Overexpression of housekeeping gene FveIPT2 enhances anthocyanin and terpenoid accumulation in strawberry fruits with minimal impact on plant growth and development
Lijun Gan, Manman Wei, Shanqi Cao, Hui Zhang, Xuechun Wang, Mingjia Chen, Na Yang, Changhua Zhu, Yi Li

TL;DR
Overexpressing the FveIPT2 gene in strawberries boosts anthocyanin and terpenoid levels, improving fruit quality without affecting plant growth.
Contribution
Demonstrates that overexpression of a tRNA-type IPT gene can enhance fruit nutritional value with minimal developmental impact.
Findings
Transgenic strawberries showed increased anthocyanin and terpenoid accumulation.
Nine anthocyanins and 24 terpenoids were significantly elevated in overexpressing fruits.
FveIPT2 overexpression upregulated genes involved in anthocyanin and terpenoid biosynthesis.
Abstract
Anthocyanins and terpenoids are secondary metabolites with well-documented health benefits. Isopentenyl transferases (IPTs) are key enzymes in cytokinin (CK) biosynthesis. While ADP/ATP-type IPTs and their associated trans-zeatin (tZ)-CKs and iP-CKs are considered to play regulatory roles in growth and development, as well as stress acclimation in plants, tRNA-type IPTs and cis-zeatin CKs (cZ-CKs), which may serve housekeeping functions, remain less studied. In this study, the tRNA-type IPT gene FveIPT2 was overexpressed in woodland strawberries (Fragaria