Application of Green Cobalt Nanoparticles in the Diet of Broiler Chickens to Improve Sustainable Production and Health
Fayiz M. Reda, Abdullah S. Alawam, Hemat K. Mahmoud, Mohamed T. El‐Saadony, Ayman S. Salah, Hassan A. Rudayni, Ahmed A. Allam, Karima El‐Naggar, Mahmoud Alagawany, Most Khairunnesa

TL;DR
Green cobalt nanoparticles added to chicken feed improve growth, health markers, and immune function in broiler chickens.
Contribution
Demonstrates that green cobalt nanoparticles in feed enhance broiler performance and health more effectively than traditional methods.
Findings
BNCo supplementation improved body weight, weight gain, and feed conversion ratio in broilers.
Immune organs like the spleen and thymus increased in size with BNCo treatment.
Blood markers such as hemoglobin, RBCs, and WBCs improved significantly with BNCo.
Abstract
Nanoparticles (NPs), such as green cobalt NPs (CoNPs), are easier to pass through cell membranes in animals and interact rapidly with biological systems. Therefore, using green CoNPs is one of the recommendations for enhancing the bioavailability of cobalt, thereby improving its absorption. The goal of this study was to explore the influences of biological nano‐cobalt (BNCo) as a feed supplement on broiler growth performance, haematology, blood chemistry, antioxidant activities, immunological status, digestive enzymes and carcass characteristics. A total of 300 Arbour Acre broiler chicks, all unsexed, were distributed into 5 treatment groups, each containing 60 chicks, at random. Five replications of each group were formed, each containing 12 chicks. The first group received a control diet free of BNCo, whereas the second, third, fourth and fifth groups were administered diets…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
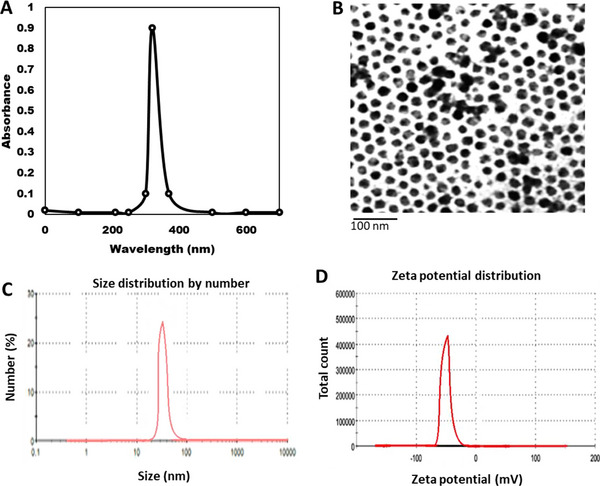 FIGURE 1
FIGURE 1| Items | Starter | Finisher |
|---|---|---|
|
| ||
| Yellow corn | 56.85 | 61.25 |
| Soybean meal | 34.30 | 32.80 |
| Soybean oil | 0.70 | 2.50 |
| Corn gluten (62% CP) | 4.50 | 0.00 |
| Di‐calcium phosphate | 0.20 | 0.25 |
| Limestone | 2.20 | 2.00 |
| Vit‐min Premixa | 0.30 | 0.30 |
| NaCl | 0.30 | 0.30 |
| DL methionine | 0.17 | 0.20 |
|
| 0.28 | 0.25 |
| Choline chloride 60% | 0.20 | 0.15 |
| Total | 100 | 100 |
|
| ||
| CP | 23.11 | 20.07 |
| ME Kcal/kg diet | 2913 | 3107 |
| Ca | 1.00 | 0.97 |
| P (Available) | 0.46 | 0.45 |
| Lysine | 1.40 | 1.29 |
| M + C | 0.92 | 0.82 |
| CF | 3.62 | 3.57 |
| Linoleic acid | 1.38 | 1.47 |
| BNCo (ppm) | |||||||
|---|---|---|---|---|---|---|---|
| Items | 0 | 100 | 200 | 300 | 400 | SEM |
|
|
| |||||||
| 1 week | 184.83 | 185.5 | 184.17 | 185.53 | 184.83 | 1.538 | 0.9718 |
| 3 week | 874.63c | 902.03ab | 912.33a | 910.87ab | 884.47bc | 8.150 | 0.0306 |
| 6 week | 2047.90c | 2143.97b | 2233.48a | 2074.20c | 2015.70c | 19.297 | 0.0001 |
|
| |||||||
| 1–3 week | 49.27c | 51.18abc | 52.01a | 51.81ab | 49.97bc | 0.595 | 0.0358 |
| 3–6 week | 55.87c | 59.14b | 62.91a | 55.40c | 53.87c | 0.941 | 0.0004 |
| 1‐6 week | 53.23c | 55.95b | 58.55a | 53.96c | 52.31c | 0.524 | <0.0001 |
|
| |||||||
| 1–3 week | 73.61 | 74.75 | 74.94 | 72.86 | 73.23 | 0.962 | 0.5444 |
| 3–6 week | 123.47 | 124.86 | 124.09 | 128.19 | 128.3 | 1.932 | 0.3099 |
| 1–6 week | 103.53 | 104.82 | 104.43 | 106.06 | 106.27 | 1.361 | 0.6106 |
|
| |||||||
| 1–3 week | 1.49 | 1.46 | 1.44 | 1.41 | 1.47 | 0.019 | 0.1253 |
| 3–6 week | 2.21ab | 2.11bc | 1.97c | 2.31a | 2.38a | 0.047 | 0.0019 |
