Two new species and a new record of Comesomatidae (Nematoda, Araeolaimida) from the Southern Ocean

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
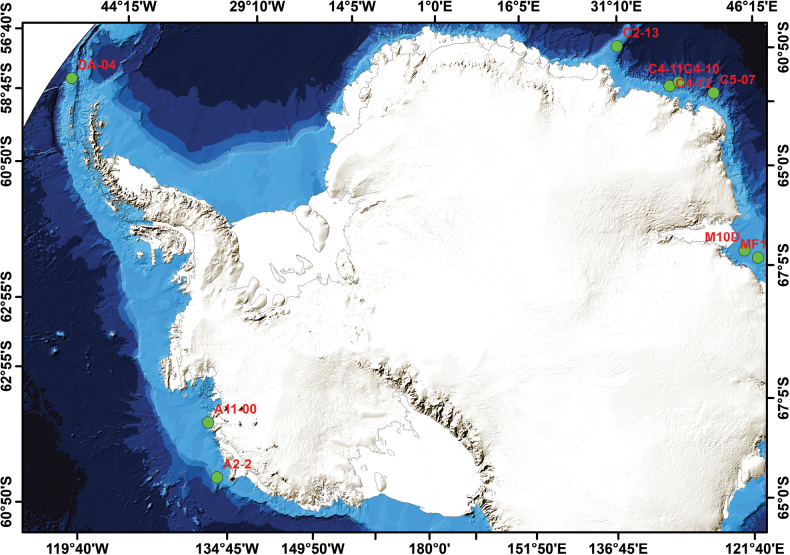 Figure 1