vesca). Overexpression had minimal impact on plant growth and CK levels but resulted in transgenic fruits exhibiting a significant increase in total phenolic, flavonoid, and anthocyanin contents, indicating enhanced fruit quality. Metabolite profiling revealed substantial increases in nine specific anthocyanins and 24 out of 47…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
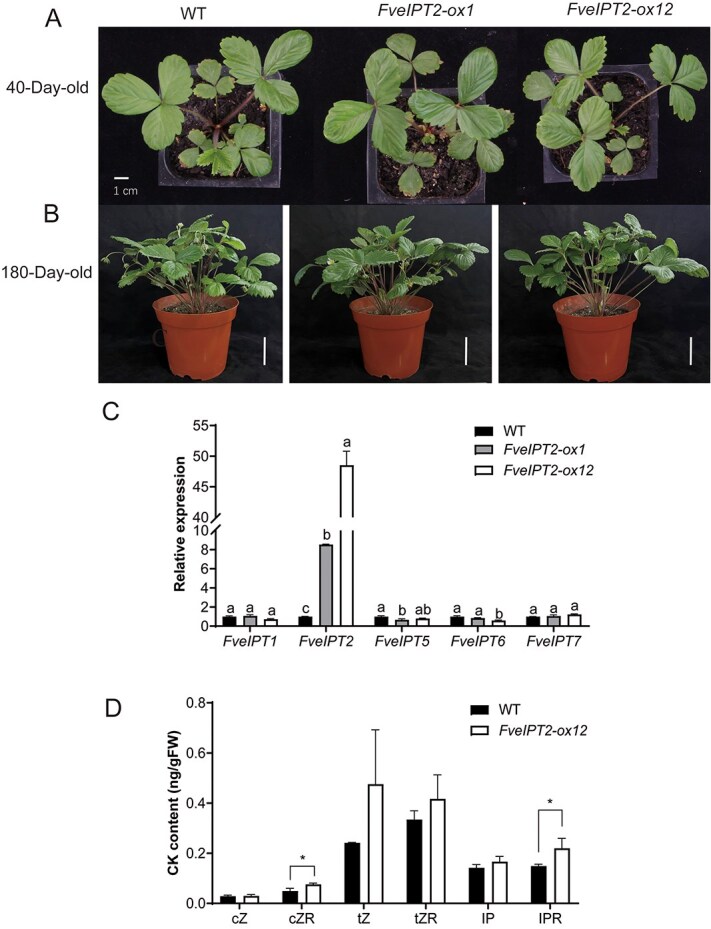 Figure 1
Figure 1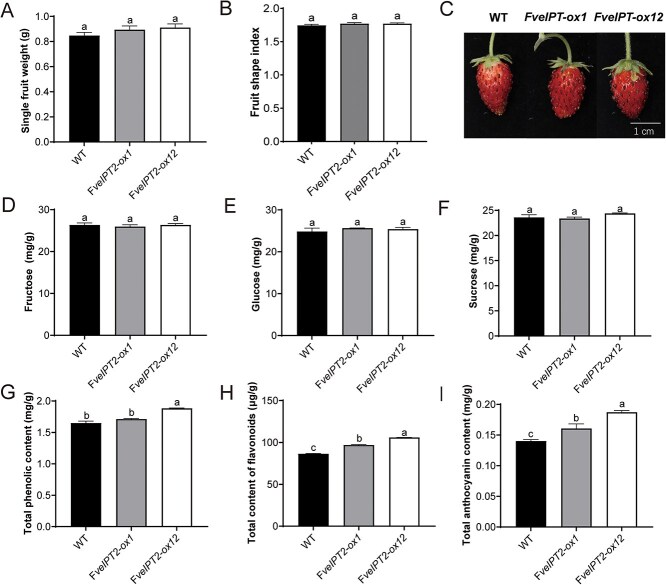 Figure 2
Figure 2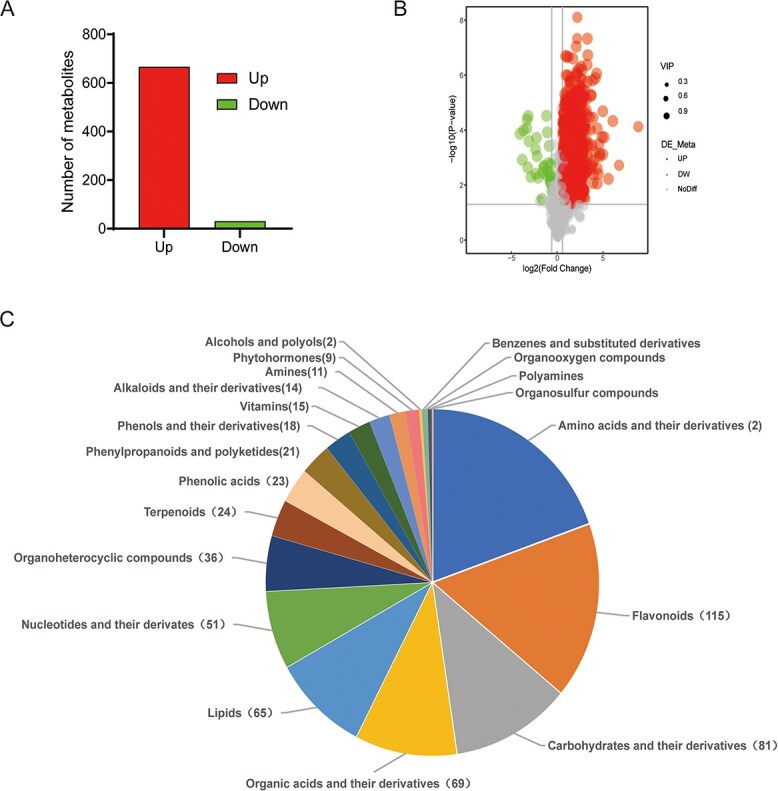 Figure 3
Figure 3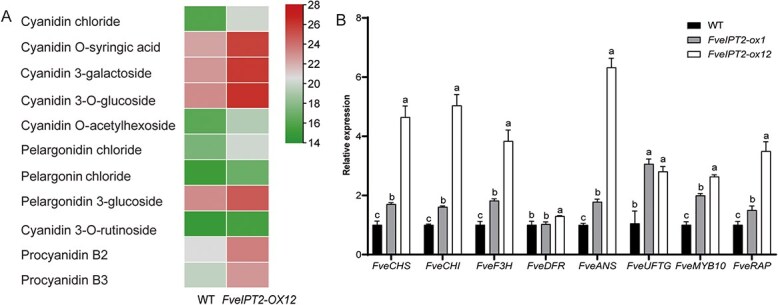 Figure 4
Figure 4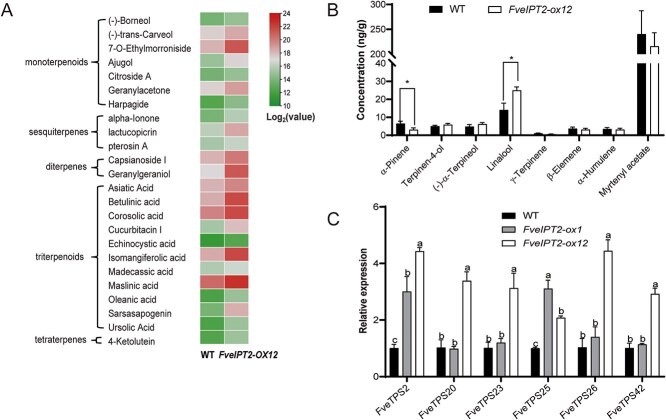 Figure 5
Figure 5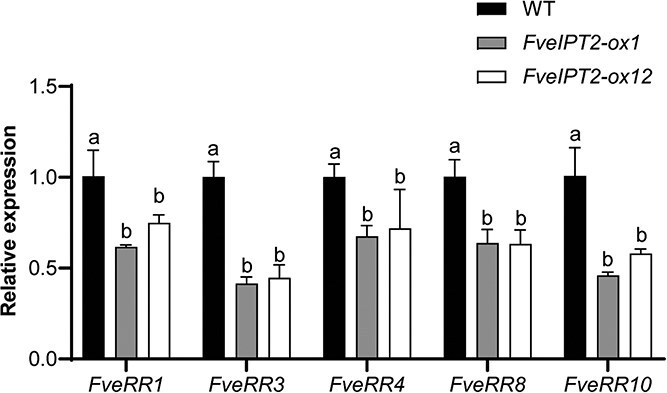 Figure 6
Figure 6- —the National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBerry genetics and cultivation research · Horticultural and Viticultural Research · Plant tissue culture and regeneration
Introduction
Cytokinins (CKs) are essential plant hormones that modulate numerous facets of plant growth and developmental processes. These functions include controlling apical dominance, promoting shoot and root development, regulating leaf senescence, and facilitating fruit formation [1–3]. In higher plants, the main types of CKs are zeatin (in trans and cis isomers, abbreviated as tZ and cZ) and isopentenyl adenine (iP), along with their riboside conjugates (tZR, cZR, iPR) [2, 4].
