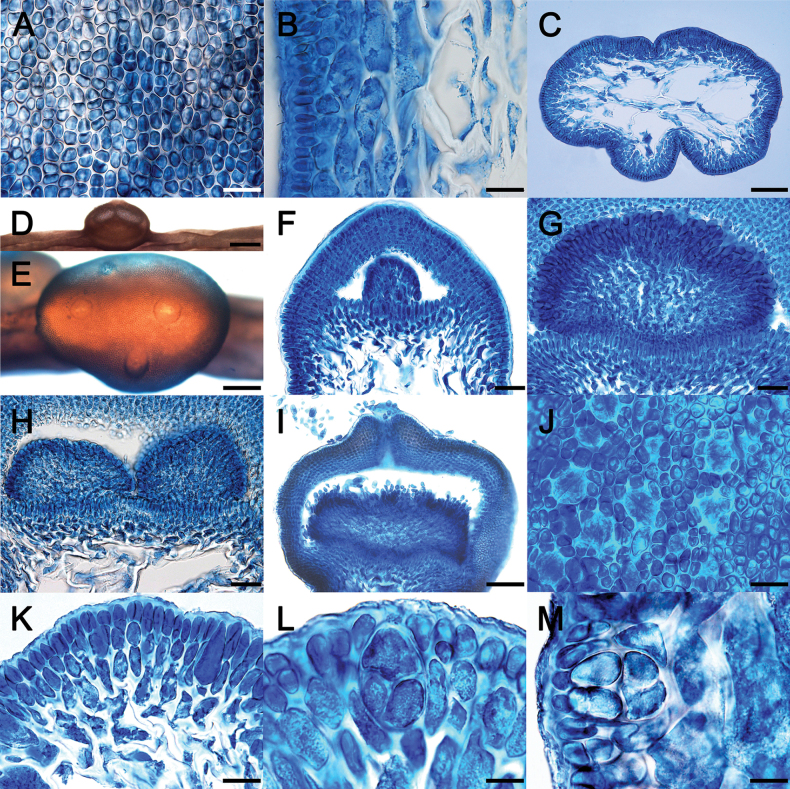 Figure 4
Figure 4| Voucher code | Collection site and date | COI-5P | |
|---|---|---|---|
| PKNU00672 (isotype) | Tamarind Beach, Nusa Lembongan Island, Bali, Indonesia; |
|
|
| PKNU00673 (holotype) | Tamarind Beach, Nusa Lembongan Island, Bali, Indonesia; |
|
|
| PKNU00674 (isotype) | Tamarind Beach, Nusa Lembongan Island, Bali, Indonesia; |
| – |
| PKNU00676 (isotype) | Tamarind Beach, Nusa Lembongan Island, Bali, Indonesia; |
| – |
| Type locality | Tamarind Beach, Lembongan Island, Bali, Indonesia | Wu-Shih-Bi Harbor, Tou-Cheng Township, Taiwan | Enoura, Namazu city, Shizuoka Pref., Japan | Yinggehai, Hainan, Guangdong, China | Ao Len, Trat Peninsula, Thailand | Nungwi Maraní, Unguja Island, Zanzibar, Tanzania | Cua Be, Nha Trang, Southern Vietnam |
| Thallus length | up to 13 cm | up to 22 cm | up to 200 cm | up to 70 cm | up to 10 cm | >150 cm | up to 18 cm |
| Main axes | percurrent, terete, 640 μm in diam. | terete, 3 mm in diam., up to 8 main axes arising from a holdfast | more or less percurrent, cylindrical, compressed, up to 5 mm in diam. | percurrent or not, cylindrical, up to 3 mm in diam. | percurrent, cylindrical, 2–2.5 mm in diam. | percurrent or not, cylindrical throughout, 1–3 mm in diam. | cylindrical to terete, up to 2.2 mm in diam., up to 15 main axes arising from a holdfast |
| Indeterminate branches | sparsely irregular | 5–7 densely clustered in the middle of axes, regenerating branches at the tip | alternate or irregular, long | long, up to four orders, easily broken, irregularly alternate, secund or furcate | very irregular to secund, sometimes inflated in middle, up to third orders | unbranched at the base, scattered, alternate to irregular, up to four orders | branched 1–2 orders from the base |
| Determinate branches | filiform, up to 5 mm long, scarce, irregular | absent | filiform, absent in young thalli, but often numerous, short | short, spinose, gradually tapered, non-constricted at the base | short, sometimes spine-like, frequently crowded | absent | numerous, racemose |
| Cortex | 1–2 layers of small cells | up to 3 layers of ovoid cells | up to 3 layers of globular cells with dense cytoplasm, with the subcortex of 3–4 layers of elongated cells | 2–3 layers of small, roundish cells | 1–2 layers of cortical cells | 2 layers of isodiametric to elongate cells, with the subcortex of 1–3 layers | 3–4 layers of ovoid cells, 6–7 μm in diam., with the subcortex of 1–3 layers |
| Medulla | large, thin-walled cells | large thin-walled cells | 5–7 layers of large, polygonal to spherical, vacuolated cells | large, parenchymatous cells | 5–12 cell layers | Large globose cells, thin-walled, highly vacuolated | large, thin-walled, vacuolated cells |
