The pattern of gene amplification of members of the Plasmodium vivax erythrocyte binding-like proteins family across the Amazon rainforest
Karla S. Guedes, Gabriela M. Fernandes, Bruno A. M. Sanchez, Francis B. Ntumngia, John H Adams, Flora S. Kano, Cor J. F. Fontes, Tais N. Sousa, Luzia H. Carvalho

TL;DR
This study examines gene amplifications in Plasmodium vivax across the Amazon and finds regional differences but no link to host DARC genotypes.
Contribution
The study reports regional variation in dbp and ebp2 gene amplifications in P. vivax isolates from the Amazon rainforest.
Findings
dbp gene amplification was highest in the western Amazon region (25%) with up to 8 copies per isolate.
ebp2 amplification was less frequent, detected in 15% of isolates in the western region and 1–4% elsewhere.
No association was found between gene copy number variations and DARC genotypes in the study population.
Abstract
In Plasmodium vivax malaria, the Duffy Binding Protein (DBP), a key erythrocyte binding-like (EBL) protein, enables invasion of DARC (Duffy Antigen/Receptor for Chemokines) positive reticulocytes. Another EBL member, the erythrocyte binding protein 2 (EBP2, also known as EBP/DBP2), shares structural features with DBP, though its function in erythrocyte invasion remains unclear. While copy number variation (CNV) in EBL genes have been well-documented, data on dbpand ebp2 amplifications in P. vivax isolates from South America remain scarce. This study investigates CNV in these two ebl genes across Amazon populations and examines their association with host DARC genotypes. A total of 192 P. vivax isolates from three malaria-endemic sites of the Amazon region (eastern, western, and southwestern) were analyzed. DARC polymorphisms were genotyped by real-time PCR with allele-specific…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
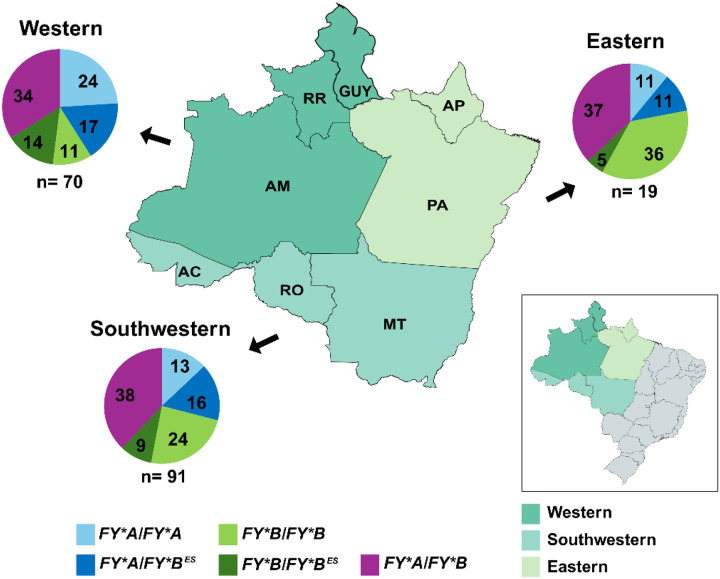 Figure 1
Figure 1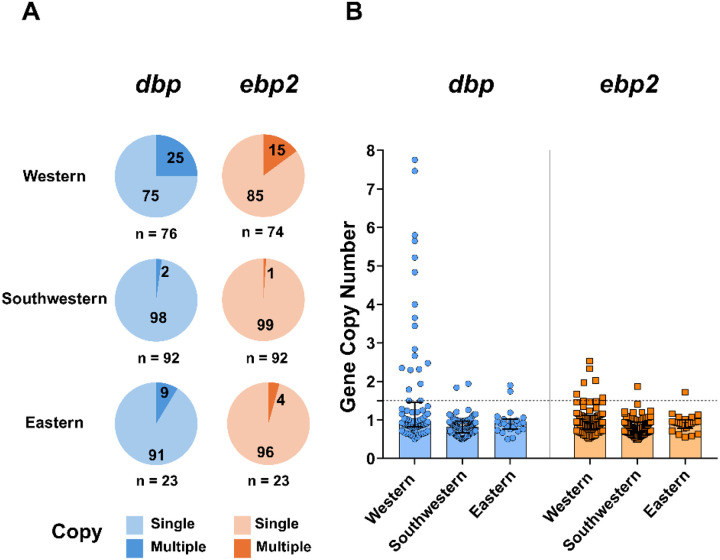 Figure 2
Figure 2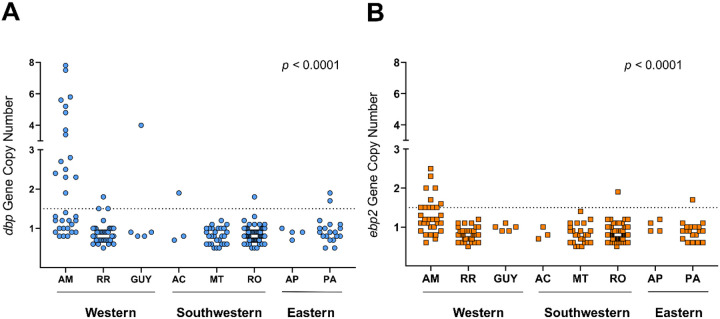 Figure 3
Figure 3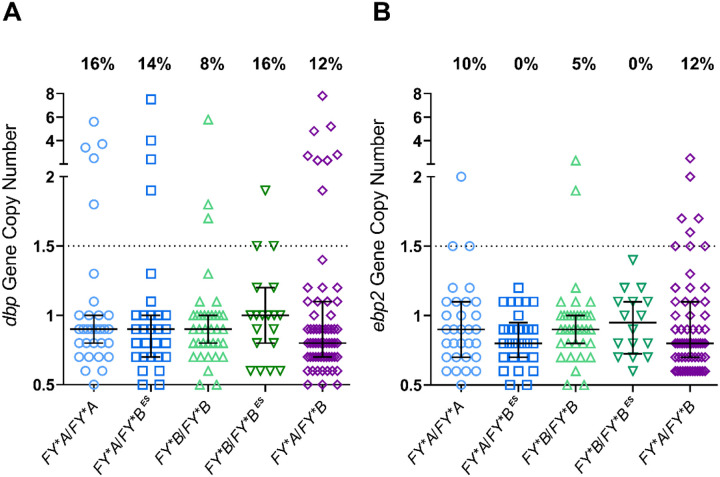 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Mosquito-borne diseases and control · Hemoglobinopathies and Related Disorders
Background
