First molecular identification of Cryptosporidium species isolated from canal water bodies in Minya Al-Qamh district, Northern Egypt
Marwa Omar, Samia E. Etewa, Tahani I. Farag, Samar Abd EL-Nabi, Heba Abdelal

TL;DR
This study found Cryptosporidium parvum in canal water in Egypt, highlighting a potential health risk due to poor water quality.
Contribution
First molecular identification of Cryptosporidium in canal water from Sharqia Province, Egypt.
Findings
Only 1.4% of water samples tested positive for Cryptosporidium.
The identified species was Cryptosporidium parvum based on COWP gene sequencing.
Contamination likely originates from human or animal sources.
Abstract
This study aimed to assess the prevalence of Cryptosporidium genotypes in the canal water bodies of Minya Al-Qamh District in Sharqia Governorate, Northern Egypt. Rural populations in Egypt lack access to clean water. They obtain their water supplies from different drains and canals, which are frequently exposed to contamination by human activities and untreated agricultural waste, introducing Cryptosporidium infection to unprotected waterways. A total of (72) canal water samples served for the molecular detection of Cryptosporidium species by PCR amplification and sequencing. Only one sample 1.4% (1/72) belonging to the village of Al-Aziziyyah was PCR-positive for Cryptosporidium contamination. Based on the (COWP) gene sequencing, this species was identified as Cryptosporidium parvum (C. parvum). The current work’s findings marked the first report on the molecular characterization of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2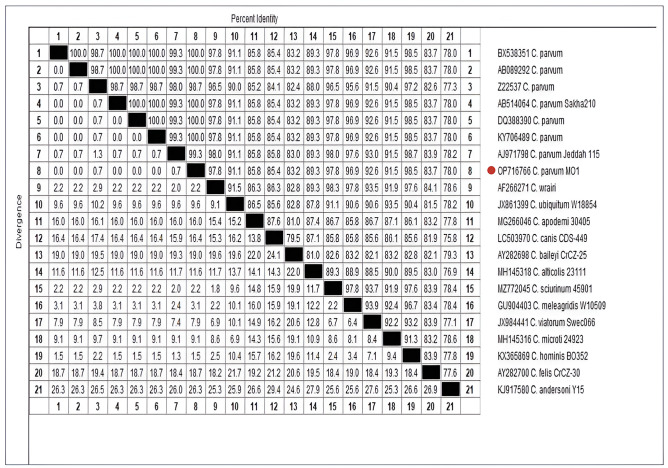 Figure 3
Figure 3- —Zagazig University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasitic Infections and Diagnostics · Amoebic Infections and Treatments · Mycobacterium research and diagnosis
Introduction
Safe and clean water is critical to good health and life sustainability. Around 1.1 billion people worldwide lack access to improved water supplies. One of the most common risks associated with water is its contamination by pathogenic microorganisms, including bacteria, viruses and protozoa [1]. Cryptosporidiosis, caused by a parasitic protozoan of the genus Cryptosporidium, is a major water-borne diarrheal disease in both humans and animals. Contaminated water constitutes a vital transmission route for Cryptosporidium infection. Between 2004 and 2010, Cryptosporidium was the causative agent of nearly 60.3% of the water-borne protozoan outbreaks recorded globally [2].
The significance of Cryptosporidium as a water-borne pathogen is based on its generalized presence in the environment, low infective threshold, and extreme resistance to conventional water disinfection practices such as chlorination. The thick-walled Cryptosporidium oocysts are highly stable and can remain infective outside the mammalian host for prolonged periods. They can also cross the physical barriers used to remove contaminants. The parasite defies water and health authorities by its ability to withstand chlorine disinfection and filtration. All these factors contribute to environmental contamination, enhancing water-borne parasitic transmission [1, 2].
Global climate change affects water quality through the gradual elevation of temperature and the extreme changes in seasonal patterns [3]. Due to its environmentally mediated life cycle, Cryptosporidium protozoan is particularly sensitive to these changes [4]. The summer peak of cryptosporidiosis could be attributed to several ecological factors, including warm temperature, humidity, and stagnation of water. These factors promote parasitic growth and prolong the infective period. Furthermore, increased human outdoor activities during the summer could potentially enhance the transmission cycle of the parasite [5].