| Cystocarp | hemispherical, scattered on main axes or branches | dome-shaped, broad-based, carposporangia in branched chains | slightly beaked, constricted at bases | prominently protruding or subconical, around 1,000 μm in diam., non-constricted at the base | dome-shaped, not constricted, 200 μm in diam. | prominent, not constricted at the base | prominent, not constricted at the base |
| Pericarp | 9–14 cell layers | 10–14 cell layers | 6–8 cell layers | 7–8 cell layers | 10–12 cell layers | 8–13 cell layers | 11–17 cell layers |
| Spermantangia | not found | not found | scattered, continuous over branch surface | scattered, continuous over branch surface | continuous or discontinuous cluster | scattered, irregular pale patches | superficial |
| Tetrasporangia | scattered, cruciate, 20–34 × 19–24 μm in size | scattered, cruciate, 40–50 × 25–30 μm in size | scattered on surface, cruciate, 46–56 × 26–35 μm in size | scattered, cruciate to irregularly tetrahedral, 33–36 × 16–26 μm in size | cruciate, 28–35 µm in diam. | scattered, decussate to cruciate, 20–60 × 15–32 μm in size | cruciate, 20–30 × 10–20 μm in size |
| Distribution | Southern Indonesia | Taiwan, Japan | China, Japan, Korea | China, Malaysia, Philippines | Thailand | Tanzania | Vietnam |
| Reference | This study |
| Le and Lin 2006 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine and coastal plant biology · Echinoderm biology and ecology · Marine and Coastal Ecosystems
Introduction
Gracilariopsis E.Y.Dawson (Gracilariaceae) is generally known as one of the major sources of agar used in food, healthcare and biotechnology industries. Gracilariopsis was established by Dawson (1949) to accommodate species that were previously placed in Gracilaria Greville and presented a small–celled, broad-based gonimoblast and the absence of nutritive filaments connecting gonimoblast with the pericarp. The established type species for the genus was Gp.sjoestedtii (Kylin) E.Y.Dawson. However, Papenfuss (1967) merged Gracilariopsis into Gracilaria because of the lack of morphological difference at the genus level based on observations of Gp.sjoestedtii and Gracilariaverrucosa (Hudson) Papenfuss, the generitype of Gracilaria. Ohmi (1958) reinstated Gracilariopsis based on the observation of Gp.chorda (Holmes) Ohmi from Japan, whereas Yamamoto (1978) included it within Gracilaria, despite his beautiful illustrations showing the superficial formation of spermatangia and the absence of nutritive tubular filaments in cystocarps. Fredericq and Hommersand (1989) reinstated Gracilariopsis including four species which have superficial spermatangia and lack nutritive tubular cells. Gracilariopsislemaneiformis (Bory) Dawson, Acleto & Foldvik was designated as the generitype because of its priority over Gp.sjoestedtii.