Isopentenyl transferases (IPTs), which attach an isopentenyl group to an adenosine molecule, catalyze the first committed step of CK production [2]. IPTs exist in two forms: adenylate-type IPTs (ATP/ADP-type IPTs) and transfer RNA-type IPTs (tRNA-type IPTs), with ATP/ADP and tRNA serving as their respective adenine substrates [4, 5]. A particular adenine at position 37 on the antiloop of tRNAs that identify codons starting with uracil is the target of tRNA-type IPTs [4]. In Arabidopsis, there are nine IPT genes (AtIPT1–9), with seven (AtIPT1, 3, 4–8) encoding ATP/ADP-type IPTs and the remaining two (AtIPT2 and AtIPT9) encoding tRNA-type IPTs [5]. The atipt1,3,5,7 quadruple mutant significantly decreases the levels of iP-CKs and tZ-CKs, demonstrating that ATP/ADP-type IPTs are responsible for generating both types of CKs [5]. Mutations in atipt2, atipt9, or the atipt2,9 double mutant exert negligible influence on tZ- or iP-CKs concentrations yet greatly impact cZ-CK levels [5, 6]. This implies that the bulk of cZ-CKs production likely stems from the degradation of prenylated tRNA [5].
Previous studies have established that tZ- and iP-CKs are the main biologically active CKs in plants, while cZ has shown lower activity [4, 7, 8]. However, ongoing research has uncovered significant levels of cZ-CKs in specific crops such as rice [9] and maize [10, 11], as well as during certain developmental stages associated with restricted growth [4, 11, 12]. Intriguingly, maize roots contain a much higher concentration of cZ-CKs compared to iP- and tZ-CKs combined [4, 10]. The reasons behind this elevated cZ-CKs content in these plants remain unknown. While the roles of tZ- and iP-CKs in plants have been extensively explored, the functions of cZ-type CKs remain less studied [4].
In our previous study [13], we demonstrated that tRNA-type IPTs are highly conserved, consistently retained, and constitutively expressed across diverse tissues and environmental conditions in angiosperms. In contrast, ATP/ADP-type IPTs have undergone significant expansion and functional divergence, exhibiting tissue- or organ-specific expression patterns and responsiveness to environmental stresses. Based on these findings, we hypothesized that tRNA-type IPT genes, along with their associated cZ-CKs, serve an essential housekeeping role in plants, while ATP/ADP-type IPT genes and their iP/tZ-CK counterparts are involved in regulatory functions related to growth and development, as well as stress acclimation in plants [13]. However, the functional characterization of cZ-CKs and tRNA-type IPT genes is still incomplete, requiring further experimental evidence to confirm this hypothesis.
Housekeeping genes typically support essential cellular functions rather than directly influencing growth, developmental timing, or stress responses. This raises the question: could overexpressing a housekeeping tRNA-type IPT gene impact specific metabolic pathways? Anthocyanins, the primary pigments in strawberry fruits [14], and terpenoids—essential secondary metabolites in plants [15, 16]—both contribute to fruit quality and nutritional value. These secondary metabolites are well known for their various health benefits [17–19]. Studies have shown that CK can induce anthocyanin accumulation across various plant species, including Arabidopsis [20], apple [21], strawberry [22], and Eucalyptus [23], and regulate terpenoid biosynthesis in Artemisia alba [24] and Thymus vulgaris [25]. Given the health benefits of anthocyanins and terpenoids, we explored whether manipulating a housekeeping IPT gene could enhance the production of anthocyanins and terpenoids.
The woodland strawberry (Fragaria vesca) has emerged as a valuable model plant for investigating fruit development in other members of the Rosaceae family. This is due to its short growth cycle, availability of its genome sequence, and ease of gene editing [14, 26–28]. In F. vesca, two genes, FveIPT2 and FveIPT7, belong to the tRNA-type IPT family, while five genes, FveIPT1 and FveIPT3–6, are classified as ATP/ADP-type IPTs [29]. To gain deeper insights into the role of tRNA-type IPT in fruit quality, we overexpressed FveIPT2 in woodland strawberries. Surprisingly, overexpression of FveIPT2 has minimal impact on the levels of CKs. However, it leads to a significant elevation of the concentrations of terpenoids and anthocyanins, which are secondary metabolites associated with fruit quality and pigmentation. These findings provide experimental evidence for the roles of tRNA-type IPT genes and enhance our understanding of the regulatory mechanisms underlying secondary metabolite synthesis in strawberry fruits.