Cryptosporidium is not only a human pathogen. It also affects a wide range of wild and domestic animals. Thus, the transmission is sustained by both anthroponotic and zoonotic cycles [1]. Amongst the species frequently found in water are Cryptosporidium parvum (C. parvum),* C. andersoni*, C. hominis, C. canis, and C. felis. However, C. parvum and C. hominis are the dominant genotypes, which account for most global human infections. C hominis primarily infects humans, while C. parvum has a wide range of hosts, particularly domestic livestock [6].
Egypt is one of middle-income countries with poor resource settings and limited diagnostic facilities. Hence, the choice of an optimal applicable diagnostic tool for cryptosporidiosis can be challenging. It is determined by the cost, affordability and availability of equipment and experienced personnel [7]. As traditional techniques fail to provide any information on the species or genotypes of Cryptosporidium present in samples, molecular tools based on the amplification of the parasite DNA have emerged. Molecular identification of Cryptosporidium to the species/genotype level has enabled the tracing of the contamination sources through their association with the type of host they parasitize [6]. In this context, this work was conducted to investigate the prevalence and molecular characterization of Cryptosporidium species isolated from the drainage canals in Minya Al-Qamh District, which could serve as a potential source for cryptosporidiosis in Sharqia Governorate, Northern Egypt.
Main text
Methods
Study area
The current cross-sectional study was conducted in the district of Minya Al-Qamh, Sharqia Governorate, Northern Egypt. Sharqia Governorate is located on the eastern branch of the Nile Delta, about 80 km northeast of Cairo. It covers approximately 4,911 km² and comprises numerous administrative centres, cities, and towns [8]. Minya Al-Qamh District was chosen to represent the rural community of the governorate, where networks of extended drains and canals serve as an irrigation system for agriculture. The canal water samples included in the study were obtained from four villages in the district, namely Al-Aziziyyah, Al-Maymounah, Kafr-Shalshalamun, and Beshet-Amer (Fig. 1).
Fig. 1. Geographical locations of the sampling villages. Water samples were collected from different canals distributed across four villages (Al-Aziziyyah, Al-Maymounah, Kafr-Shalshalamun, and Beshet-Amer), all located in Minya Al-Qamh District, Northern Egypt
Sample collection and processing
Seventy-two (72) raw water samples were obtained from 4 villages representing Minya Al-Qamh District (18 samples/village). Three samples were gathered from 6 different points in each area for better coverage. The source of all water samples was the drainage canals passing through the selected villages. The samples, collected in separate clean containers (5 L/sample), were carefully transported to the Medical Parasitology Department (Post-Graduate Research Laboratory), Faculty of Medicine, Zagazig University. Each sample was processed according to the methodology implemented by Omar et al. [7]. Briefly, after the application of the mesh sieve to remove coarse particles, each water sample was filtered through a Whatman^®^ nitrocellulose membrane filter (0.45 μm pore size and 110 mm in diameter, Cytiva, Marlborough, MA, USA). The membrane filter was next eluted with phosphate-buffered saline (PBS) in a 50 ml conical centrifugation tube. Each elute was concentrated by centrifugation at 6000 g for 10 min. After decanting the supernatant fluid, the recovered sedimentation pellets containing potential parasitic material were kept at ˗20 °C for further molecular analysis.
DNA extraction
The total genomic DNA of each sample was extracted using the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The extraction protocol was adapted for processing water samples, as previously described by Helmi et al. [9]. In brief, a volume of (200 µl) of the purified sample was added to a (1.5) ml microcentrifuge tube and homogenizd with (180 µl) of the lysis buffer (buffer ATL). The mixture was digested with proteinase K (20 µl) at 56 °C overnight, then incubated in (200 µl) buffer AL at 72˚C for 10 min. The mixture was carefully applied to the QIAamp mini spin column, and centrifugated at 8000 rpm for 1 min after adding (100 µl) buffer AE. To monitor PCR inhibition in environmental samples, both Chelex 100 (Sigma-Aldrich, Saint Louis, MO, USA) and Polyvinylpyrrolidone (PVP 360) (Sigma-Aldrich, Saint Louis, MO, USA) were added to the lysis buffer (ATL) at final concentrations of 20% and 2% respectively. The DNA extracts were stored at ˗20 °C for further PCR testing.