| 1–6 week | 1.95ab | 1.87bc | 1.78c | 1.97ab | 2.03a | 0.027 | 0.0014 |
| BNCo (ppm) | |||||||
|---|---|---|---|---|---|---|---|
| Items (%) | 0 | 100 | 200 | 300 | 400 | SEM |
|
| Carcass | 73.03 | 74.46 | 74.39 | 74.19 | 73.23 | 0.938 | 0.7369 |
| Liver | 2.37 | 2.35 | 2.24 | 2.61 | 2.51 | 0.140 | 0.4668 |
| Gizzard | 3.14 | 3.17 | 3.41 | 3.27 | 3.28 | 0.141 | 0.7213 |
| Heart | 0.55 | 0.72 | 0.68 | 0.69 | 0.59 | 0.068 | 0.3779 |
| Giblets | 6.06 | 6.24 | 6.34 | 6.58 | 6.38 | 0.190 | 0.5613 |
| Dressing | 79.09 | 80.7 | 80.73 | 80.77 | 79.61 | 1.077 | 0.7321 |
| BNCo (ppm) | |||||||
|---|---|---|---|---|---|---|---|
| Items | 0 | 100 | 200 | 300 | 400 | SEM |
|
| Haemoglobin | 7.67b | 7.95ab | 8.25a | 7.85b | 7.97ab | 0.106 | 0.0404 |
| RBCs | 3.31c | 3.65bc | 4.00ab | 4.12a | 3.79ab | 0.106 | 0.0039 |
| WBCs | 19.87b | 22.68a | 22.49a | 23.44a | 23.51a | 0.623 | 0.0159 |
| BNCo (ppm) | |||||||
|---|---|---|---|---|---|---|---|
| Items | 0 | 100 | 200 | 300 | 400 | SEM |
|
| TP (g/dL) | 3.38b | 4.25a | 4.47a | 4.28a | 4.14a | 0.116 | 0.0006 |
| ALB (g/dL) | 1.95c | 2.47ab | 2.61a | 2.32b | 2.52ab | 0.076 | 0.0011 |
| GLOB (g/dL) | 1.43c | 1.77ab | 1.86a | 1.96a | 1.62bc | 0.070 | 0.0027 |
| A/G (%) | 1.36ab | 1.40ab | 1.41ab | 1.18b | 1.56a | 0.059 | 0.0330 |
| AST (IU/L) | 75.48ab | 60.79cd | 52.38d | 67.90bc | 82.18a | 3.355 | 0.0008 |
| ALT (IU/L) | 20.41ab | 17.84bc | 15.30c | 21.22ab | 23.49a | 1.154 | 0.0048 |
| Creatinine (mg/dL) | 1.05ab | 0.83bc | 0.72c | 1.01ab | 1.22a | 0.070 | 0.0061 |
| Uric acid (mg/dL) | 7.04 | 5.16 | 6.01 | 6.78 | 7.41 | 0.544 | 0.0953 |
| Urea (mg/dL) | 3.51a | 2.60bc | 2.34c | 3.59a | 3.19ab | 0.244 | 0.0206 |
| BNCo (ppm) | |||||||
|---|---|---|---|---|---|---|---|
| Items | 0 | 100 | 200 | 300 | 400 | SEM |
|
| TC (mg/dL) | 168.34ab | 153.50bc | 150.00c | 151.38c | 172.32a | 4.735 | 0.0289 |
| TG (mg/dL) | 80.03a | 64.53b | 61.69b | 56.50b | 69.63ab | 3.935 | 0.0189 |
| HDL (mg/dL) | 51.26ab | 53.28ab | 59.92a | 46.94b | 48.45b | 2.666 | 0.0490 |
| LDL (mg/dL) | 101.07ab | 87.31cd | 77.74d | 93.14bc | 109.94a | 3.822 | 0.0019 |
| VLDL (mg/dL) | 16.01a | 12.91b | 12.34b | 11.30b | 13.93ab | 0.787 | 0.0189 |
| BNCo (ppm) | |||||||
|---|---|---|---|---|---|---|---|
| Items | 0 | 100 | 200 | 300 | 400 | SEM |
|
| MDA (nmol/mL) | 0.42a | 0.22b | 0.26b | 0.23b | 0.49a | 0.042 | 0.0035 |
| SOD (U/mL) | 0.31b | 0.58a | 0.56a | 0.56a | 0.35b | 0.042 | 0.0023 |
| CAT (mg/dL) | 0.23b | 0.47a | 0.52a | 0.34ab | 0.36ab | 0.053 | 0.0273 |
| TAC (ng/mL) | 0.27c | 0.54a | 0.57a | 0.38bc | 0.43ab | 0.041 | 0.0046 |
| GSH (mg/dL) | 0.22c | 0.57ab | 0.72a | 0.52b | 0.31c | 0.047 | 0.0002 |
| BNCo (ppm) | |||||||
|---|---|---|---|---|---|---|---|
| Items (%) | 0 | 100 | 200 | 300 | 400 | SEM |
|
| Spleen | 0.08c | 0.13a | 0.15a | 0.12ab | 0.09bc | 0.01 | 0.0040 |
| Thymus | 0.39c | 0.55ab | 0.60a | 0.45bc | 0.52ab | 0.038 | 0.0262 |
| Bursa of fabricius | 0.27 | 0.34 | 0.32 | 0.29 | 0.26 | 0.032 | 0.5158 |
| BNCo (ppm) | |||||||
|---|---|---|---|---|---|---|---|
| Items | 0 | 100 | 200 | 300 | 400 | SEM |
|
| IgA (mg/dL) | 0.41c | 0.59ab | 0.72a | 0.44c | 0.49bc | 0.040 | 0.0018 |
| IgM (mg/dL) | 0.56b | 0.90a | 0.79a | 0.86a | 0.53b | 0.045 | 0.0004 |
| IgY (mg/dL) | 0.73c | 0.94ab | 1.04a | 0.96ab | 0.84bc | 0.055 | 0.0235 |
| C3 (mg/dL) | 32.23c | 52.58a | 52.03a | 45.84ab | 36.98bc | 2.733 | 0.0014 |
| Lysozyme (mg/dL) | 0.39c | 0.48ab | 0.55a | 0.45bc | 0.49ab | 0.024 | 0.0118 |