Results
Overexpression of 35S::FveIPT2 led to a slight rise in cytokinin levels
To elucidate the function of the tRNA-type IPT gene, FveIPT2, it was overexpressed using the promoter of cauliflower mosaic virus (CaMV) 35S in F. vesca. Two independent transgenic lines, denoted as FveIPT2-ox1 and FveIPT2-ox12, were selected for further analysis. As shown in Fig. 1A and B, no apparent morphological changes were observed in any of the transgenic lines. The relative expression levels of FveIPT2 in 40-day-old seedlings between the transgenic lines and wild type (WT) were compared utilizing real-time quantitative polymerase chain reaction (RT-qPCR). The findings revealed a marked upregulation of FveIPT2 expression in FveIPT2-ox1 and FveIPT2-ox12, with expression levels 8.54 and 48.5 times higher than those in the WT, respectively. However, minor alterations were observed in the expression levels of other FveIPT genes (Fig. 1C). Owing to the previously reported low expression levels of FveIPT3 and FveIPT4, we were unable to measure their expression in seedlings [29].
Overexpression of 35S::FveIPT2 in strawberry has minimal impact on plant growth, morphology, and cytokinin levels. No noticeable effects were observed in the 35S::FveIPT2 transgenic plants (40-day-old seedlings were shown in A and 180-day-old seedlings were shown in B). WT: wild type; FveIPT2-ox1 and FveIPT2-ox12 are 35S::FveIPT2 transgenic plants. Bar = 5 cm in (B). (C) FveIPT2 was expressed at high levels in 40-day-old transgenic seedlings. Error bars are SD (n = 3). P < 0.05. (D) A slight elevation in the concentrations of certain CKs was observed in 40-day-old transgenic seedlings. Error bars are SD (n = 3). P < 0.05.
In order to assess the impact of FveIPT2 overexpression on CK levels, we measured CK levels in 40-day-old seedlings of the transgenic line FveIPT2-ox12 and WT using UHPLC-TQ-MS/MS (ultra-high-performance liquid chromatography coupled with triple quadrupole electrospray tandem mass spectrometry). The findings revealed that the overexpression of FveIPT2 resulted in a slight elevation in the concentrations of certain CKs such as cZR and iPR (Fig. 1D). These CKs are products of the degradation of prenylated tRNA.
Overexpression of 35S::FveIPT2 increased fruit quality
To investigate the impact of overexpression of 35S::FveIPT2 on fruit growth and development, we compared the vegetative growth phase (from seed germination to first flowering) and the duration from anthesis to fruit ripening between transgenic and WT plants. Both developmental stages showed no statistically significant differences between transgenic and WT plants (Supplementary Fig. S1). Furthermore, measurements of single fruit weight and fruit shape revealed no significant changes in these parameters in transgenic fruits compared with WT (Fig. 2A and B). However, transgenic fruits exhibited a slightly darker red coloration compared with WT fruits (Fig. 2C). Further analysis showed that the overexpression of 35S::FveIPT2 had no significant impact on the concentrations of soluble sugars, such as fructose, glucose, and sucrose. However, it led to a marked elevation in the accumulation of total anthocyanin, total phenolic, and total flavonoid contents, thereby enhancing fruit quality (Fig. 2D–I). For example, relative to WT, the total anthocyanin content in FveIPT2-ox1 and FveIPT2-ox12 was 1.15-fold and 1.34-fold, respectively (Fig. 2I).
Overexpression of 35S::FveIPT2 in strawberry enhanced fruit quality. Measurements of single fruit weight (A) and fruit shape index (B). (C) Fruit phenotype. Overexpression of FveIPT2 did not affect the levels of fructose (D), glucose (E) or sucrose (F). Overexpression of FveIPT2 elevated the levels of total phenolic (G), total flavonoid (H) and total anthocyanin (I). Error bars are SD (n = 3). P < 0.05.
Comprehensive metabolite profiling of red receptacles
To comprehensively analyze the changes in metabolites influenced by the overexpression of FveIPT2, the receptacles of FveIPT2-ox12 and WT at the red fruit stage were examined using the quasi-targeted metabolomics technique. The principal component analysis (PCA) plot demonstrated that FveIPT2-ox12 showed distinct separation in PC1 compared to the WT, indicating altered metabolite accumulation in the transgenic fruits (Supplementary Fig. S2A). In total, 1058 metabolites were identified in these samples (Supplementary Table S1), which were further categorized into 21 types including 47 terpenoids, 194 amino acids and their derivatives, 150 flavonoids, and other categories (Supplementary Fig. S2B).
Using specific criteria (VIP > 1.0, FC > 1.5 or FC < 0.667, and P-value <0.05), we identified a total of 697 differentially expressed metabolites (DEMs) when comparing FveIPT2-ox12 to the WT. Among these DEMs, 666 metabolites showed increased abundance, while 31 metabolites exhibited reduced levels (Supplementary Table S1; Fig. 3A and B). These compounds were classified as amino acids and their derivatives (136), flavonoids (115), carbohydrates and their derivatives (81), and others (Fig. 3C).
The levels of amino acids and their derivatives, and flavonoids were significantly altered in the FveIPT2-ox12 transgenic fruits. (A) 666 metabolites were increased and 31 metabolites were reduced in transgenic fruits. Red-stage fruits (without seeds) from both WT and FveIPT2-ox12 plants were used for quasi-targeted metabolomics analysis. The criteria used to identify DEMs were as follows: VIP > 1.0, FC > 1.5 or FC < 0.667 and P-value < 0.05. (B) Volcano plot of DEMs between FveIPT2-ox12 and WT. The size of each dot reflects the VIP value. (C) The classification of DEMs revealed substantial alterations in amino acids and their derivatives, as well as flavonoids, in the fruits of FveIPT2-ox12 compared to those of WT. The numbers in parentheses indicate the counts of DEMs.