Figure 1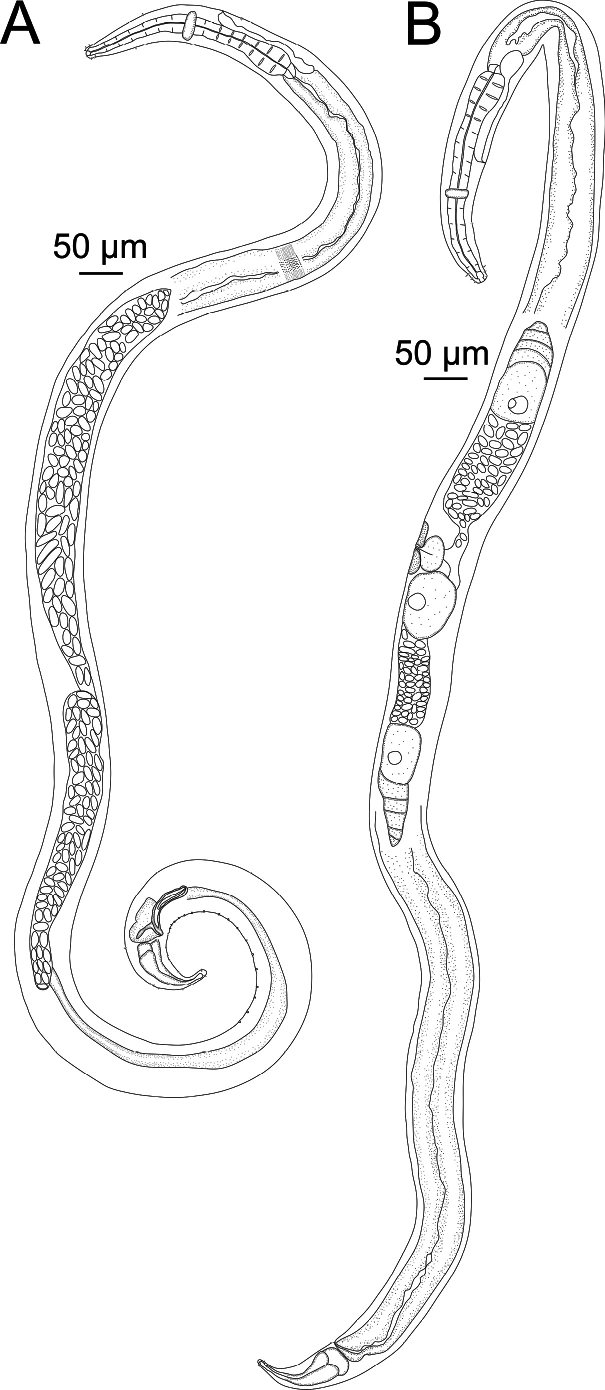 Figure 2
Figure 2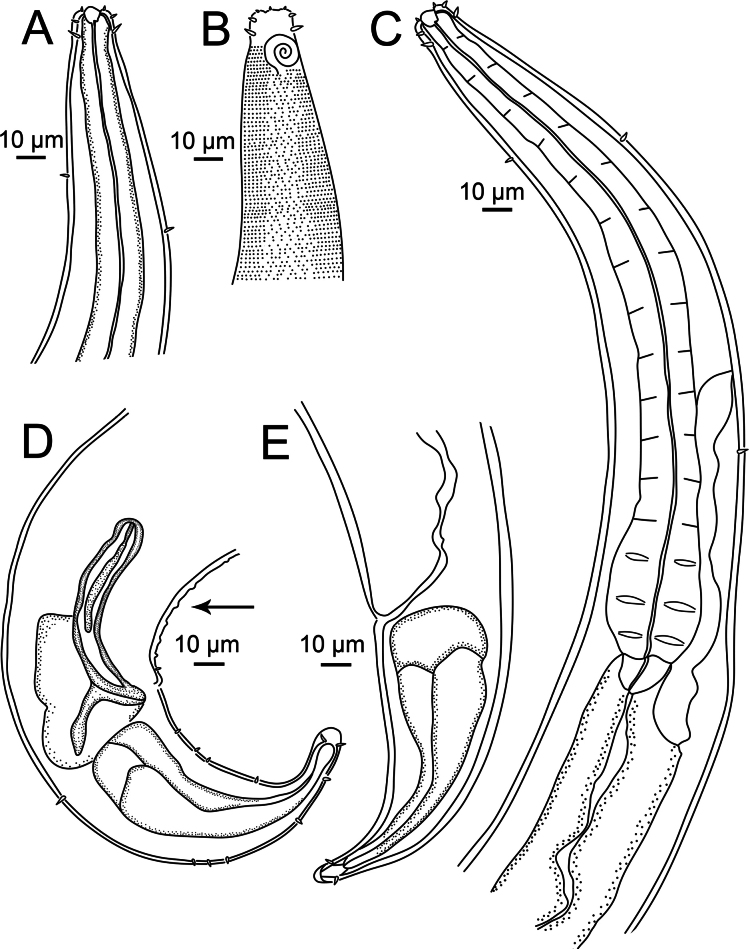 Figure 3
Figure 3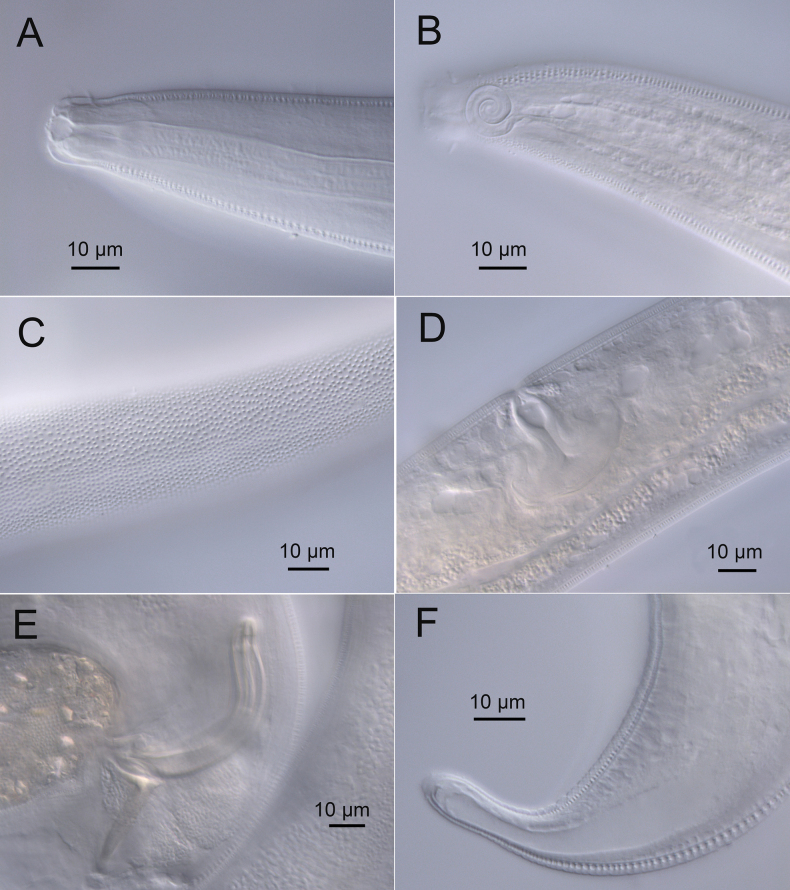 Figure 4