The invasion of red blood cells by Plasmodium spp. is a complex biological process that requires the coordinated engagement of multiple parasite ligands with specific receptors on the erythrocyte membrane [1, 2]. In the case of Plasmodium vivax, the most widespread human malaria parasite [3], there are distinct ligand-receptor interactions that provide strict selectivity for reticulocytes [4–6]. The process of reticulocytes invasion by P. vivax depends on the Erythrocyte Binding-Like (EBL) proteins family [7] whose best-characterized member is the Duffy binding protein (DBP), which binds to the human atypical chemokine receptor 1 (ACKR1 also named Duffy antigen receptor for chemokines, DARC) [8]. The interaction between the ligand domain of DBP (region II, DBPII) and DARC receptor is central to blood-stage infection, as individuals lacking DARC on the surface of the RBCs (DARC-negative) exhibit significantly reduced susceptibility to P. vivax infection [9]. Thus, a vaccine that elicits antibodies able to block the DBPII/DARC interface would be expected to block parasite invasion (reviewed by [10]).
The long-standing hypothesis that P. vivax invades reticulocytes exclusively through the DBPII-DARC interaction has been increasingly challenged by numerous reports of P. vivax infections in DARC-negative individuals [11]. Towards understanding the mechanisms of P. vivax invasion in DARC-negative individuals, numerous studies have investigated other parasite ligands and alternative receptors (reviewed by [12]). In 2013, a notable study by Hester et al. [13], which involved de novo assembly of a field isolate genome, revealed novel P. vivax erythrocyte invasion genes. One of these genes coding a novel P. vivax DBP homolog, erythrocyte binding protein 2 (EBP2 also termed EBP/DBP2). EBP2 primary structure has the key conserved domain features characteristic of EBL protein family, including a DBL ligand domain (region II) considered essential for receptor recognition and merozoite invasion [7]. While some studies suggest that EBP2 binds preferentially to immature DARC-positive reticulocytes [14], others suggest that it binds to both reticulocytes and normocytes, independently of the DARC receptor [15]. Although the role of EBP2 in P. vivax reticulocyte invasion is still unknown, the combined antibody response to DBP and EBP2 has an additive effect in reducing the risk of P. vivax malaria in PNG children [16, 17].