Anthocyanins were significantly increased in the transgenic fruits
Flavonoids constitute one of the largest categories of secondary metabolites found in fruits. They not only contribute to the flavor and sensory properties of fruits but also serve as powerful antioxidants in the human diet [30]. Anthocyanins, a subclass of water-soluble flavonoids, are particularly important for determining the color, fragrance, and taste profile of fruits [14, 31]. In our metabolomics study, we observed significant changes in 115 flavonoid compounds in FveIPT2 transgenic fruits (Fig. 3C). Out of these, nine anthocyanins exhibited a substantial increase in abundance in FveIPT2-ox12 fruits compared to the WT. These included cyanidin chloride (18.2-fold), cyanidin O-syringic acid (9.96-fold), cyanidin 3-galactoside (9.57-fold), cyanidin 3-O-glucoside (9.16-fold), cyanidin O-acetylhexoside (8.22-fold), pelargonidin chloride (6.82-fold), pelargonin chloride (3.25-fold), pelargonidin 3-glucoside (3.27-fold), and cyanidin 3-O-rutinoside (1.52-fold) (Fig. 4A). Additionally, two procyanidins (procyanidin B2 and procyanidin B3) were also found to be elevated in fruits from FveIPT2-ox12 (Fig. 4A).
Overexpression of 35S::FveIPT2 in strawberry enhanced anthocyanin production. (A)Anthocyanin production was enhanced in transgenic fruits. Red-stage fruits (without seeds) from both WT and FveIPT2-ox12 plants were subjected to quasi-targeted metabolomics analysis. Each value in the heatmap represents the log2-transformed original value of the anthocyanins. (B) The expression levels of anthocyanin-related genes were elevated in fruits from transgenic plants. Pre-turning stage fruits (without seeds) were used for RT-qPCR analysis, with FveCHC serving as the internal reference. Error bars are SD (n = 3). P < 0.05.
To further explore the effect of FveIPT2 overexpression on anthocyanin accumulation, receptacles at the pre-turning stage were used to analyze the expression of genes related to anthocyanin production, as this is the stage when anthocyanin accumulation initiates [32]. Our RT-qPCR data, in line with the metabolic findings, demonstrated significantly elevated expression levels of several genes associated with the pathways of anthocyanin biosynthesis and transport in the transgenic fruits compared to the WT. Key anthocyanin structural genes such as FveCHS, FveCHI, FveF3H, FveANS, and FveUFTG were notably upregulated in the transgenic fruits (Fig. 4B). FveRAP performs a crucial role in anthocyanin transport in strawberry fruits [33, 34]. The expression levels of FveRAP in FveIPT2-ox1 and FveIPT2-ox12 were 1.5 and 3.5 times higher, respectively, than those in the WT (Fig. 4B). Furthermore, the MYB transcription factor FveMYB10, known for its pivotal role in regulating anthocyanin production in fruits [35], showed increased expression in the transgenic fruits (Fig. 4B).
Overexpression of 35S::FveIPT2 in strawberry enhanced terpenoid accumulation in fruits
Compared with the WT, FveIPT2-ox12 fruits exhibited significantly higher levels of terpenoids. Among the 47 terpenoids detected in these samples, 24 terpenoids showed a marked elevation in the fruits of FveIPT2-ox12, including 11 triterpenoids, 2 diterpenoids, 3 sesquiterpenoids, 7 monoterpenoids, and 1 tetraterpene (Fig. 5A). The top five terpenoids with the highest increase in FveIPT2-ox12 were identified as sarsasapogenin, geranylgeraniol, betulinic acid, isomangiferolic acid, and 7-o-ethylmorroniside. Their levels in FveIPT2-ox12 were 25.55, 16.86, 10.12, 9.83, and 8.19 times higher than those in the WT, respectively (Fig. 5A).
Overexpression of 35S::FveIPT2 in strawberry elevated terpenoid accumulation in fruits. (A) Terpenoid production was elevated in transgenic fruits. Red-stage fruits (without seeds) from both WT and FveIPT2-ox12 plants were subjected for quasi-targeted metabolomics analysis. Each value in the heatmap represents the log2-transformed original value of terpenoids. (B) Linalool content was elevated, while α-pinene was decreased in FveIPT2-ox12 fruits. The volatile terpenoid contents in red fruits of both FveIPT2-ox12 and WT were detected by GC–MS. P < 0.05. (C) The expression levels of several genes associated with terpenoid production were significantly altered in transgenic fruits. Pre-turning stage fruits (without seeds) were used for RT-qPCR analysis, with FveCHC serving as the internal reference. Error bars are SD (n = 3). P < 0.05.