| BNCo (ppm) | |||||||
|---|---|---|---|---|---|---|---|
| Items | 0 | 100 | 200 | 300 | 400 | SEM |
|
| Amylase (μ/L) | 143.55bc | 163.14ab | 182.10a | 153.26bc | 136.90c | 6.815 | 0.0077 |
| Lipase (μ/L) | 21.77c | 47.34a | 42.22ab | 30.01bc | 23.11c | 3.800 | 0.0040 |
| Protease (μ/L) | 0.38d | 2.59ab | 3.17a | 1.73bc | 0.99cd | 0.292 | 0.0010 |
- —Deanship of Scientific Research at Imam Mohammad Ibn Saud Islamic University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Nutrition and Physiology
Introduction
1
The fields of animal and veterinary sciences are currently witnessing the advent of nano‐biotechnology for several beneficial applications, including nutritive, therapeutic and diagnostic (Patra et al. 2020; El‐Maddawy et al. 2022; AL‐Ruwad et al. 2024). Nanoparticles (NPs) of important minerals, sized between 1 and 100 nm, may provide a substitute for traditional elemental forms in animal nutrition (Mohamed et al. 2016; Swain et al. 2016; Scott et al. 2018; Kociova et al. 2020). It is posited that significantly reduced quantities of NPs will be sufficient to cover the elemental requirements of animals compared to bulk minerals (Abdollahi et al. 2020; Szuba‐Trznadel et al. 2021; Ouyang et al. 2021; Amr et al. 2025), thereby mitigating the environmental effects associated with high concentrations of inorganic salts (Vijayakumar et al. 2014; Ouyang et al. 2021). Decreased mineral concentrations in animal feeds may lead to reduced feed expenses. Furthermore, the characteristics of reduced size, enhanced similarity, considerable surface area and increased physical activity of nano‐forms of minerals may boost bioavailability in the gastrointestinal tract (GIT) of the animals (Hill et al. 2017; Youssef et al. 2019; Hidayat et al. 2021). Animals may derive advantages from the biological characteristics of NPs, including reduced dosage, decreased antagonism, increased absorption rate and improved tissue distribution in animals. The considerable capacity of NPs, especially at small dosages, has been extensively proven in animal nutrition studies for growth efficacy, feed efficacy and health conditions.
NPs of micro‐ and macro‐minerals reliably enhance body weight, average daily increase and feed conversion efficiency (Bąkowski et al. 2018; Reda et al. 2023). Nutritional products are employed to fulfil an animal's dietary requirements, enhance productivity, boost immune function and microbial composition and reduce disease risk. NPs are recognized for their antiviral, antifungal, antibacterial, antiprotozoal and antioxidant qualities, among others. Alternatives to antibiotics that promote growth and health include silver, copper, selenium, zinc and cobalt NPs (CoNPs) (Pineda et al. 2012; Abd El‐Hack et al. 2018; Hidayat et al. 2021; Ouyang et al. 2021; El‐Maddawy et al. 2022).
In animal nutrition, nanotechnology is applied for disease detection and treatment, as well as for the delivery of vitamins, minerals, probiotics and medications (Fesseha et al. 2020). Nano minerals, nano enzymes and other additives are examples of how nanotechnology is utilized in animal feed (Pundir et al. 2015; Marappan Gopi et al. 2017). By lessening the antagonistic impact of bivalent cations, particularly in minute minerals, NPs improve nutrient absorption and are advantageous for the nutrition of cattle and poultry as well as for better feed and supplemental use (Marappan Gopi et al. 2017).