Gracilariopsis has been robustly supported in phylogenies based on nuclear small subunit ribosomal DNA (SSU rDNA) and plastid rbcL gene sequence (Bird et al. 1994; Gurgel et al. 2003a, b). Gurgel et al. (2003b) designated the generitype of Gracilariopsis as Gp.andersonii (Grunow) Dawson, based on material from the northwest coast of America. The authors confirmed that Gp.lemaneiformis, for a long time considered a widespread species, is likely restricted to the Peruvian coast, and the collections of Gp.lemaneiformis from northwestern America correspond to Gp.andersonii. They also indicated that the collections of Gp.lemaneiformis from China and Japan may represent an undescribed species that is related to Gp.heteroclada J.-F.Zhang & B.-M.Xia.
The knowledge on species diversity of Gracilariopsis has improved with the increase of sequences availability, especially COI-5P and rbcL data from various regions (Bellorin et al. 2008; Gurgel et al. 2003a, b; Iyer et al. 2005; Le and Lin 2006; Lin 2008; Muangmai et al. 2014; Suzuki and Terada 2022). Recently, phylogenetic relationships of species within Gracilariopsis were further investigated using organellar genomes (Iha et al. 2018; Lyra et al. 2021). To date, a total of 23 species have been listed in the AlgaeBase (Guiry and Guiry 2025). Because most species of Gracilariopsis lack distinctive vegetative and reproductive characteristics that allow for reliable differentiation from other species (Bellorin et al. 2008), DNA sequence data is needed to evaluate the species diversity of Gracilariopsis.
Weber-van Bosse (1928) reported Gracilariopsislemaneiformis as Gracilarialemaneiformis (Bory) Greville in Indonesia, but it has not been collected since (Meinita et al. 2021). The objectives of this study were to assess the occurrence of Gracilariopsis species in southern Indonesia and to elucidate the taxonomic identities of the species present. During collection trips in southern Indonesia, several Gracilariopsis-like plants were collected from Nusa Lembongan Island, Bali. Based on detailed morphological comparisons and analyses of plastid rbcL and mitochondrial COI-5P sequences, we recognize these specimens as representing a new species of Gracilariopsis.
Materials and methods
Specimens were collected in June 2017 at Tamarind Beach (8°40'47.51"S, 115°26'10.20"E), Nusa Lembongan Island, Bali, Indonesia (Table 1). Specimens were mounted on herbarium sheets and tissue samples were dehydrated in silica gels for DNA sequencing. For anatomical observation, plants were sectioned using razor blades and were stained with 1% aqueous aniline blue. Photographs were taken with a DP-71 camera (Olympus, Tokyo, Japan) mounted on a BX-51 microscope (Olympus). Vouch specimens are housed at the Herbarium of the Department of Marine Biology, Pukyong National University, Busan, Korea (PKNU).
DNA extraction, polymerase chain reaction amplification, and sequencing procedures followed Boo et al. (2016). The primer set used for amplifying and sequencing was F145, F754, R898, and R1442 for plastid rbcL (Kim et al. 2010), and GazF1 and GazR1 for mitochondrial COI-5P (Saunders 2005). All sequences were aligned together with publicly available sequences of Gracilariopsis species in GenBank, using the MUSCLE algorithm in MEGA7 (Kumar et al. 2016) with default parameters and the alignment was manually adjusted. Curdiearacovitzae Hariot, Gracilariavermiculophylla (Ohmi) Papenfuss, and Melanthaliaobtusata (Labillardiere) J.Agardh were used as outgroups based on previous studies of the Gracilariaceae (Gurgel and Fredericq 2004; Bellorin et al. 2008; Iha et al. 2018). Sequences generated in the present study were deposited in GenBank (PV106179–PV106182, PV424435, PV424436).
