Role of dopamine in responsiveness to acute ethanol exposure in Caenorhabditis elegans
Deepa Gayadin, McKenna Prunty, Stephanie JB Fretham

TL;DR
This study explores how dopamine affects how the worm C. elegans responds to alcohol exposure.
Contribution
The study identifies dopamine's role in acute ethanol sensitivity in C. elegans.
Findings
Ethanol exposure causes dopamine release-dependent swimming-induced paralysis in C. elegans.
Dopamine signaling influences acute sensitivity to ethanol in the worms.
Findings suggest dopamine mediates acute ethanol responses in C. elegans.
Abstract
Alcohol use and abuse is a common and prevalent disorder characterized by complex and individually variable physiological effects. C. elegans demonstrate multiple ethanol-induced behaviors and are an effective model for experimentally isolating environmental and genetic factors underlying the actions of ethanol. Using wild type and dopamine signaling mutant C. elegans , the current study found that ethanol exposure results in dopamine release-dependent swimming induced paralysis and that dopamine influences acute sensitivity to ethanol. Taken together, the findings support a role for dopamine in mediating acute responses to ethanol.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
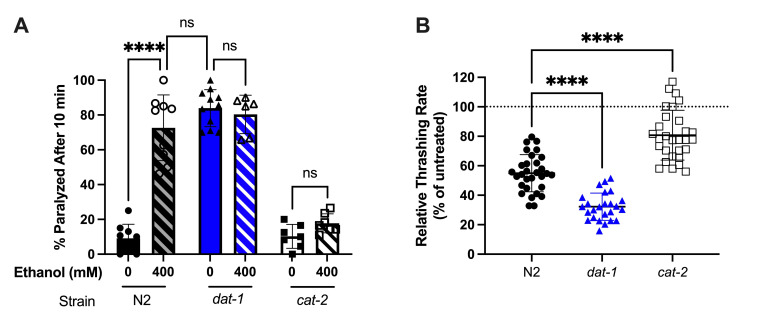 Figure 1
Figure 1|
|
|
|
|
|
|
CGC |
|
|
|
CGC |
|
|
|
CGC |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAlcoholism and Thiamine Deficiency
Description
Alcohol (ethanol, EtOH) use and abuse is a common and prevalent disorder that affected approximately 28.9 million Americans in 2022 (National Survey on Drug Use and Health, 2023). Ethanol has complex physiological effects across multiple neurotransmitter systems that can vary greatly due to individual developmental, environmental, and genetic factors (Abrahao et al., 2017; Egervari et al., 2021). Dopamine is of particular interest due to its role in reward and addiction (Nutt et al., 2015). The nematode model organism * Caenorhabditis * *elegans * ( * C. elegans * ) exhibits multiple reversible and dose-dependent behavioral responses to ethanol exposure, in particular ethanol affects locomotion rates on solid and in liquid media (Davies, 2003; Johnson et al., 2013; Scholz, 2019). The * C. elegans * nervous system is extensively characterized including dopaminergic circuitry with high homology to mammalian systems of synthesis, vesicular packaging and release, receptors, and metabolism (Engleman et al., 2016; McMillen and Chew, 2024). Furthermore, * C. elegans * exhibit several easily observable dopamine-dependent phenotypes and provide an excellent system for examining the relationship between ethanol exposure and dopamine, though relatively few studies have addressed this relationship directly. Swimming and crawling locomotion patterns, particularly the transition from swimming to crawling, are altered by acute ethanol, an effect that may be mediated through dopamine signaling (Topper et al., 2014). In addition, dopamine may facilitate context-dependent learning involving extended exposure to ethanol cues (Bettinger and McIntire, 2004; Lee et al., 2009) and contribute to the development of functional tolerance during extended ethanol exposure in * C. elegans * (Pandey et al., 2021). However, the role of dopamine in acute responses to ethanol is unclear.
First, to determine if acute ethanol exposure alters extracellular dopamine, swimming induced paralysis (SWIP) was assessed in L4 animals exposed to 0 or 400mM EtOH in ddH 2 O for 10 minutes ( ** Fig. 1A ** ). SWIP occurs when dopamine accumulates in the synapse due to excessive release or to ineffective clearance by the dopamine transporter dat-1 . Increased extracellular dopamine causes paralysis through prolonged activation of the dopamine receptor dop-3 on cholinergic motor neurons (McDonald, 2007). Notably SWIP is not observed in salty solutions such as NGM (Kudumala et al., 2019). In ddH 2 O, 400mM EtOH significantly induced SWIP in wild type N2 animals compared to 0mM EtOH. The level of ethanol-induced SWIP in N2 animals was similar to observations in * dat-1 ( ok157 ) * mutants. * cat-2 * ( * e1112 ) * dopamine-deficient tyrosine hydroxylase mutants had similar SWIP compared to N2 at 0mM EtOH, however 400mM EtOH did not induce SWIP. Taken together, these observations suggest that SWIP observed in N2 animals results primarily from ethanol-induced alterations in extracellular dopamine rather than a general paralytic effect of ethanol.