The detection of gene copy number variation (CNV) in DBP and EBP2 added another layer of genetic complexity to P. vivax infections. In 2013, Menard et al. reported the first evidence of dbp gene amplification in P. vivax isolates from Madagascar, where infections occur in both DARC-positive and DARC-negative individuals [18]. The same authors detected dbp duplication in P. vivax infected blood samples from travelers returning from America, Africa, Asia and Melanesia. Subsequent studies confirmed that P. vivax strains carrying multiple copies of the dbp gene have a global distribution [19–24] and do not seem to have any association with the DARC negativity barrier [25, 26]. At this point, there is still no consensus about the role of dbp gene amplification in the P. vivax infection [12]. Several hypotheses have been proposed, including the possibility that gene amplification enables the parasite to evade the host immune response [26, 27].
In contrast to DBP, few studies have explored the genetic diversity of ebp2 gene. Most available data is from Asian and, to a lesser extent, African populations [25, 28]. Recently, we have analyzed P. vivax isolates from the Brazilian Amazon region and found that the DBL domain of EBP2 is significantly less polymorphic than of DBP [29], a finding consistent with reports from other regions [13, 25, 28, 30]. Although studies on ebp2 gene amplification remain scarce, available data suggest that gene multiplication is common in African populations, in both DARC-positive and DARC-negative individuals [22, 23, 25].
To gain insight into the gene amplification of the two main EBL proteins currently known to be involved in reticulocyte invasion (DBP and EBP2), this study assessed the copy number variation of the dbp and ebp2 genes in P. vivax isolates from different endemic areas of the Amazon region, where malaria transmission ranges from hypoendemic to mesoendemic [31]. Considering that malaria transmission in the Amazon rainforest is associated with DARC-positive carriers, we further explore the correlation of gene copy number with DARC genotypes.
Methods
Ethical approval and consent to participate
The ethical and methodological procedures of this study were approved by the Human Research Ethics Committee of the René Rachou Institute / FIOCRUZ Minas (approval No. 4,041,859; CAAE 15632719.0.0000.5091), as well as by the Research Ethics Committee of the Júlio Muller University Hospital (approval No. 53041521.6.0000.5541). The study participants were provided with information regarding the objectives and procedures of the study, and their voluntary participation was requested and confirmed through written formal consent. In the case of child participants, written formal consent was obtained from their next of kin, caregivers, or guardians. Data were separated from personal identifiers through use of a code. All biological samples analyzed in this study were stored in the biorepository of the René Rachou Institute – FIOCRUZ Minas, under the following approved protocols: No. 07/2006, No. 07/2009, No. 12/2010, No. 26/2013, and No. 1.821.955/2016. The biorepository is curated by L. H. Carvalho and F. S. Kano.
The current study was conducted according to Laboratory biosafety and biosecurity policy guidelines of the Oswaldo Cruz Foundation (FIOCRUZ, Ministry of Health, Brazil (http://www.fiocruz.br/biosseguranca/Bis/manuais/biosseg_manuais.html).
Study sites and sample collection
A total of 192 Plasmodium vivax field isolates were collected from patients across three regions of the Amazon, grouped as follows (Fig. 1, on the inset map): eastern Amazon — Amapá (AP, n = 4) and Para (PA, n = 19); western Amazon — Amazonas (AM, n = 32), Roraima (RR, n = 39), and Guyana (GUY, formerly British Guiana, n = 5); and southwestern Amazon — Rondônia (RO, n = 60), Mato Grosso (MT, n = 30), and Acre (AC, n = 3). The samples were collected between 2017 and 2023. Blood samples (5 mL) were obtained via venipuncture using EDTA as an anticoagulant, following written informed consent from all study participants.