Phylogenies of both datasets were reconstructed using maximum likelihood (ML) and Bayesian inference (BI). The ML analysis was performed using the W-IQ-tree webserver (Trifinopoulos et al. 2016) with 1,000 ultrafast bootstrap (BS) replications (-bb 1000) and model test option (-m TEST). The BI analysis was performed with MrBayes v.3.2.1 (Ronquist et al. 2012) using the Metropolis-coupled Markov Chain Monte Carlo (MC3) with the best-fitting substitution model selected by IQ-tree. Four million generations of two independent runs were performed with four chains and sampling trees every 100 generations. The burn-in period was identified graphically by tracking the likelihoods at each generation to determine whether they reached a plateau. Twenty-five percent of saved trees were removed, and the remaining trees were used to infer Bayesian posterior probabilities (BPP).
Results
Molecular phylogeny
Six sequences were generated in the present study, two rbcL and four COI-5P sequences. A total of 25 rbcL sequences were aligned, including 23 publicly available sequences of Gracilariopsis and three outgroups. In the rbcL phylogeny (Fig. 1), the Indonesian taxon was distinct from the other species in the genus, and formed a sister relationship with the clade of Gp.heteroclada, Gp.irregularis (I.A.Abbott) N.Muangmai, A.Chirapart & A.Lewmanomont, and Gp.mclachlanii Buriyo, Bellorin & M.C.Oliveira (98% MLBS, 1.0 BPP). The pairwise divergence of rbcL sequences between the Indonesian taxon and related species was 2.6–3.3%, with identical sequences among the Indonesian specimens.
Maximum likelihood (ML) phylogeny of Gracilariopsis using plastid rbcL sequences. ML bootstrap values (≥50%) and Bayesian posterior probabilities (≥0.9) are shown at branches. Bold letters indicate Gp.grevogerungii sp. nov.
In the COI-5P phylogeny (Fig. 2), the Indonesian taxon placed in a position largely congruent with that in the rbcL phylogeny, forming a clade with Gp.heteroclada and Gp.mclachlanii (100% MLBS, 1.0 BPP). The pairwise divergence of COI-5P sequences between the Indonesian taxon and related species was 4.7–6.1%, with identical sequences among the Indonesian specimens.
Maximum likelihood (ML) phylogeny of Gracilariopsis using mitochondrial COI-5P sequences. ML bootstrap values (≥50%) and Bayesian posterior probabilities (≥0.9) are shown at branches. Bold letters indicate Gp.grevogerungii sp. nov.
Morphological observations
Details of morphological features are introduced in description and illustration below (Figs 3, 4). The Gracilariopsis species from Indonesia is characteristic of the genus in the absence of nutritive tubular cells between the gonimoblast and pericarp. The species can be distinguished from other species of Gracilariopsis by a combination of terete main axes with irregular branches with sparse, filiform branchlets (up to 4 mm), scattered tetrasporangia on axes and branches, and hemispherical cystocarps with up to 3 ostioles and 9–14 cell-layered pericarps. A morphological comparison of Indonesian Gracilariopsis with other terete species of the genus is given in Table 2. However, without DNA sequences, it is difficult to recognize the Indonesian Gracilariopsis.
A–C. Habit of Gracilariopsisgrevogerungii sp. nov. A. Holotype specimen (PKNU00673) from Tamarind Beach, Nusa Lembongan Island, Bali, Indonesia; 21 June, 2017; B. Isotype specimen having tetrasporangia (PKNU00676); C. Isotype specimen bearing cystocarps (PKNU00672). Scale bars: 2 cm (A–C).