Identification of Cryptosporidium species
Water samples were genotyped with conventional polymerase chain reaction (PCR) followed by DNA sequencing. To identify the Cryptosporidium species, 553 bp of the molecular marker, Cryptosporidium oocyst wall protein (COWP) gene, was used. A partial sequence of the gene was amplified with the primers Cry-9: GGACTGAAATACAGGCATTATCTTG and Cry-15: GTAGATAATGGAAGAGATTGTG, as previously described by Feltus et al. [10]. Amplification was conducted in a Veriti 96-Well Thermal Cycler (Applied Biosystems, Singapore). The cycling conditions were 94 °C for 5 min, 35 cycles of 94 °C for 30 s, 55 °C for 40 s and 72 °C for 45 s, followed by a final extension of 72 °C for 10 min. We used DNA from Cryptosporidium spp. provided by Theodor Bilharz Research institute (TBRI) (Giza Governorate, Egypt) as the positive control, and type I water as the negative control.
The PCR products were electrophoresed in 1% agarose gel stained with the ethidium bromide dye, then PCR photos were captured using a gel documentation system (Alpha Innotech, Biometra), using Automatic Image Capture Software (ProteinSimple, CA, USA). The QIAquick PCR product Purification Kit (Qiagen, Gmbh, Germany) was used to purify the positive PCR products.
Sequencing and phylogenetic analysis
A purified PCR product was sequenced in the forward and reverse directions using Big-Dye^®^ Terminator v3.1. Ready Reaction Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA, Cat. No. 4336817) following the manufacturer’s instructions. Purification of the sequencing reaction was conducted using a Centrisep spin column (Cat. No. CS-901), according to the instructions of the manufacturer.
The obtained nucleotide sequences were compared to reference sequences in the GenBank database using the basic local alignment search tool (BLAST) (http://www.ncbi.nlm.nih.gov/BLAST/). The sequences were edited and aligned in the ClustalW v.1.8 multiple sequence alignment programme [11]. The phylogenetic tree was constructed with the Molecular and Evolution Genetic Analysis (MEGA v6.0) software package, using the maximum likelihood approach with the Tamura-3 parameter model. The robustness of the tree was tested with 1000 bootstrap replications [12]. The nucleotide sequences acquired in the present study were deposited in the GenBank database under accession number (OP716766) (https://www.ncbi.nlm.nih.gov/nuccore/OP716766.1/).
Results and discussion
Despite the recent assessment of water-borne cryptosporidiosis in Sharqia Province [7], this report is the first to identify Cryptosporidium species in the drainage canals extending across the district of Minya Al-Qamh in the East Delta Governorate. In the current study, C. parvum was isolated from one sample 1.4% (1/72) belonging to the village of Al-Aziziyyah. Water and environmental samples are known to be a challenging matrix for molecular analysis due to the presence of PCR inhibitors, which could affect the process of DNA extraction [13]. In addition, contamination of water samples with human or animal faeces, soil, and sand can interfere with the release of DNA from the oocysts [14]. All these factors may lead to false negative results during DNA amplification, contributing to the reported low recovery rate of Cryptosporidium infection.
In the current study, the partial sequencing of the (COWP) gene has enabled the classification of the Cryptosporidium strain isolated from the drainage canals of Al-Aziziyyah village. According to the obtained phylogenetic tree (Fig. 2), the isolated Cryptosporidium strain analyzed in the current study was identified as Cryptosporidium parvum (C.parvum), since it clustered in the same clade with other C.parvum strains. Rural Egypt marks greater exposure to livestock and other animals, which have free access to nearby streams and watercourses. In addition, the significant level of farming activities in such agricultural areas is among the factors explaining the dominance of bovine cryptosporidiosis in the Nile Delta Region [15]. These findings correspond well with previous research assessing the prevalence rates of Cryptosporidium infection among children in rural and urban settings. Children in rural communities had higher infection rates of C. parvum than children in urban areas where C. hominis predominates [16].
Fig. 2. Phylogenetic analysis of Cryptosporidium species based on partial (COWP) gene nucleotide sequence. The phylogenetic tree was constructed using the Tamura-3 parameter model with MEGA v6.0. Bootstrap values were calculated with 1000 replicates. Reference nucleotide sequences were retrieved from the GenBank. The representative Cryptosporidium isolate detected in the present study is marked by a red circle. The scale bar indicates nucleotide substitutions per site
Data about Cryptosporidium genotyping and species identification in different Egyptian water supplies remain scarce. The documented reports are only available from 5 studies in Ismailia, Gharbiya (Tanta City), Assiut and Beni-Suef governorates. Most of these studies, in line with our findings, recorded a predominance of the C. parvum genotype contaminating canal and tap water sources (76.2%) in Ismailia Governorate [17], tap and tank water supplies (55%) in Tanta City [15], water treatment plants (79%) and potable tap water in Assiut Governorate [18, 19, respectively]. Conversely, the governorate of Beni-Suef exhibited a predominance of the anthroponotic species C. hominis (75.9%) over C. parvum (20.7%) in the tested tap water samples, suggesting that human activities with person-to-person transmission are the primary sources of water contamination [20].