Malaria diagnosis
At the time of blood collection, all individuals were submitted to a finger-prick for malaria diagnosis by light microscopy. The Giemsa-stained thick blood smears were prepared and examined by experienced local microscopists, according to the malaria diagnosis guidelines of the Brazilian Ministry of Health. Genomic DNA was extracted from whole blood samples collected in EDTA, using the Puregene blood core kit B (Qiagen, Minneapolis, MN, USA), according to manufacturers’ instructions. Species-specific real-time PCR was performed as previously described [32].
DARC genotyping
Two TaqMan assays (Applied Biosystems, USA) were used to genotype major DARC polymorphisms: the T-33C substitution in the RBC-specific GATA1 transcription factor binding motif (rs2814778, ID assay reference: C_15769614_10), and the G125A polymorphism in exon 2 (rs12075, ID assay reference: C_2493442_20). The amplification reactions were carried out on a ViiA7 Real-Time PCR System (Applied Biosystems, USA), using 2.5 μL of TaqMan^®^ 2x Universal PCR Master Mix, 0.25 μL of the specific TaqMan^®^ SNP Genotyping Assay, 1.25 μL of DNase and RNase-free water, and 1 μL of genomic DNA (~ 10 ng/μL). Each assay included both negative controls (water in place of DNA) and positive controls, consisting of previously genotyped samples, as previously described [33]. Data analysis was performed using QuantStudio Real-Time PCR Software v1.3.7 (Applied Biosystems, USA).
Relative quantification of dbp and ebp2 copy number
Gene copy number was estimated by quantitative PCR (qPCR), following the optimized protocol of Roesch et al. (2018) [25]. The β-tubulin gene was used as an internal reference. qPCR reactions were performed in 10 μL total volumes, containing 5 μL of GoTaq^®^ qPCR Master Mix 2X (Promega), 2.2 μL of DNase and RNase-free water, and 1 μL of template DNA. Primer concentrations were as follows: 900 nM for dbp (CN_DBP_F and CN_DBP_R); 300 nM for ebp2 (CN_EBP2_F and CN_EBP2_R) and β-tubulin (CN_3-tubulin_F, and CN_ β-tubulin _R) (Table S1).
Amplification and fluorescence detection were carried out on a ViiA7 Real-Time PCR System (Applied Biosystems, USA), using the following cycling conditions: 50°C for 2 minutes, 95°C for 10 minutes, followed by 40 cycles of 95°C for 15 seconds and 60°C for 1 minute, with a final step of 95°C for 15 seconds. All reactions were run in triplicate on 96- or 384-well plates. The copy numbers were estimated relative to a standard curve by using synthetic genes construct (gBlock) containing a single copy each of β-tubulin, dbp, and ebp2, on a 1:1:1 ratio, designed by Integrated DNA Technologies, Inc. (IDT, USA, 2023). Two commercial plasmids, pIDTSMART-AMP, also synthesized by IDT, were used for optimization: one containing dbp and β-tubulin inserts (1:1), and the other with ebp2 and β-tubulin (1:1). Data were processed and 2^−ΔΔCT^ method (where CT is the cycle threshold) was used to determine the number of copies of each sample. A positive control consisting of an isolate previously validated to carry one copy of each gene (confirmed by independent methods) and a negative control (water in place of DNA) were included in all runs. Replicates with a standard deviation greater than 0.3 or with CT values above 32 were excluded from analysis. Samples showing copy number variation were re-tested in independent experiments for confirmation. Isolates with estimated copy numbers below 0.5 were discarded. Final copy number values were rounded as follows: 0.5–1.4 to 1 copy, ≥ 1.5–2.4 to 2 copies, ≥ 2.5–3.4 to 3 copies, ≥ 3.5–4.4 to 4 copies, ≥ 4.5–5.4 to 5 copies, ≥ 5.5–6.4 to 6 copies, ≥ 6.5–7.4 to 7 copies, and ≥ 7.5–8.4 to 8 copies. Reactions were analyzed using QuantStudio Real-Time PCR Software v1.3.7 (Applied Biosystems, USA).