A–M. Morphology and anatomy of Gracilariopsisgrevogerungii sp. nov. A. Surface view of axis showing irregularly arranged cortical cells; B. Longitudinal section of axis showing compact cortex and medulla; C. Transverse section of axis showing abrupt transition in cells size from cortex to medulla; D. Hemispherical cystocarp; E. Three ostioles on a single cystocarp; F. Longitudinal section of cystocarp showing early stage of gonimoblast; G. Longitudinal section of cystocarp lacking nutritive tubular cells between gonimoblast and pericarp; H. Two gonimoblasts sometimes formed within a single cystocarp; I. Longitudinal section of cystocarp showing releasing carpospores through ostiole; J. Tetrasporangia partially immersed by cortical cells; K. Tetrasporangial initial formed from inner cortical cells; L, M. Decussately to cruciately divided tetrasporangia. Scale bars: 20 μm (A, B, J–M); 100 μm (C, I); 400 μm (D); 200 μm (E); 40 μm (F–H).
Gracilariopsis
grevogerungii
Taxon classificationPlantaeGracilarialesGracilariaceae
G.H.Boo & I.K.Hwang sp. nov.
68F9B310-9ED1-5C47-BAE5-E8AB15D0AE09
Description.
Thallus up to 13 cm tall, solitary, yellow-green to pale red in color. Main axes cylindrical throughout, about 640 μm in diameter, arising from a small disk-like holdfast (Fig. 3). Indeterminate branches often unbranched at the base, scattered, alternate to irregularly arising to 1–2 orders, slightly constricted at the base and tapering gradually toward apices. Determinate branches up to 4 mm long, irregularly alternate or scattered. Cortices composed of two to three layers of small isodiametric or anticlinally elongate cells (Fig. 4A, B), measuring 3.2–6.4 × 7.5–12.8 μm in size, with dense content, heavily pigmented, connected only with their parental cells by primary pit connections. Medulla composed of large globose cells, 114–163 μm in diameter in transverse sections, thick-walled and highly vacuolated, lacking pigments. Transition in cell size from cortex to medulla abrupt (Fig. 4C). Large basal cells of deciduous hairs frequently occurring near the surface. Cystocarps 492–805 μm in diameter and 689–884 μm in height, hemispherical, scattered on main axes or determinate branches, slightly constricted at the base. Cystocarps slightly constricted at the base, with up to three ostioles (Fig. 4D, E). Carposporangia initials formed in long chains and radially elongated (Fig. 4F, G). Two gonimoblasts formed in a single cystocarp (Fig. 4H). Mature cystocarps released carpospores through ostioles (Fig. 4I). Pericarps about 115 μm thick, formed by 9–14 cell layers. Tetrasporangia embedded in the cortex (Fig. 4J). Tetrasporangial initials formed from inner cortical cells (Fig. 4K). Tetrasporangia decussately or cruciately divided, ovoid, 20–34 × 19–24 μm in size (Fig. 4L, M).
Diagnosis.
Diagnosed by a combination of characters: simple terete axes with irregular long branches and scarce short branchlets, hemispherical cystocarps with up to 3 ostioles, 9–14 cell-layered pericarp, and DNA sequences (accession number: PV106182 for rbcL and PV106180 for COI-5P).
Type.
Indonesia • Bali, Nusa Lembongan Island, Tamarind Beach, 8°40'47.51"S, 115°26'10.20"E, 21 Jun. 2017, collected by Sung Min Boo without collection numbers (Holotype: PKNU00673!; Isotypes: PKNU00672!, PKNU00674!, and PKNU00676!; Paratype: PKNU00675!). Types are deposited in the herbarium of the Department of Marine Biology, Pukyong National University, Busan, Korea (PKNU).
Habitat and distribution.
Gracilariopsisgrevogerungii grows on intertidal to shallow subtidal sandy-muddy substrates. It is currently demonstrated in the type locality solely using DNA sequences, but its range is likely expanded to the surrounding waters with additional collections (see Discussion).
Etymology.
Species epithet is given in honor of Dr Grevo Soleman Gerung for his contributions to the knowledge of seaweed diversity in Indonesia.