Results of the sequence analysis (Fig. 3) has revealed the presence of the bovine genotype, C. parvum (OP716766), which showed 100% homology with the sequences isolated from dairy calves at Kafr El Sheikh Province in Egypt (AB514064) [21], infected humans and cattle in The Netherlands (DQ388390) [22], the sea mussels species, Mytilus galloprovincialis and Mytilus edulis farmed and sold in Italy (KY706489) [23], and an adult woman in Japan, who suffered severe diarrhoea (AB089292) [24].
Fig. 3. Sequence distance matrix showing percentages of identity and divergence among the analyzed (COWP) gene sequences. A red circle marks the study isolate. The nucleotide sequences were retrieved from the GenBank database after performing BLAST analysis
It has been suggested that zoonotic transmission is the reason behind the predominance of C. parvum in a population. Yet, the Cryptosporidium host adaptation is not strictly host-specific, and cross-species transmissions may occur. Therefore, the source of C. parvum in humans can also be of human origin, and many cases of human C. parvum infections may not be zoonotic. Recent molecular subtyping has revealed the wide distribution of human-adapted C. parvum, especially in developing countries, proving the anthroponotic transmission of C. parvum subtypes shared by humans and cattle [6]. Hence, we suggest that the source of the canal water contamination reported in the present work could originate from humans and/or cattle. In the rural Sharqia Governorate, animal and human waste are frequently dumped into canals and drains, introducing Cryptosporidium oocysts to the waterways used by farmers. Also, heavy rainfall contributes to the transfer of Cryptosporidium from soil contaminated with human and animal faeces to unprotected surface water [7].
Conclusions
The involvement of water in the transmission chain of cryptosporidiosis is a topical health concern of direct relevance to public health authorities and water consumers alike. Therefore, molecular tracing is critical to identifying the host source of the parasites contributing to environmental contamination. As a result, there is a need for comprehensive studies based on robust analysis of several different gene markers to allow the identification of Cryptosporidium to the species and sub-species levels.
Study limitations
Although this work is the first to document the molecular identification of Cryptosporidium isolates contaminating the canal water bodies in Sharqia Province, the study has certain limitations related to:
- The relatively low number of samples.
- The limited geographical representativeness.
- The presence of PCR inhibitors in the tested environmental samples.
- The use of a single marker, the (COWP) gene, for the genotyping of the recovered Cryptosporidium species.
- Contamination of water with human and animal waste, which could dilute the target DNA.
- The absence of alternative diagnostic measures, such as immunochromatographic tests (ICT) or immunofluorescence assays (IFA).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cama VA, Bern C, Roberts J, Cabrera L, Sterling CR, Ortega Y, Gilman RH, Xiao L. Cryptosporidium species and subtypes and clinical manifestations in children, Peru. Emerg Infect Dis. 2008;14:1567–74. 10.3201/eid 1410.07127310.3201/eid 1410.071273 PMC 260988918826821 · doi ↗ · pubmed ↗
- 2Omar M, Etewa SE, Mahmoud SAM, Farag TI. Assessment of the potential occurrence of Cryptosporidium species in various water sources in Sharqia governorate, Egypt. J Parasit Dis 2024 48:358–69. 10.1007/s 12639-024-01675-110.1007/s 12639-024-01675-1PMC 1114797138840871 · doi ↗ · pubmed ↗
- 3Amer S, Honma H, Ikarashi M, Tada C, Fukuda Y, Suyama Y, Nakai Y. Cryptosporidium genotypes and subtypes in dairy calves in Egypt. Vet Parasitol. 2010;169:382–6. 10.1016/j.vetpar.2010.01.01710.1016/j.vetpar.2010.01.01720193982 · doi ↗ · pubmed ↗
- 4Tedde T, Marangi M, Papini R, Salza S, Normanno G, Virgilio S, Giangaspero A. Toxoplasma gondii and other zoonotic protozoans in mediterranean mussel (Mytilus galloprovincialis) and blue mussel (Mytilus edulis): A food safety. Concern? J Food Prot. 2019;82:535–42. 10.4315/0362-028X.JFP-18-15710.4315/0362-028X.JFP-18-15730810381 · doi ↗ · pubmed ↗