CoNPs are utilized in both technical and medical industries due to their enhanced qualities (Iravani et al. 2020). Research has indicated that CoNPs can induce cell killing (Francis et al. 2021), suggesting their potential as innovative nanomedicines for use in phototherapy, thermotherapy and chemotherapy. Cobalt can enhance protein metabolism and facilitate the creation of enzymes, hence improving immunological function. Cobalt NPs are being utilized as prospective treatment agents for several infectious diseases due to their advantageous properties (Anwar, Chi Fung et al. 2019; Sharma et al. 2020). The CoNPs are recognized for their capacity to produce reactive oxygen species (ROS) that exert their inhibitory activities against certain microbes (Turecka et al. 2018). The information about the incorporation of biological cobalt NPs in poultry diets and their influence on performance and health is limited, and it is hypothesized that the addition of dietary biological nano‐cobalt (BNCo) will have beneficial effects on broiler chickens. The present trial was targeted to measure the antimicrobial properties of BNCo and its influence on growth performance, carcass traits, haematological parameters, renal and hepatic functions, immunological and antioxidant status, lipid profile and digestive enzymes in broiler chickens.
Materials and Methods
2
Biosynthesis and Characterization of Green Cobalt NPs
2.1
The produced green cobalt NPs were characterized using four instruments: ultraviolet (UV), transmission electron microscopy (TEM), Zeta sizer and Zeta potential, with the following results: The spherical BNCos absorb UV at 320 nm, have a TEM of 45–75 nm, have a Zeta potential of −24.6 mV and have a size of 48 nm (Figure 1).
Characterization of green cobalt nanoparticles (A) UV absorbance of BNCo, (B) the shape of BNCo by TEM, (C) the acute size of BNCo by zeta sizer and (D) net charge of BNCo by zeta potential.
Ethical Approval Statement
2.2
The experiment was carried out in the chicken farm within the Faculty of Agriculture, Poultry Department, Zagazig University, Zagazig, Egypt. The trial adheres to the regulations provided by the Zagazig University Ethics Committee regarding the use of experimental animals (Approval No. ZU‐IACUC/2/F/313/2023).
Animals, Design and Diets
2.3
A total of 300 Arbour Acre broiler chicks, all unsexed, were separated into 5 treatment groups, each consisting of 60 chicks, at random. Five replications of each group were formed, each containing 12 chicks. The first group followed a control diet free of BNCo, whereas the second, third, fourth and fifth groups received diets fortified with BNCo at concentrations of 100, 200, 300 and 400 ppm, respectively. Composition and chemical analysis of the experimental diets (starter and finisher diets) are displayed in Table 1. BNCo has been characterized and provided by the Department of Agricultural Microbiology, Faculty of Agriculture, Zagazig University, Zagazig, Egypt.
Growth Performance and Carcass Traits
2.4
Live body weight (LBW) was recorded at 1, 3 and 6 weeks of age. All growth performance metrics of the birds were documented at three intervals (1–3, 1–6 and 3–6 weeks of age). At 42 days of age, we randomly chose 25 chickens to assess carcass traits. Each bird was weighed and then euthanized. The weight of the heart, liver and gizzard was calculated and stated as a percentage of the LBW before euthanasia.
Blood Metabolites
2.5
Five blood samples from each treatment were randomly taken into heparinized test tubes to assess the blood picture parameters: white blood cells (WBCs, 10^3^/µL), red blood cells (RBCs, 10^6^/µL) and haemoglobin (Hb, g/dL). To ascertain the blood biochemical parameters, the residual blood was subjected to centrifugation to obtain plasma. The biochemical components, including protein and its portions, aspartate aminotransferase (AST), alanine aminotransferase (ALT), renal function parameters such as creatinine, urea and uric acid, also triglycerides (TG), total cholesterol (TC) and its subtype as high‐density lipoprotein (HDL), low‐density lipoprotein (LDL) and very‐LDL (VLDL), and immunoglobulins fractions (IgY, IgA and IgM), were analysed using an automatic analyser with commercial kits from Bio‐diagnostic Company (Giza, Egypt) in accordance with the manufacturer's protocol. Superoxide dismutase (SOD), malondialdehyde (MDA), catalase (CAT), total antioxidant capacity (TAC) and reduced glutathione (GSH) levels in plasma were quantified using commercial kits from Biodiagnostic Company (Giza, Egypt).
Digestive Enzymes
2.6
The Somogyi (1960) technique was utilized to evaluate amylase activity, whereas the analysis of lipase enzyme adhered to the protocol established by Tietz and Fiereck (1966). The methodologies stated by Lynn and Clevette‐Radford (1984) were utilized to evaluate the action of the protease enzyme.
Statistics
2.7
Statistical analyses were conducted utilizing SAS software. The growth metrics, carcass traits, blood profile, blood parameters and digestive enzymes were applied using a one‐way analysis of variance (p < 0.05).