Discussion
Gracilariopsisgrevogerungii is the only described southern Indonesian species of Gracilariopsis investigated by both rbcL and COI-5P sequences, as well as morphology. The cystocarp anatomy revealed the chains of carposporangia and the lack of nutritive tubular cells, typical characters of Gracilariopsis. Gracilariopsisgrevogerungii was likely misidentified as Gp.lemaneiformis based on specimens collected in Flores Island and Tanah Djampea, Indonesia (Weber-van Bosse 1928). Its illustration of cystocarp having 10–17 cell layers in pericarp and lacking the nutritive tubular cells matches well with Gp.grevogerungii.
Gracilariopsisgrevogerungii has likely been misidentified as terete species of Gracilaria. For example, during the present study, Gracilariaedulis (S.G.Gmelin) P.C.Silva was collected in Benoa Bay, Bali, very close to Nasa Lembongan Island. It is a common species in Indonesia and also included in the export list of Indonesian hydrocolloid seaweeds (Meinita et al. 2021; Basyuni et al. 2024). However, COI-5P sequences from our collections of G.edulis (GHB, unpubl.) revealed its difference from Gp.grevogerungii. Again, Gracilaria is well segregated from Gracilariopsis by the presence of nutritive filaments in the cystocarp and by molecular data (Gurgel et al. 2003b; Bellorin et al. 2008).
Gracilariopsis sp. from Zamboanga city, Philippines, which lacks fine, determinate branchlets (Hurtado-Ponce and Liao 1998), is morphologically similar to Gp.grevogerungii. DNA sequences from the Philippine collection are necessary to confirm its identity. Gracilariopsislemaneiformis has still been reported in China (Wang et al. 2023), even though it has been confirmed by rbcL sequences that the Chinese specimens do not correspond to Gp.lemaneiformis (whose type locality is in Peru), but to Gp.chorda (whose type locality is in Japan). Additional sampling at sand-muddy coastal regions will likely extend the range of Gp.grevogerungii into other locations in Indonesia or surrounding waters.
Two species of Gracilariopsis were closely related to Gp.grevogerungii in both rbcL and COI-5P phylogenies. Gracilariopsismclachlanii, originally described for Tanzanian specimens, is recognized by large cylindrical form of thallus, 8–13 cell layers of pericarp, and cleavage of spermatangia from spermatangial mother cells through concavo-convex oblique septa (Bellorin et al. 2008). Gracilariopsisheteroclada, first described for Chinese specimens, is characterized by slender, filiform indeterminate branches that arise alternatively and are beset with fine, regularly-disposed determinate branchlets (Zhang and Xia 1988; Hurtado-Ponce and Liao 1998).
Several species of Gracilariopsis, including Gp.heteroclada, have been previously recognized in Southeast Asia (Pham 1969; Le and Lin 2006; Lin 2008; Muangmai et al. 2014). Most species except foliose Gp.mageshimensis Mas.Suzuki & R.Terada from Japan resemble Gp.grevogerungii in their habits with filiform thalli having irregular lateral branches from main axes (Table 2). Of these, Gp.chiangii Showe M.Lin, Gp.heteroclada, and Gp.nhatrangensis Nhu Hau Le & Showe M.Lin have been well delimited by DNA sequences and morphology (Le and Lin 2006; Lin 2008; Wang et al. 2023). Gracilariopsischiangii is characterized by small-sized thallus (15–22 cm in length) and 1–2(–3) orders of branches from the base to middle part of thallus (Lin 2008). Gracilariopsisnhatrangensis is characterized by 1–15 main branches (up to 18 cm tall), arising from a discoid holdfast and numerous, racemose branchlets on main axes (Le and Lin 2006).