Volatile terpenoids represent the largest family of plant volatile compounds. Therefore, we analyzed the concentrations of volatile terpenes in the receptacles of both FveIPT2-ox12 and WT. The results showed that FveIPT2-ox12 had a significantly lower amount of α-pinene compared to the WT. In contrast, FveIPT2-ox12 exhibited a higher level of linalool than the WT (Fig. 5B). Terpinene-4-ol, (−)-α-terpineol, γ-terpinene, β-elemene, α-humulene, and myrtenyl acetate were also detected in strawberry receptacles, but their concentrations did not show significant alterations in the fruits of FveIPT2-ox12 (Fig. 5B).
Terpene synthase (TPS) enzymes catalyze the key step in the conversion of terpenoid precursors into various terpenoid compounds [36, 37]. We analyzed the expression of several genes encoding TPS that have been previously reported to be highly expressed in strawberry receptacles [37]. We observed that these TPS genes were significantly upregulated in the fruits of FveIPT2-ox12 compared with the WT (Fig. 5C).
Type-A response regulator genes were significantly downregulated in transgenic fruits
Type-A response regulator (ARR) genes, which are rapidly induced by CK and serve as marker genes for CK signaling pathway [2, 7], were measured to investigate whether the increased levels of anthocyanins and terpenoids in fruits overexpressing FveIPT2 are related to the activation of CK signaling. We measured the expression levels of three ARR genes, namely FveRR1, FveRR3, and FveRR4, in the receptacles. Interestingly, these marker genes were significantly downregulated in transgenic fruits. Additionally, two type-B RR genes, namely FveRR8 and FveRR10, were also downregulated in the transgenic fruits (Fig. 6). These findings indicate that the increased contents of anthocyanins and terpenoids in FveIPT2 overexpressing fruits may not be directly associated with type-A RR-mediated CK signaling pathway.
Altered expression levels of CK-regulated genes in 35S::FveIPT2 transgenic fruits. CK-responsive genes were downregulated in transgenic fruits. Pre-turning stage fruits (without seeds) were used for RT-qPCR analysis, with FveCHC serving as the internal reference. Error bars are SD (n = 3). P < 0.05.
Discussion
Housekeeping genes are characterized as those that exhibit consistent expression across various tissues, are essential for fundamental cellular maintenance functions, and are evolutionarily conserved across diverse species [38]. Previously, we proposed that tRNA-type IPT genes serve a fundamental housekeeping role in plants, while ATP/ADP-type IPTs are involved in regulatory functions related to growth and development, as well as stress acclimation in plants [13]. Further experimental validation is required to substantiate this hypothesis. While the phenotypic effects of manipulating ATP/ADP-type IPTs have been widely studied, the effects of altering tRNA-type IPTs, particularly through overexpression, remain largely unexplored [4].
In this study, we overexpressed FveIPT2 in woodland strawberries using the 35S CaMV promoter. Although FveIPT2 expression was significantly elevated in transgenic plants compared with the WT, this overexpression led to merely a slight elevation in the overall concentrations of cZR and iPR in young seedlings (Fig. 1). However, because our current method measures total CK levels across all analyzed tissue types, we cannot rule out the potential for significant changes in CK levels within specific tissues. Recently, the knockout of two tRNA-type IPT genes, atipt2 and atipt9, in Arabidopsis revealed distinct effects on cZ-CKs levels in siliques: the atipt2 mutant exhibited reduced total cZ-CKs, while the atipt9 mutant showed increased levels compared with the WT [6]. These findings suggest that cZ-CKs synthesis in plants is a complex, finely regulated process, likely influenced by tissue- and organ-specific factors that affect post-transcriptional regulation.
In tomato, constitutive expression of SIIPT3, an adenylate-type IPT gene, resulted in the overproduction of CKs (levels increased up to 12-fold) and significant alterations in plant phenotype such as dwarfism, and branchy and ligneous stems [39]. Similar effects were reported in Arabidopsis when an adenylate-type IPT gene, ZmIPT2, was overexpressed [40]. However, in the present study, the overexpression of FveIPT2 in strawberry had minimal impact on plant growth, morphology, and CK levels (Fig. 1). The lack of phenotypic changes associated with FveIPT2 overexpression stands in stark contrast to the significant morphological alterations typically observed when adenylate-type IPT genes are overexpressed [39, 40]. Housekeeping genes are defined by their essential roles in maintaining basic cellular functions. They are consistently expressed across all cell types under both normal and stress conditions, rather than being involved in the regulation of developmental processes or environmental responses [38]. The unchanged morphology, developmental progression, and growth patterns in FveIPT2-overexpressing plants further support the hypothesis that tRNA-type IPT genes function as housekeeping genes in higher plants [13].