Results and Discussion
3
Growth Performance
3.1
The growth performances of broiler chicks as influenced by dietary supplementation with BNCo are illustrated in Table 2. The finding illustrates a significant increase in LBW at 3 and 6 weeks (p < 0.05, p < 0.01). The third group, treated with 200 ppm BNCo, showed increased results (912.33 and 2233.48 g). Moreover, body weight gain was significantly increased during the 3–6 and 1–6 weeks (p < 0.001, p < 0.0001), and the third group showed the most pronounced results (62.91 and 58.55 g). However, the feed intake showed no significant variations throughout the trial period: 1–3 weeks, 3–6 weeks, 1–6 weeks. Furthermore, the third group presented significantly better FCR during the 3–6 week period (p < 0.01) and the 1–6 week period (p < 0.01), with values of 1.97 and 1.78, respectively.
Most poultry studies have focused on the effects of supplementing with mineral nanoforms to promote growth and mineral retention (Hossain 2025; Nechitailo et al. 2025). Our result discovered a significant improvement (p < 0.05) in birds' body weight, body weight gain and feed conversion ratio, whereas feed intake was influenced non‐significantly. The improvement of BWG in the BNCo‐treated group may be due to the greater bioavailability of Co in the NPs. Our finding aligns with the data of Halle et al. (2011), who demonstrated that treatments with 0.65 mg Co enhanced feed consumption and, consequently, the growth of broilers. The FCR was influenced mainly during the initial 2 weeks by elevated concentrations of vitamin B12 and vitamin B12 combined with BNCo in the feed. With the same result, Ibrahim et al. (2017) demonstrated that the broilers given diets supplemented with nano‐ZnO (110 mg/kg) revealed improvement in BWG and FCR compared to those given diets treated with organic Zn and inorganic ones.
Furthermore, our findings agree with Mohammadi et al. (2015), who noted enhanced growth performance in chickens treated with nano‐zinc‐methionine and nano‐zinc‐max at 80 mg/kg of diet. Conversely, dietary nano‐zinc sulphate decreases growth performance in broiler chicken. Nano‐silver treatments at 4 mg/kg enhanced BWG and optimal FCR in broilers (Elkloub et al. 2015). Andi et al. (2011) confirmed that silver, which acts as an antimicrobial agent against harmful gut microbes, may promote gut health, leading to improved nutrient absorption, as demonstrated by increased body mass, feed intake and feed conversion rate in broiler chickens consuming feed with AgNP.
Carcass Characteristics
3.2
The carcass characteristics of broiler chicks influenced by dietary supplementation with BNCo are illustrated in Table 3. The data illustrated a non‐significant variation in carcass characteristics (carcass, liver, gizzard, heart, giblets and dressing) percentage. The second group showed an increased carcass and heart percentage (74.46% and 72%, respectively), whereas the fourth group presented increased liver, giblet and dressing percentages (2.61%, 6.58% and 80.77%), respectively.
The carcass parameters used in this study, which included the broiler's carcass, gizzard, heart, giblets and dressing percentage, did not significantly differ depending on the concentration of Nano‐Co treatments. The adoption of a good, balanced basal diet may be the reason for the lack of notable variations in carcass criteria. After a certain amount of BNCo, carcass features may no longer be improved. Research has demonstrated that the addition of 0.3–0.5 mg/kg of nano‐selenium to feed had no influence on the carcass characteristics and organ weights of broilers, which aligns with our findings (Bakhshalinejad et al. 2019; Ashour et al. 2025). Similarly, Wang et al. (2011) demonstrated that there were no appreciable changes in carcass criteria after supplementing with nano‐selenium at doses of 50–150–300 µg/kg. Moreover, our findings match with the results of Esfahani et al. (2015), who stated that the addition of zinc oxide NPs in a dry chicken diet enhanced carcass traits and the proportional weight of digestive and lymphoid organs in comparison to a wet diet at the starter (1–21) day period.
Haematology
3.3
The results showed a significant elevation in Hb and RBCs (p < 0.05, p < 0.01) in groups supplemented with BNCo, as explained in Table 4. The third group illustrated an elevated level of Hb (8.25 g/dL), whereas the fourth group presented increased numbers of RBCs (4.12 × 10^6^/µL) relative to the control. The BNCo groups presented a significant (p = 0.0159) rise in the number of WBCs. The finding reported a significant rise in Hb, RBCs and WBCs in the groups supplemented with BNCo. Our result agrees with Diaz et al. (1994), who stated an elevation in erythrocytes and Hb in broilers treated with a diet of cobalt. Conversely, Kato et al. (2003) reported that the haematocrit, Hb, erythrocytes and leukocyte counts were unaffected by cobalt and vitamin B12 treatment in the diet, whereas erythrocyte and Hb levels were unaffected by cobalt‐only treatment. Cobalt, as the primary cofactor of vitamin B12, is essential for nucleotide combination and can also activate the haematological system in bone marrow, hence enhancing Hb synthesis and increasing RBC production (Chen et al. 2021). The complicated methods by which cobalt stimulates haematopoiesis are summarized as follows: First, as mentioned earlier, cobalt is involved in the metabolism of RNA and blood components as the active centre of vitamin B12, affecting the haematopoietic process (Danzeisen et al. 2020). A lack of vitamin B12 can lead to megaloblastic anaemia because of the suppression of the synthesis of DNA due to reduced pyrimidine and purine availability, resulting in increased (RBCs) size and the increase of substantial, undeveloped precursor cells (megaloblasts) of RBCs in the bone marrow and blood (Tjong et al. 2022).