Gracilariopsisirregularis from Thailand is distinguished by having short (up to 10 cm) and succulent axes with an irregular secondary branching pattern (Muangmai et al. 2014). However, Gp.irregularis requires reexamination by COI-5P and other molecular markers because of its homogeneity to Gp.heteroclada in rbcL. It is important to reexamine the holotypes or topotype materials of two Vietnamese species, Gp.nganii Pham and Gp.phanthiens Pham, which have not been recorded since the original publication (Le and Lin 2006; Pham 1969), to elucidate their relationships with Gp.grevogerungii and other Gracilariopsis species. Because most Gracilariopsis species have similar vegetative traits (Bellorin et al. 2008), it is difficult to discriminate Gp.grevogerungii from other cylindrical species of Gracilariopsis in Southeast Asia without DNA sequences.
Conclusions
DNA sequence analyses were essential for the recognition of a new tropical species, Gracilariopsisgrevogerungii, from southern Indonesia. This study highlights the critical role of molecular data in elucidating the biodiversity of a morphologically simple group of marine red algae. Gracilariopsisgrevogerungii, along with Gracilaria species, is expected to be included in the export list of Indonesian hydrocolloid seaweeds. Our integrative taxonomy, combining molecular and morphological evidence, contributes to the clarification of commercial seaweed species and the cataloguing of red algal biodiversity. This study emphasizes the necessity for continued taxonomic and molecular investigations of the agar-producing genus Gracilariopsis in Indonesia and adjacent regions.
Supplementary Material
XML Treatment for Gracilariopsis grevogerungii
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abbott IA (1988) Some species of Gracilaria and Polycavernosa from Thailand. In: Abbott IA (Ed.) Taxonomy of Economic Seaweeds with Reference to Some Pacific and Caribbean Species (Vol.II). California Sea Grant College Program, La Jolla, 137–150.
- 2Basyuni M Puspita M Rahmania R Albasri H Pratama I Purbani D Aznawi AA Mubaraq A Al Mustaniroh SS Menne F Rahmila YI Salmo III SG Susilowati A Larekeng SH Ardli E Kajita T (2024) Current biodiversity status, distribution, and prospects of seaweed in Indonesia: A systematic review. Heliyon 10: e 31073. 10.1016/j.heliyon.2024.e 31073 PMC 1110982938779002 · doi ↗ · pubmed ↗
- 3Bellorin AM Buriyo A Sohrabipour J Oliveira MC Oliveira EC (2008) Gracilariopsismclachlanii sp. nov. and Gracilariopsispersica sp. nov. of the Gracilariaceae (Gracilariales, Rhodophyta) from the Indian Ocean.Journal of Phycology 44: 1022–1032. 10.1111/j.1529-8817.2008.00552.x 27041621 · doi ↗ · pubmed ↗
- 4Bird CJ Ragan MA Crichley AT Rice EL Gutell RR (1994) Molecular relationships among Gracilariaceae (Rhodophyta): Further observations on some undetermined species.European Journal of Phycology 29: 195–202. 10.1080/09670269400650641 · doi ↗
- 5Boo GH Le Gall L Miller KA Freshwater DW Wernberg T Terada R Yoon KJ Boo SM (2016) A novel phylogeny of the Gelidiales (Rhodophyta) based on five genes including nuclear Ces A, with descriptions of Orthogonacladia gen. nov. and Orthogonacladiaceae fam. nov.Molecular Phylogenetics and Evolution 101: 359–372. 10.1016/j.ympev.2016.05.01827223999 · doi ↗ · pubmed ↗
- 6Dawson EY (1949) Studies of the northeast Pacific Gracilariaceae.Occasional Papers of the Allan Hancock Foundation 7: 1–105.
- 7Fredericq S Hommersand MH (1989) Comparative morphology and taxonomic status of Gracilariopsis (Gracilariales, Rhodophyta).Journal of Phycology 25: 228–241. 10.1111/j.1529-8817.1989.tb 00117.x · doi ↗
- 8Guiry MD Guiry GM (2025) Algae Base. World-wide electronic publication, National University of Ireland, Galway. https://www.algaebase.org/ [accessed 5 January 2025]