Second, to determine if dopamine contributes to the depressive locomotion effects of acute ethanol exposure, thrashing behavior was observed in N2 , * dat-1 ( ok157 ) * , and * cat-2 * ( * e1112 ) * animals ( ** Fig. 1B ** ). In 0mM EtOH liquid NGM, which does not induce SWIP behavior, no paralysis was observed at 10 minutes and all strains maintained similar thrashing rates per 30s ( N2 =53.1±8.6; * dat-1 ( ok157 ) * =51.9±6.0; * cat-2 * ( * e1112 ) * =51.5±6.8). Addition of 400mM EtOH reduced thrashing rates by 45% relative to untreated controls in N2 animals after 10 minutes, consistent with previous studies (Mitchell et al., 2007; Johnson et al., 2013). Ethanol reduced thrashing rates by 68% relative to untreated controls in * dat-1 ( ok157 ) * mutants with increased extracellular dopamine, indicating increased sensitivity to acute ethanol. Conversely, thrashing rates were only reduced by 19% in dopamine deficient * cat-2 * ( * e1112 ) * animals, indicating reduced sensitivity to acute ethanol compared to N2 animals. Taken together, these observations suggest that dopamine has a central role in determining sensitivity to acute ethanol.
Collectively, these findings indicate that dopamine mediates acute behavioral change in response to ethanol exposure, potentially through ethanol-induced dopamine release or inhibition of dopamine clearance. The SWIP phenotype has been used to study other substances including amphetamine and cannabidiol (CBD) (Carvelli et al., 2010; Kudamala et al., 2019; Shrader et al., 2020), however to our knowledge, this study is the first to demonstrate SWIP in response to ethanol and provides an additional avenue for examining genetic and environmental variants that influence ethanol sensitivity.
Methods
** * C. elegans Maintenance * **
All strains were obtained from the Caenorhabditis Genetics Center (University of Minnesota) and maintained at 20ºC on solid nematode growth medium (NGM) seeded with E. coli ( OP50-1 ). Age-synchronous animals from each strain were prepared by small-scale sodium hypochlorite treatment of gravid animals (Stiernagle, 2006) with resulting eggs maintained at 20ºC on seeded NGM plates for 3 days until reaching L4/young adult stage.
** Swimming Induced Paralysis (SWIP) Assay **
SWIP was measured as previously described (McDonald et al., 2007). Briefly, for each trial, 10 L4 animals were transferred from seeded NGM agar plates into 100μL of ddH 2 O on a depression slide. Transfer time was less than one minute. Immediately following transfer, 5μL of 8.4M EtOH or ddH 2 O was added to the slide to achieve a final EtOH concentration of 400mM. After 10 minutes, worms were scored for paralysis using a stereomicroscope. 7-11 independent replicates consisting of 3-5 trials each were measured for each strain and ethanol condition over multiple days by two different experimenters. SWIP was compared between ethanol conditions and across strains using 2-way ANOVA and Tukey's multiple comparisons in Prism (GraphPad).
** Thrashing Assay **
Thrashing was assessed according to Johnson et al., 2013. Five to seven L4/young adult worms were transferred into a depression slide containing 100µL liquid NGM and allowed to acclimate for 2-3 minutes. A baseline video recording was captured from a stereomicroscope before addition of 5µL of ddH 2 O (control) or 8.4M EtOH to achieve 400mM EtOH. A second recording was taken 10 minutes following ethanol exposure. Thrashing, defined as head movement from one side to the other, was counted for 30 seconds to determine thrashing rates. 30 worms were assessed in each strain and ethanol condition across 6 individual replicates. Thrashing rates were compared using 1-way ANOVA and Tukey's multiple comparisons in Prism (GraphPad).
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abrahao KP Salinas AG Lovinger DM 20171220 Alcohol and the Brain: Neuronal Molecular Targets, Synapses, and Circuits.Neuron 9660896-62731223123810.1016/j.neuron.2017.10.03229268093 PMC 6566861 · doi ↗ · pubmed ↗
- 2Bettinger JC Mc Intire SL 2004101 State-dependency in C. elegans.Genes Brain Behav 351601-184826627210.1111/j.1601-183X.2004.00080.x 15344920 · doi ↗ · pubmed ↗
- 3Carvelli L Matthies DS Galli A 2010421 Molecular mechanisms of amphetamine actions in Caenorhabditis elegans.Mol Pharmacol 7810026-895X 15115610.1124/mol.109.06270320410438 PMC 2912056 · doi ↗ · pubmed ↗
- 4Davies AG Pierce-Shimomura JT Kim H Van Hoven MK Thiele TR Bonci A Bargmann CI Mc Intire SL 20031212 A central role of the BK potassium channel in behavioral responses to ethanol in C. elegans.Cell 11560092-867465566610.1016/s 0092-8674(03)00979-614675531 · doi ↗ · pubmed ↗
- 5Egervari G Siciliano CA Whiteley EL Ron D 20211023 Alcohol and the brain: from genes to circuits.Trends Neurosci 44120166-22361004101510.1016/j.tins.2021.09.00634702580 PMC 8616825 · doi ↗ · pubmed ↗
- 6Engleman EA Katner SN Neal-Beliveau BS 20151124 Caenorhabditis elegans as a Model to Study the Molecular and Genetic Mechanisms of Drug Addiction.Prog Mol Biol Transl Sci 1371877-117322925210.1016/bs.pmbts.2015.10.01926810004 PMC 4834862 · doi ↗ · pubmed ↗
- 7Johnson JR Kashyap S Rankin K Barclay JW 20131114 Rab-3 and unc-18 interactions in alcohol sensitivity are distinct from synaptic transmission.P Lo S One 811e 81117 e 8111710.1371/journal.pone.008111724244732 PMC 3828271 · doi ↗ · pubmed ↗
- 8Kudumala S Sossi S Carvelli L 201943 Swimming Induced Paralysis to Assess Dopamine Signaling in Caenorhabditis elegans.J Vis Exp 14610.3791/5924331009010 PMC 7263014 · doi ↗ · pubmed ↗