Liver and Kidney Functions
3.4
Hepatic and renal functions of broiler chicks as impacted by dietary supplementation with BNCo are shown in Table 5. The conclusions illustrated a significant rise in TP, ALB and GLOB (p < 0.001, p < 0.01, p < 0.01), and the third group revealed increased levels of TP and ALB (4.47, 2.61 g/dL), whereas the fourth group showed the increased GLOB level (1.96 g/dL) relative to control and other groups. Liver enzymes AST and ALT showed a significant variation (p < 0.001, p < 0.01) with BNCo treatments, and the third group revealed decreased levels (52.38, 15.30 IU/L) relative to control and other treatments. Kidney function tests (creatinine and urea) illustrated a significant improvement with BNCo treatments (p < 0.01, p < 0.05), and the third group revealed decreased levels of creatinine and urea (0.72 and 2.34 mg/dL). On the other hand, uric acid levels were not significantly affected by BNCo supplementation (p = 0.0953).
The liver function test significantly improved by increasing total protein, albumin and globulin in the blood serum of broiler chicks, whereas AST and ALT serum levels significantly decreased by BNCo supplementation. In the meantime, all treatments with BNCo levels showed a significant drop in serum liver function enzymes (ALT and AST) and renal function tests (creatinine, urea and uric acid). The improved liver and kidney functions seen in broilers may be a result of the complex effects of BNCo, which include antioxidant qualities and metabolic regulation. Our findings agreed with Shokraneh et al. (2020), who confirmed that nano‐selenium significantly reduced the concentrations of AST and ALT. Similarly, Qin et al. (2016) demonstrated that nano‐selenium dietary treatment significantly enhanced the liver and kidneys of rabbits. Notably, ALT and AST enzyme levels can be used to measure liver oxidative damage. In this investigation, feed supplementation with BNCo significantly decreased the serum concentration of creatinine, uric acid, urea, ALT and AST in chickens. These findings were corroborated by Sheiha et al. (2020), who found that the blood serum of Cobb chicken supplemented with nano‐selenium at doses of 0.15, 0.075 and 0.0375 ppm had lower levels of AST, ALT and creatinine. Accordingly, adding nano‐SE to the broiler diet may have contributed to the substantial drop in both ALT and AST, which was determined to be caused by nano‐selenium's major effects on fat metabolism through the thyroid (T3) hormone (Dalia et al. 2017).
Lipid Profile
3.5
Table 6 illustrates the lipid profile of broiler chicks influenced by feeding treatments with BNCo. The findings revealed a significant improvement in the lipid profile, and the third group demonstrated a significant (p < 0.05, p < 0.01) reduction in TC and LDL (150 and 77.74 mg/dL). Furthermore, the fourth group presented a significant (p < 0.05, p < 0.05, p < 0.05) decrease in TG, HDL and VLDL (56.50, 46.94 and 11.30 mg/dL).
According to this study, broiler lipid profile levels were affected by BNCo dietary supplementation, which reported a substantial drop (p < 0.05) in cholesterol, TG, LDL and VLDL at doses of 100, 200 and 300 ppm, respectively. Our findings align with earlier research, particularly that by Mohapatra et al. (2014), who confirmed that broilers given 0.3 mg/kg of nano‐selenium had substantially lower cholesterol and triglyceride concentrations than the control group. Similarly, Saleh and Ebeid (2019) confirmed that adding 0.5 mg/kg of dietary nano‐SE to broiler feeds significantly decreased plasma TG and TC.
Antioxidant Status
3.6
The antioxidant condition of broiler chicks as impacted by dietary supplementation with BNCo is illustrated in Table 7. The data presented a significant boost in antioxidant status with BNCo treatments. The second group presented a significant drop (p < 0.01) in MDA activity (0.22 nmol/mL) and a significant progress (p < 0.01) in SOD activity (0.58 U/mL). Furthermore, the third group showed a significant boost (p < 0.05, p < 0.01, p < 0.001) in CAT, TAC and GSH (0.52 mg/dL, 0.57 ng/mL, 0.72 mg/dL), respectively, compared to the control and other treatments.