Statistical analysis
A database was created using EpiData software (www.epidata.dk). The graphics and the statistical analyses were conducted using GraphPad Prism version 9.5.0 (GraphPad Software, La Jolla, CA, USA) and Stata software version 12.0 (StataCorp, College Station, TX, USA. Data normality was evaluated using the Shapiro–Wilk test. For comparisons of non-parametric continuous variables, the Kruskal-Wallis test was employed. Categorical variables were analyzed using the Chi-square test or Fisher’s exact test, as appropriate. Two-sided p-values < 0.05 were considered statistically significant. All data necessary to replicate the study’s findings are openly available (Table S2).
Results
Molecular confirmation of P. vivax infection and DARC genotypes in the study population
Of the 192 P. vivax infections detected by microscopy in this study, 182 (92%) were confirmed as P. vivax monoinfections by a species-specific PCR assay, with 10 (5%) of them identified as mixed infections with P. vivax and P. falciparum (Table S2). Most P. vivax patients included in the study were male (n = 146, 76%), with a median age of 36 years (IQR 26–45 years). In the study area, all individuals were DARC positive, with a predominance of FYA/FYB genotype in all Amazon regions (34–38%) (Fig. 1). DARC positive homozygous (FYA/FYA and FYB/FYB) and DARC-null positive genotypes (FYA/FYB^ES^ and FYB/FYB^ES^) were also detected in high frequencies in the study area (Fig. 1). Except for the genotype FYB/FYB, there were no significant differences between the Amazon regions (Table S3).
dbp and ebp2 gene copy number variation in the Amazon
The copy number variation (CNV) in the dbp and ebp2 genes among P. vivax isolates showed differences across the distinct regions of the Amazon. The western region showed higher CNV frequencies, with 25% (19 of 76) and 15% (11 of 76) of isolates harboring CNVs in dbp and ebp2, respectively (Fig. 2A). In contrast, in the southwestern region, gene duplication frequencies were detected at much lower numbers, ranging from 2–1% for dbp (2 of 91) and ebp2 (single isolate), respectively. Low CNV frequencies were also detected in the eastern region for both genes (9 to 4%; Fig. 2).
In terms of region, only P. vivax isolates from the western region exhibited up to eight copies of the dbp gene (Fig. 2B), while those from the eastern and southwestern regions exhibited a maximum of three copies. For the ebp2 gene, the western region presented an estimated copy number of approximately three, whereas isolates from the other regions showed up to two copies. It is noteworthy that 10 out of 26 P. vivax isolates exhibited concurrent amplification of the dbp and ebp2 genes, while most of them (16 out of 26) occurred in only one of the genes, i.e., 13 for dbp and 3 for ebp2 (Table S4).
At the local level, CNVs in dbp gene were detected in samples from Amazonas (AM), Roraima (RR), Para (PA), Guyana (GUY), Acre (AC), and Rondonia (RO), with AM presenting most of the gene multiplication (15 out of 23, 65%) (Fig. 3A). In contrast, ebp2 CNV was restricted to isolates from AM, PA, and RO, with Amazonas accounting for 11 of 13 (85%) gene multiplications (Fig. 3B).
Distribution of dbp and ebp2 gene copy numbers according to DARC genotype
Finally, it was investigated whether the occurrence and frequency of amplification of the dbp and ebp2 genes could be associated with DARC genotypes. For this purpose, the gene amplification analysis stratified the total population according to host DARC genotype (Fig. 4). Taken together, the results demonstrated that neither dbp nor ebp2 copy numbers were associated with any DARC genotype (Table S5, Fig. 4).
Discussion
In different Amazon endemic settings, this study investigated copy number variation (CNV) in two P. vivax genes encoding proteins implicated in blood-stage invasion: DBP, a well characterized ligand that mediates invasion of DARC positive reticulocytes, and EBP2, a paralog with putative functional redundancy. The results of the gene amplification showed significant differences among the Amazon endemic sites, with the western region accounting for most parasites with multiple copies of the dbp or ebp2 genes. Specifically, 25% of P. vivax isolates from the western region showed dbp gene amplification of up to 8 copies, compared to only 2–9% in the southwestern and eastern regions with 2–3 copies.. A similar pattern was observed for ebp2, with 15% in the western versus 1–4% in other regions. Unfortunately, the limited data available on dbp CNV- and the absence of data on ebp2- in the Brazilian Amazon region preclude any comparison of our findings with previous studies in the region. Despite of that, multiple copies of DBP were detected in 10% of P. vivax isolates from urban areas of Acre state, southwestern Amazon region [20]. Taken together, these results from the Amazon support the current view that copy number variation in genes associated with P. vivax reticulocyte invasion seems to be a common phenomenon occurring on a worldwide scale [18, 20, 21, 23–25, 34].