Despite minimal effects on CK levels and plant growth, FveIPT2 overexpression significantly elevated anthocyanin content in transgenic fruits. Nine anthocyanins, including six cyanidin derivatives and three pelargonidin derivatives, accumulated at higher levels in transgenic fruits accompanied by upregulated expression of biosynthetic (FveCHS, FveCHI, FveF3H, FveANS, and FveUFTG), regulatory (FveMYB10), and transport-related (FveRAP) genes (Fig. 4). Similarly, various terpenoids, including monoterpenoids, sesquiterpenoids, diterpenoids, triterpenoids, and tetraterpenoids, were elevated in the transgenic fruits, with altered expression of TPS genes in FveIPT2 overexpression plants (Fig. 5A). Linalool, which is one of the most important terpenoids in strawberries, possesses a sweet, floral, and citrus-like aroma that enhances the fruit’s fragrance [41]. On the other hand, the olefinic monoterpene α-pinene contributes to the turpentine-like, woody, resinous, and unpleasant odor in strawberries [41]. Interestingly, in the present study, a notable reduction in α-pinene concentration and an elevation in linalool content in the fruits of FveIPT2-ox12 were observed (Fig. 5B), indicating that the overexpression of the FveIPT2 gene improves the scent of strawberry fruits.
The inductive effect of FveIPT2 overexpression on anthocyanin and terpenoid accumulation parallels studies in Arabidopsis and Thymus vulgaris L, where CKs modulate these metabolites [20, 24, 42, 43]. We explored the underlying mechanisms responsible for the observed metabolic changes. Our analysis of CK-responsive gene expression revealed a marked downregulation of canonical type-A RR-mediated signaling in transgenic plants (Fig. 6). This suggests that traditional CK signaling pathways are unlikely to be the primary drivers of the increased anthocyanin and terpenoid accumulation.
tRNA-IPT catalyzes the prenylation of adenosine-37 within tRNAs, which leads to cZ-CK production through the degradation of prenylated tRNAs [6, 44]. In Arabidopsis, mutations of tRNA-type IPT genes (AtIPT2 and AtIPT9) resulted in developmental defects such as smaller meristem size and reduced primary root elongation, yet exogenous cZ fails to rescue these phenotypes, implying direct roles for tRNA modification beyond cZ biosynthesis [45]. It is also possible that externally applied cZ does not effectively reach the tissues or cells essential for these phenotypes. Further research is needed to explore whether the enhanced fruit quality observed in FveIPT2-overexpressing transgenic plants is linked to tRNA modifications.
In conclusion, our study reinforces the housekeeping role of the tRNA-type IPT gene FveIPT2 and demonstrate that its overexpression enhances strawberry fruit quality without compromising growth. The metabolic improvements, mediated through upregulated anthocyanin and terpenoid pathways, highlight FveIPT2 as a promising target for the genetic improvement of strawberry fruit quality by increasing health-promoting secondary metabolites. Future studies are needed to elucidate the underlying molecular mechanisms.
Materials and methods
Plant materials and growth conditions
The woodland strawberry (F. vesca) accession ‘Ruegen’ was cultivated in a controlled greenhouse at Nanjing Agricultural University, China, under fluorescent lighting at an intensity of 120 μmol·m^−2^·s^−1^. The environment was controlled at a temperature of 22 ± 2°C, with a 16-h photoperiod and 60% relative humidity. For fruit phenotype analysis, flowers from plants of the same age that bloomed on the same day were selected for labeling and manual pollination to ensure consistency. There were no significant differences between transgenic and WT plants in the time from flowering to fruit ripening (Supplementary Fig. S1); therefore, fruits were harvested 30 days postpollination for analysis.
Stable transformation of strawberry
To construct the FveIPT2 overexpression vector, the complete coding sequence (CDS) of FveIPT2 (FvH4_3g29650) was cloned into the 2300GNR vector using specific primers (Supplementary Table S2), driven by the CaMV35S promoter. The construct was transformed into‘Ruegen’ via Agrobacterium (GV301)-mediated transformation performed following the previously described method [33]. Leaf explants were used for transformation, with putative transgenic calli selected through antibiotic resistance screening. Genotypic analysis was performed via PCR using a forward primer specific to the 35S promoter and a reverse primer targeting the coding sequence (Table S2), followed by Sanger sequencing confirmation. T3 plants were used for further analysis because this also demonstrates that the observed phenotypes are heritable and stable after multiple generations of sexual reproduction.
Extraction, purification, and measurement of CKs
For the analysis of CKs, 40-day-old seedlings from both WT and transgenic plants were harvested. CKs were extracted with 2-propanol/water/concentrated HCl (200:100:0.2, v/v/v). Following 12 h of shaking at 100 rpm (4°C), dichloromethane was added, and the mixture was shaken again. After phase separation, the lower organic phase was filtered through a 0.45-μm membrane. The samples were then dried, redissolved in 200 μl methanol and 200 μl water, and centrifuged (12 000 rpm, 10 min, 4°C). The supernatant was filtered through a 0.22-μm membrane and analyzed using an Agilent 1290 UPLC system (Agilent, USA) coupled with an AB SCIEX QTRAP® 6500 mass spectrometer (Applied Biosystems, USA), following the procedures outlined in our previous study [1].
Assessment of single fruit weight and fruit size
The weight of fruits was determined using a precision balance. The length and diameter of fruits were assessed using Vernier calipers and the fruit shape index was derived by dividing the fruit length by its diameter.