The antioxidant status was improved significantly with BNCo supplementation. These outcomes align with Scott et al. (2016) who stated that CoNPs can dramatically boost the creation of lipid peroxidation, and ROS, and boost caspase‐3 enzymes to protect the cells from oxidation. In addition to their average TAC and total reducing impact, the BNCo established exceptional radical scavenging potential (Matinise et al. 2018). Shahzadi et al. (2019) also confirmed that bioinspired CoNPs have radical scavenging activity; they also noted that the antioxidant activity and scavenging power are dosage dependent, meaning that as activity rises, so does CoNP concentration. The green‐synthesized NPs exhibited significant free radical‐scavenging action and yielded remarkable results (Hou et al. 2020). Similarly, it has been observed that cobalt oxide NPs produced from Sesbania sesban extract have lower DPPH radical scavenging efficacy than silver and copper oxide NPs (Ghadi et al. 2018). Moreover, Elkhateeb et al. (2024) confirmed that MDA significantly dropped as nano‐Se concentration increased to 0.4 mg/kg, and the TAC provided a substantial linear rise with elevation of nano‐selenium concentration at 0.4 mg/kg. Moreover, Hafez et al. (2020) reported that broiler hens' antioxidant status was improved by adding zinc oxide NPs at 40 or 80 mg/kg to their feed, as shown by amplified SOD and CAT action and diminished MDA levels. Zhao et al. (2014) confirmed that the group administered 20 mg/kg nano‐ZnO exhibited a reduction in serum and liver MDA concentration, enhanced TAO action in both blood and liver tissue and elevated serum CAT action.
Additionally, Zhao et al. (2014) confirmed that feeds supplemented with 60 or 100 mg Zn/kg from ZnO NP had increased action of serum CAT and serum and liver SOD. MDA is typically employed as an oxidative stress indicator (Mateos et al. 2005). The lower MDA in our study following Zn supplementation was consistent with earlier research (Liu et al. 2015).
Immunity
3.7
The weight of immunological organs (spleen, thymus and bursa of Fabricius) influenced by BNCo treatments is presented in Table 8. The findings demonstrated a significant (p < 0.01, p < 0.05) increase in spleen and thymus gland weight percentage, and the third group presented an increased weight percentage (0.15% and 0.60%). In contrast, the second group presented a nonsignificant increase in the bursa of Fabricius weight percentage (0.34%) relative to the control and other treated groups.
The immune organs’ weight, especially the spleen, thymus and bursa Fabricius, significantly boosted (p < 0.05) with BNCo supplementation to the broilers’ diet. Our results agreed with Grodzik and Sawosz (2006), who confirmed the impact of silver NPs at 10 ppm in embryonic bursa Fabricius and development and found no discernible influence on chicken growth but a decrease in follicle size and quantity. Ahmadi et al. (2010) examined the shifts in the proportional weight of the bursa following the supplementation of Ag–NPs at 20, 40 and 60 ppm concentrations, and there were fewer and smaller follicles. This could be because AgNPs’ antibacterial qualities impact the gut's microbial populations. Ag–NPs undoubtedly slow the development of anaerobic microbes and transfer available oxygen. Therefore, the growth of the bursa of Fabricius is negatively impacted.
On the other hand, the destiny of Ag–NPs in the host may depend on their capacity to elicit an immunological response when they are recognized spontaneously or when immune recognition is absent. The inflammatory reaction by NPs may impact the immunological status and the stability of T helper 1 and T helper 2 cells. According to the findings of Matsumura et al. (2003), a similar impact of silver zeolite may be caused by Ag absorption into bacterial cells upon contact, which impairs cellular processes and causes cell destruction. However, it may be described by the production of reactive oxygen molecules, which prevent cells from functioning properly. It makes sense that microorganisms in the gastrointestinal system are required for the bursa to grow and mature in healthy broilers. The antiviral effectiveness of an Ag–NP solution against the virus that causes infectious bursal disease in chicken embryos was documented in another investigation (Pangestika and Ernawati 2017).
The immunological status of broiler chicks as influenced by dietary supplementation with BNCo is demonstrated in Table 9. The results presented a significant boost in immunity parameters, and the third group revealed a significant increase (p < 0.01, p < 0.05, p < 0.05) in IgA, IgY and lysozymes (0.72, 1.04 and 0.55 mg/dL) relative to the control and other treatments. The second group illustrated a significant boost (p < 0.001) in IgM (0.90 mg/dL) relative to control and other treatments. Furthermore, the second group presented a significant increase (p < 0.01) in complement 3 (52.58 mg/dL). Additionally, the third group presented a significant increase (p < 0.05) in lysozyme levels (0.55 mg/dL) compared to the control and other treatments.
The immunological status was positively affected by BNCo treatment, particularly for IgA, IgY and IgM; additionally, complement 3 and lysozyme were also positively affected. IgM and IgG levels do not seem to be impacted by Ag–NPs (Pineda et al. 2012). The immune system, which is essential to good health and connected to all physiological systems, guards the host against infections (Gao et al. 2014) and continuously monitors native cells that could be dangerous, such as tumour‐forming cells (Pan Na et al. 2011). According to reports, metals are crucially involved in controlling the host's defence against invasive pathogens as well as the innate immune system's perception of them (Anwar, Awais et al. 2019), indicating that metals are involved in controlling the immune system's resistance to infection. A recent study has provided a new medical approach that utilizes metal‐based composites for disease treatment, revealing that these compounds may regulate autophagy, a crucial host immune response (Sahoo et al. 2014).