In the current study, the reason for the different spatial distribution of dbp and ebp2 copy numbers among the three P. vivax-endemic Amazon settings is not clear. It is unlikely that variation in the host DARC gene accounts for the observed regional differences in gene duplication. Although the distribution of FYB/FYB genotype showed slight variation between the three regions, the overall DARC genotype frequencies were similar across the three Amazonian regions, with a predominance of the FYA/FYB genotype; this pattern reflects a population structure influenced by both Amerindian and European ancestry components [35]. Notably, most P. vivax isolates from the western Amazon originated from a long-term P. vivax-exposed population (Rio Pardo, AM, Table S2) formed mainly by Amazonian natives previously characterized by high contribution of Native Amerindian ancestry [35–37]. In contrast, Plasmodium vivax isolates from the southwestern and eastern Amazon are mainly derived from migrants originating in non-endemic regions of Brazil. In these regions, malaria is predominantly an occupational disease, strongly associated with activities such as deforestation and illegal mining [38]. Consequently, multiple, not-mutually exclusive, factors may have contributed to the difference in the CNV profiles between the western and southwestern/eastern regions, including differences in the intensity of malaria transmission, historical parasite population dynamics and host genetics [21]. Future studies with more presentative samples of the Amazon basin should address this topic.
Given that our previous findings and those of others have shown that susceptibility to P. vivax clinical malaria is influenced by DARC genotypes [35, 39, 40], we further investigated whether gene copy number variation was associated with DARC genotype. In this DARC positive population, no association was found between dbp or ebp2 gene amplifications and DARC homozygous or heterozygous genotypes. Similar findings were reported in P. vivax isolates from Cambodia and Ethiopia, where DBP duplication was not linked to any specific DARC-positive genotype [20, 26]. Despite this, in populations where DARC-positive and DARC-negative individuals coexist, it has been suggested that the proportion of single-versus multicopy variants of DBP or EBP2 may differ between these groups [21, 22, 34]. It seems to be a consensus that larger studies are needed to further elucidate the role (if any) of DARC in DBP/EBP2 copy number variations.
While DBP duplication has been extensively studied in parasites from diverse geographical regions (reviewed by [12]), EBP2 amplification has received far less attention [22, 23, 25]. In our study, ebp2 duplication was detected across all three surveyed regions in the Amazon (4–15%), albeit at a lower frequency than dbp amplification (2–25%). Duplication events in P. vivax isolates were predominantly gene-specific, with 62% (16 of 26) of samples displaying amplification in only one of the two erythrocyte-binding genes. Compared to isolates from the Amazon region, P. vivax isolates from Asia and Africa typically exhibit much higher frequencies of EBP2 gene duplications (19 to 56%) [22, 25]. In Ethiopia, where DBP and EBP2 duplications are common, EBP2 amplification (but not DBP) is more prevalent in DARC-positive (46%) than in DARC-negative individuals (21%) [22]. This observation is further supported by recent evidence from Central Africa, where most of P. vivax isolates from DARC-negative individuals exhibited more than two dbp copies in the absence of ebp2 amplification [23]. This is of particular interest as we and others have demonstrated that EBP2 can bind strongly to DARC-positive reticulocytes and moderately to DARC-negative reticulocytes [14, 19].
