Emergence and evolution of canonical microRNAs: A case study in Arabidopsis halleri and A. lyrata
Pei Qin Ng

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
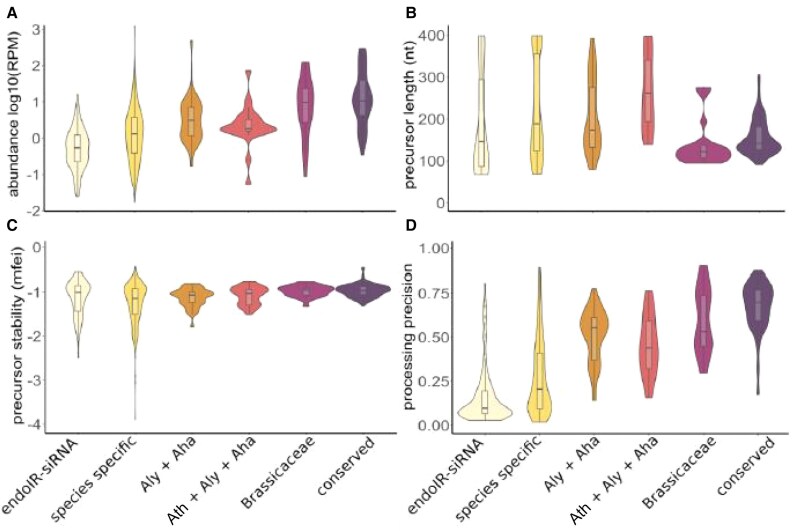 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Reproductive Biology · Photosynthetic Processes and Mechanisms
MicroRNAs (miRNAs) are a class of small RNAs that are 21 or 22 nucleotides (nts) long and have been shown to play critical roles in various plant life processes, including growth and immunity (Meyers and Axtell 2019; Lopez-Gomollon and Baulcombe 2022). Canonical miRNAs are generated through a 2-step cleavage process conducted by DICER-LIKE 1 (DCL1) from longer, imperfectly paired, hairpin-like RNA precursors (Axtell et al. 2011; Lopez-Gomollon and Baulcombe 2022; Mencia et al. 2023). The resulting mature miRNA duplex is preferentially loaded onto ARGONAUTE 1 (AGO1) proteins, forming the functional RNA-induced Silencing Complex (RISC). This complex then recognizes and silences target mRNAs through sequence complementarity. Interestingly, the miRNA population is dynamic and continuously evolving.
Unlike canonical miRNAs, which exhibit well-defined characteristics and biogenesis pathways, recently evolved or “young” miRNAs tend to have lower expression levels, less defined processing preferences, and often lack validated mRNA targets (Axtell et al. 2011; Cuperus et al. 2011). Early studies on miRNA evolution focused mainly on protein-coding genes and comparisons between distantly related plant species, limiting our understanding of the evolution of young miRNAs into canonical miRNAs.
In a recent study, Pavan and colleagues (Pavan et al. 2025) conducted small RNA sequencing on Arabidopsis lyrata and its closely related species A. halleri, which diverged approximately one million years ago (Ramos-Onsins et al. 2004). Given the fragmented state of the existing A. halleri genome, the authors generated a new high-quality, chromosome-level assembly for this study. Both the newly assembled A. halleri and the latest A. lyrata genome were annotated for miRNAs using a combination of ShortStack and MiRkwood pipelines. To validate the functionality of predicted miRNAs AGO1 and AGO4, immunoprecipitation assays were used to confirm loading, a signature feature of biologically active miRNAs. After filtering, 558 and 374 candidate miRNAs were identified in A. halleri and A. lyrata, respectively, with most miRNAs associated with AGO1.
Pavan et al. also addressed the saturation point for miRNA detection and the required sequencing depth. The authors estimated that 165 million reads are needed to detect 90% of the predicted young miRNAs, while at least 20 million reads are sufficient to detect highly expressed miRNAs. Via random subsampling, they found that miRNA detection in both species had not yet reached saturation due to limited accession numbers. These insights are valuable for guiding experimental design in studies of miRNA and other small RNAs.