Digestive Enzymes
3.8
The impact of dietary treatments with BNCo on the digestive enzymes of broiler chicks is presented in Table 10. The outcomes presented a significant enhancement in digestive enzymes with BNCo treatments, and the third group revealed a significant increase (p < 0.01, p < 0.001) in amylase and protease enzymes (182.10 and 3.17 μ/L) in relation to control and other treatments. The second group presented a significant increase (p < 0.01) in lipase enzyme (47.34 μ/L) compared to the control and other treatments.
The digestive enzymes, such as amylase, lipase and protease, significantly improved, and these discoveries align with the conclusions of Brugger et al. (2016), who illustrated that on Day 35, the HME‐Zn supplementation significantly raised the amylase activity (p < 0.05). In zinc metabolism, exocrine glands like the pancreatic bile regulate zinc homeostasis through the pancreatic digestive enzymes to be secreted by the GIT. Similar to this study, the activities of broiler pancreatic amylase, lipase and trypsin were elevated in diets treated with organic zinc sources that promoted zinc absorption (Hu et al. 2022). Nonetheless, Park et al. (2015) revealed that treatments containing 100, 2500 and 100 mg/kg of coated zinc oxide did not differ in their effects on amylase and the action of trypsin. Nevertheless, the chicks’ pancreas's amylase, lipase and trypsin activity significantly decreased as a result of the excessive additional zinc (2000 mg/kg) (Lü et al. 1988).
In conclusion, broiler chickens given BNCo supplements had better growth performance, carcass traits and antioxidant activity. The beneficial hypolipidemic effects were demonstrated by dietary BNCo supplementation, which reduced serum triglyceride and cholesterol levels. BNCo supplementation at 200 ppm/kg resulted in the greatest overall performance of broilers, suggesting that it could be a novel and useful addition to broiler diets. To better understand how BNCo works as an antioxidant, antibacterial and immune system enhancer in broilers' internal organs, as well as how it affects the sustainability of productive performance, further research is required.
Author Contributions
Fayiz M. Reda, Abdullah S. Alawam, Hemat K. Mahmoud, Mohamed T. El‐Saadony, Ayman S. Salah, Hassan A. Rudayni, Ahmed A. Allam, Karima El‐Naggar, Mahmoud Alagawany and Most Khairunnesa were involved in conceptualization, data curation, formal analysis, investigation, methodology, resources, validation, visualization, roles/writing of the original draft, writing review and editing.
Conflicts of Interest
The authors declare no conflicts of interest.
Peer Review
The peer review history for this article is available at https://publons.com/publon/10.1002/vms3.70508.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abd El‐Hack, M. E. , M. Alagawany , A. S. Salah , M. A. Abdel‐Latif , and M. F. Farghly . 2018. “Effects of Dietary Supplementation of Zinc Oxide and Zinc Methionine on Layer Performance, Egg Quality, and Blood Serum Indices.” Biological Trace Element Research 184: 456–462.29081062 10.1007/s 12011-017-1190-0 · doi ↗ · pubmed ↗
- 2Abdollahi, M. , J. Rezaei , and H. Fazaeli . 2020. “Performance, Rumen Fermentation, Blood Minerals, Leukocyte and Antioxidant Capacity of Young Holstein Calves Receiving High‐Surface Zn O Instead of Common Zn O.” Archives Animal Nutrition 74, no. 3: 189–205.10.1080/1745039 X.2019.169038931851525 · doi ↗ · pubmed ↗
- 3Ahmadi, F. , and A. H. Kurdestani . 2010. “The Impact of Silver Nanoparticles on Growth Performance, Lymphoid Organs and Oxidative Stress Indicators in Broiler Chicks.” Global Veterinaria 5: 366–370.
- 4AL‐Ruwad, S. H. , A. I. Attia , U. M. A. Monem , et al. 2024. “Dietary Supplementation With Copper Nanoparticles (Cu NP) Enhances Broiler Performance by Improving Growth, Immunity, Digestive Enzymes and Gut Microbiota.” Poultry Science 103: 104026.10.1016/j.psj.2024.104026 PMC 1133811739067121 · doi ↗ · pubmed ↗
- 5Amr, E. , S. M. Sallam , M. E. Abd El‐Hack , A. S. Salah , and M. Alagawany . 2025. “Nano Clay in Animal Diets: Properties, Structure, Applications, and Toxicity.” In Organic Feed Additives for Livestock, 139–147. Academic Press.
- 6Andi, M. A. , M. Hashemi , and F. Ahmadi . 2011. “Effects of Feed Type With/Without Nanosil on Cumulative Performance, Relative Organ Weight and Some Blood Parameters of Broilers.” Global Veterinaria 7: 605–609.
- 7Anwar, A. , L. Chi Fung , A. Anwar , et al. 2019. “Effects of Shape and Size of Cobalt Phosphate Nanoparticles Against Acanthamoeba castellani .” Pathogens 8, no. 4: 260.31766722 10.3390/pathogens 8040260 PMC 6963488 · doi ↗ · pubmed ↗
- 8Anwar, M. I. , M. M. Awais , M. Akhtar , M. T. Navid , and F. Muhammad . 2019. “Nutritional and Immunological Effects of Nano‐Particles in Commercial Poultry Birds.” World's Poultry Science Journal 75, no. 2: 261–272.