The current study has limitations that should be consider on data interpretation. First, all individuals presented an acute symptomatic P. vivax infection, with no quantitative assessment of parasitemia in thick-blood smear. Consequently, we were unable to evaluate whether levels of parasitemia would influence the gene copy variation, as it has been the subject of considerable speculation [20, 22, 23]; to proper address this topic and avoid discrepant results, it would be more appropriate to evaluate P. vivax biomass, as it is underrepresented in parasite counts obtained from peripheral blood smears [41]. Secondly, the relatively small sample size per site, combined with a cross-sectional design and substantial variability in the timing of data collection, precluded any attempt to conduct a temporal analysis of dbp/ebp2 CNVs over time. Longitudinal studies should address this topic. Finally, our qPCR approach estimates dbp and ebp2 gene copy numbers based on the dominant clone in each isolate. Notwithstanding, qPCR assay has been a more feasible protocol that has been used in different studies [25, 34]. Although outside the scope of the current manuscript, we agree with others that next-generation sequencing or a digital PCR approach should be able to overcome this issue of multiclonal infections [23, 25].
Conclusion
This study confirms the low but appreciable presence of dbp and ebp2 gene amplification in P. vivax isolates across the Amazon rainforest where DARC- negativity is rare. The results of these gene amplifications showed significant differences among the Amazon endemic settings, but with no association with any DARC positive genotype. Although the functional significance of DBP and EBP2 amplifications requires further investigation, this pioneering study examining amplification at these loci in the Amazon provides important insights that may support the development of vaccines targeting P. vivax parasites harboring multiple copies of these key invasion ligands.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Weiss GE, Gilson PR, Taechalertpaisarn T, Tham WH, de Jong NW, Harvey KL, Fowkes FJ, Barlow PN, Rayner JC, Wright GJ, Revealing the sequence and resulting cellular morphology of receptor-ligand interactions during Plasmodium falciparum invasion of erythrocytes. P Lo S Pathog. 2015;11:e 1004670.25723550 10.1371/journal.ppat.1004670 PMC 4344246 · doi ↗ · pubmed ↗
- 2Cowman AF, Tonkin CJ, Tham WH, Duraisingh MT. The Molecular Basis of Erythrocyte Invasion by Malaria Parasites. Cell Host Microbe. 2017;22:232–45.28799908 10.1016/j.chom.2017.07.003PMC 12801281 · doi ↗ · pubmed ↗
- 3Battle KE, Lucas TCD, Nguyen M, Howes RE, Nandi AK, Twohig KA, Pfeffer DA, Cameron E, Rao PC, Casey D, Mapping the global endemicity and clinical burden of Plasmodium vivax, 2000–17: a spatial and temporal modelling study. Lancet. 2019;394:332–43.31229233 10.1016/S 0140-6736(19)31096-7PMC 6675736 · doi ↗ · pubmed ↗
- 4Galinski MR, Medina CC, Ingravallo P, Barnwell JW. A reticulocyte-binding protein complex of Plasmodium vivax merozoites. Cell. 1992;69:1213–26.1617731 10.1016/0092-8674(92)90642-p · doi ↗ · pubmed ↗
- 5Malleret B, Li A, Zhang R, Tan KS, Suwanarusk R, Claser C, Cho JS, Koh EG, Chu CS, Pukrittayakamee S, Plasmodium vivax: restricted tropism and rapid remodeling of CD 71-positive reticulocytes. Blood. 2015;125:1314–24.25414440 10.1182/blood-2014-08-596015 PMC 4401350 · doi ↗ · pubmed ↗
- 6Chan LJ, Dietrich MH, Nguitragool W, Tham WH. Plasmodium vivax Reticulocyte Binding Proteins for invasion into reticulocytes. Cell Microbiol. 2020;22:e 13110.31469946 10.1111/cmi.13110 PMC 7003471 · doi ↗ · pubmed ↗
- 7Adams JH, Sim BK, Dolan SA, Fang X, Kaslow DC, Miller LH. A family of erythrocyte binding proteins of malaria parasites. Proc Natl Acad Sci U S A. 1992;89:7085–9.1496004 10.1073/pnas.89.15.7085 PMC 49650 · doi ↗ · pubmed ↗
- 8Wertheimer SP, Barnwell JW. Plasmodium vivax interaction with the human Duffy blood group glycoprotein: identification of a parasite receptor-like protein. Exp Parasitol. 1989;69:340–50.2680568 10.1016/0014-4894(89)90083-0 · doi ↗ · pubmed ↗