The authors identified miRNAs conserved across distant plant species, within the Brassicaceae family, and among A. halleri, A. lyrata, and their common ancestor, A. thaliana. They found shared miRNAs between A. helleri and A. lyrata, as well as species-specific ones. For example, 58% of A. halleri miRNAs (n = 207) and 40% of A. lyrata miRNAs (n = 91) were species specific. These represent evolutionarily young miRNAs, given the short divergence time between species. Further characterization using linear regression modeling showed that young miRNAs tend to have lower expression, longer hairpin precursors, greater structural stability, and lower processing precision compared to canonical miRNAs (Figure).
Despite these insights, a major challenge remains in experimentally validating the biological function of these young, species-specific miRNAs. Nonetheless, Pavan et al. (2025) provide valuable insights into the transition of young miRNAs toward canonical status and highlight the dynamic nature of miRNA evolution in plants. Their study offers a framework that can be applied to other plant species to deepen our understanding of small RNA biology and evolutionary adaptation.
Recent related articles in The Plant Cell
Wang et al. (2024) reported on noncanonical long-loop precursors of miRNA858 in seed plants. This work challenges the current bioinformatics workflow that excludes long-loop precursors as viable candidates for miRNA biogenesis. Li et al. (2024) reviewed the role of biomolecular condensates, such as D-bodies for miRNA processing, in mediating RNA silencing in plants. Cao et al. (2025) showed that enhanced nuclear localization of HYPONASTIC LEAVES 1 (HYL1) is responsible for the upregulation of miRNA upon heat stress exposure rather than MIR gene upregulation. Park et al. (2023) demonstrated that nascent miRNA precursors are recognized by HYL1, allowing the recruitment of chromatin remodelers to MIRNA loci.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Axtell MJ, Westholm JO, Lai EC. Vive la différence: biogenesis and evolution of micro RN As in plants and animals. Genome Biol. 2011:12(4):221. 10.1186/gb-2011-12-4-22121554756 PMC 3218855 · doi ↗ · pubmed ↗
- 2Cao Y, Zhang J, Zhao Z, Tang G, Yan J. Heat stress triggers enhanced nuclear localization of HYPONASTIC LEAVES 1 to regulate micro RNA biogenesis and thermotolerance in plants. Plant Cell. 2025:37(6):koaf 092. 10.1093/plcell/koaf 09240266261 PMC 12142590 · doi ↗ · pubmed ↗
- 3Cuperus JT, Fahlgren N, Carrington JC. Evolution and functional diversification of MIRNA genes. Plant Cell. 2011:23(2):431–442. 10.1105/tpc.110.08278421317375 PMC 3077775 · doi ↗ · pubmed ↗
- 4Li Q, Liu Y, Zhang X. Biomolecular condensates in plant RNA silencing: insights into formation, function, and stress responses. Plant Cell. 2024:36(2):227–245. 10.1093/plcell/koad 25437772963 PMC 10827315 · doi ↗ · pubmed ↗
- 5Lopez-Gomollon S, Baulcombe DC. Roles of RNA silencing in viral and non-viral plant immunity and in the crosstalk between disease resistance systems. Nat Rev Mol Cell Biol. 2022:23(10):645–662. 10.1038/s 41580-022-00496-535710830 · doi ↗ · pubmed ↗
- 6Mencia R, Gonzalo L, Tossolini I, Manavella PA. Keeping up with the mi RN As: current paradigms of the biogenesis pathway. J Exp Bot. 2023:74(7):2213–2227. 10.1093/jxb/erac 32235959860 · doi ↗ · pubmed ↗
- 7Meyers BC, Axtell MJ. Micro RN As in plants: key findings from the early years. Plant Cell. 2019:31(6):1206–1207. 10.1105/tpc.19.0031031036598 PMC 6588298 · doi ↗ · pubmed ↗
- 8Park J, Giudicatti AJ, Bader ZE, Han MK, Møller C, Arce AL, Xu Z-Y, Yang SW, Manavella PA, Yun D-J. The HIGH EXPRESSION OF OSMOTICALLY RESPONSIVE GENE 15-HISTONE DEACETYLASE 9 complex associates with HYPONASTIC LEAVES 1 to modulate micro RNA expression in response to abscisic acid signaling. Plant Cell. 2023:35(8):2910–2928. 10.1093/plcell/koad 13237195876 PMC 10396366 · doi ↗ · pubmed ↗