Measurement of total anthocyanin, flavonoid, phenolic, and soluble sugars
The total anthocyanin content was quantified using the pH differential method, in accordance with the protocol established by Tonutare et al. [46]. The total flavonoid was analyzed using colorimetry described by Xie et al. [47] and rutin was used to generate a standard curve. The total phenolic content was assessed according to the methodology described by Yan et al. [48]. The results were calculated by a standard curve of gallic acid. The quantification of soluble sugars was performed using HPLC as described by Gu et al. [32]. For each analysis, at least 10 fruits at the red stage, excluding seeds, were combined to create a single biological replicate. Three biological replicates were utilized for the analysis.
Volatile compounds analysis
To measure volatile compounds, samples were prepared using headspace solid-phase microextraction (HS-SPME), and the volatile profiles were characterized by gas chromatography–mass spectrometry (GC–MS) as described by Negri et al. [49]. 3-octanol was added to each sample as an internal standard composition. Volatile terpenoids were identified by cross-referencing mass spectra and retention times with both the NIST library (National Institute of Standards and Technology, 2014) and the Human Metabolome Database (HMDB). For each analysis, at least 10 fruits, excluding seeds, were combined to create an individual biological replicate. Three biological replicates were utilized for the analysis.
RNA extraction and RT-qPCR
Total RNA was isolated from the samples using the cetyltrimethylammonium bromide (CTAB) method, in accordance with the protocol established by White et al. [50]. Subsequently, RT-qPCR was conducted following the manufacturer’s guidelines using the extracted RNA. Primers used for RT-qPCR are detailed in Supplementary Table S2.
Quasi-targeted metabolomics analyses
To analyze the metabolomics profile, fruits (without seeds) at the red stage from both WT and OE12 plants were sent to Novogene Bioinformatics Technology Co., Ltd. (Beijing, China). The analysis of quasi-targeted metabolomics was performed following the methodology reported by Jiang et al. [51]. The LC–MS/MS analyses were conducted utilizing an ExionLC™ AD system (SCIEX) in conjunction with a QTRAP® 6500+ mass spectrometer (SCIEX). Metabolite profiling was performed using Multiple Reaction Monitoring (MRM) based on the Novogene in-house database. Metabolite identification was performed using Q1 and Q3 ions, along with retention time, declustering potential, and collision energy. Quantification was conducted using Q3. Raw data were processed with SCIEX OS software (v1.4) for peak integration and correction. Metabolites were annotated using the Kyoto Encyclopedia of Genes and Genomes (KEGG), Human Metabolome Database (HMDB), and LIPID MAPS (https://www.kegg.jp/, https://hmdb.ca/, https://www.lipidmaps.org/). The criteria used to identify DEMs were as follows: VIP (Variable Importance in Projection) score >1.0, fold change >1.5 or <0.667, and a P-value < 0.05. Three biological replicates were utilized for the analysis.
Statistical analyses
All analyses in this report were performed on three independent experiments. Statistical analyses were carried out with GraphPad Prism 8.0.1. (GraphPad, San Diego, CA, USA). Differences between treatments were tested using Duncan’s multiple range test for analysis of variance (ANOVA) with a significant level of P < 0.05. Different letters were used to indicate significant differences. Alternatively, a Student’s t-test was employed for specific comparisons.
Supplementary Material
Web_Material_uhaf130
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gan L, Song M, Wang X. et al. Cytokinins is involved in regulation of tomato pericarp thickness and fruit size. Hortic Res. 2022;9:uhab 04135043193 10.1093/hr/uhab 041PMC 8968492 · doi ↗ · pubmed ↗
- 2Kieber JJ, Schaller GE. Cytokinins. Arabidopsis Book. 2014;12:e 016824465173 10.1199/tab.0168 PMC 3894907 · doi ↗ · pubmed ↗
- 3Waldie T, Leyser O. Cytokinin targets auxin transport to promote shoot branching. Plant Physiol. 2018;177:803–1829717021 10.1104/pp.17.01691 PMC 6001322 · doi ↗ · pubmed ↗
- 4Schäfer M, Brütting C, Meza-Canales ID. et al. The role of cis-zeatin-type cytokinins in plant growth regulation and mediating responses to environmental interactions. J Exp Bot. 2015;66:4873–8425998904 10.1093/jxb/erv 214PMC 5147713 · doi ↗ · pubmed ↗
- 5Miyawaki K, Tarkowski P, Matsumoto-Kitano M. et al. Roles of Arabidopsis ATP/ADP isopentenyltransferases and t RNA isopentenyltransferases in cytokinin biosynthesis. Proc Natl Acad Sci USA. 2006;103:16598–60317062755 10.1073/pnas.0603522103 PMC 1637627 · doi ↗ · pubmed ↗
- 6Nguyen H, Butler C, Palberg D. et al. The t RNA-degradation pathway impacts the phenotype and metabolome of Arabidopsis thaliana: evidence from atipt 2 and atipt 9 knockout mutants. Plant Growth Regul. 2024;102:179–98
- 7Kieber JJ, Schaller GE. Cytokinin signaling in plant development. Development. 2018;145:dev 14934429487105 10.1242/dev.149344 · doi ↗ · pubmed ↗
- 8Silva-Navas J, Conesa CM, Saez A. et al. Role of cis-zeatin in root responses to phosphate starvation. New Phytol. 2019;224:242–5731230346 10.1111/nph.16020 · doi ↗ · pubmed ↗