Figure 4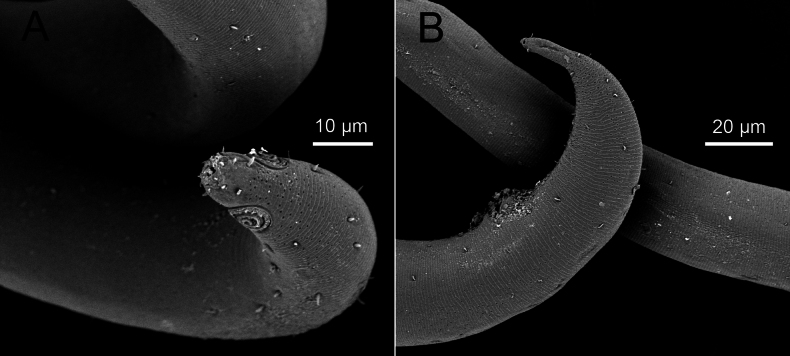 Figure 5
Figure 5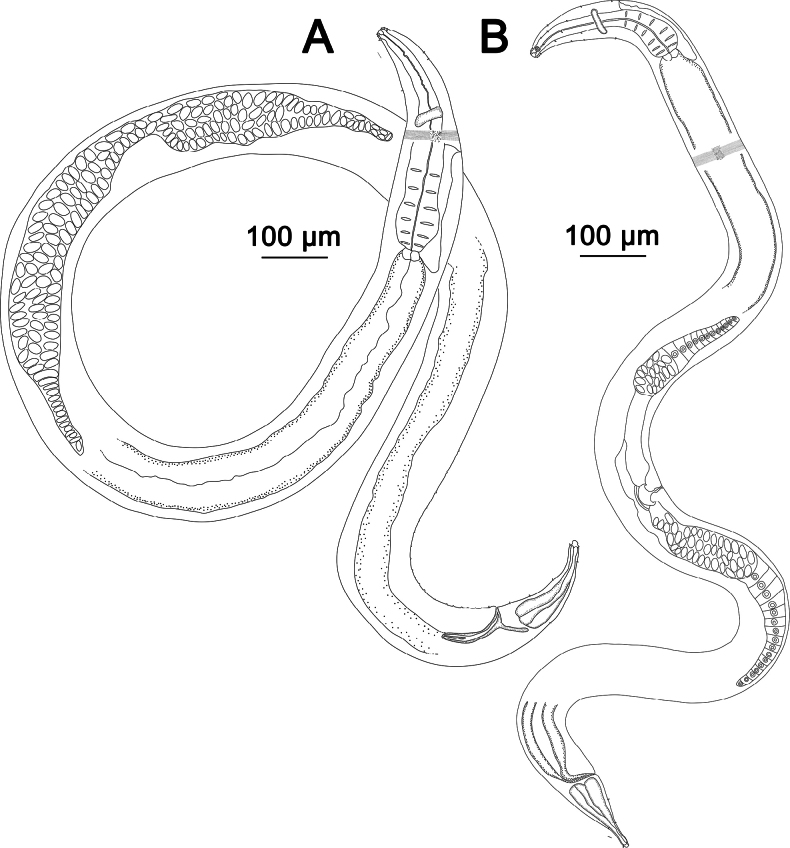 Figure 6
Figure 6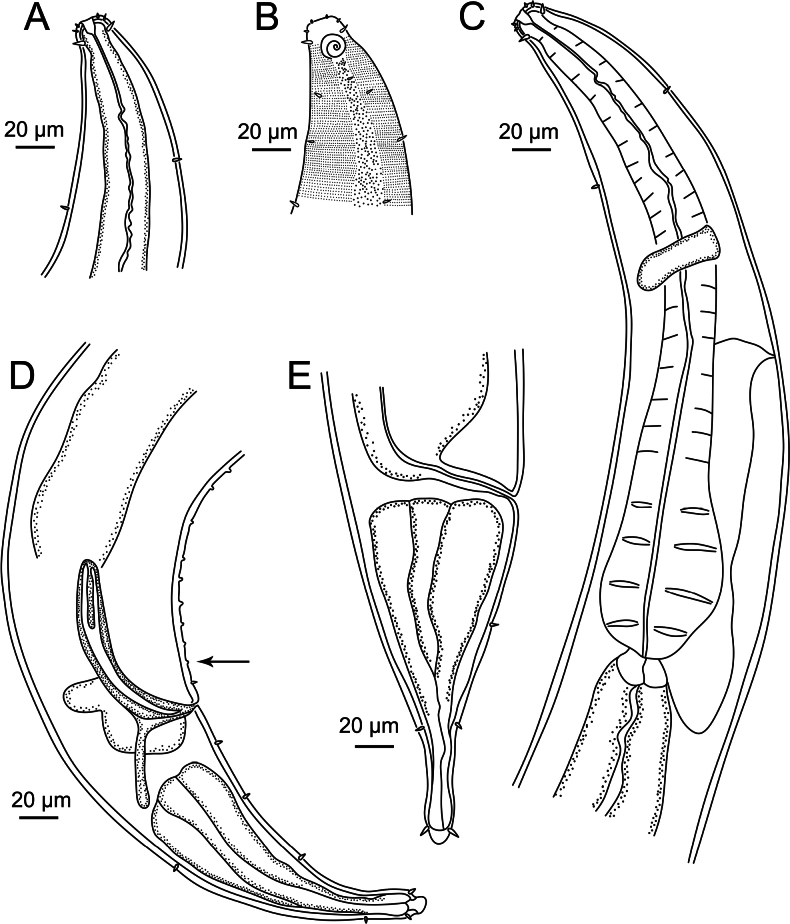 Figure 7
Figure 7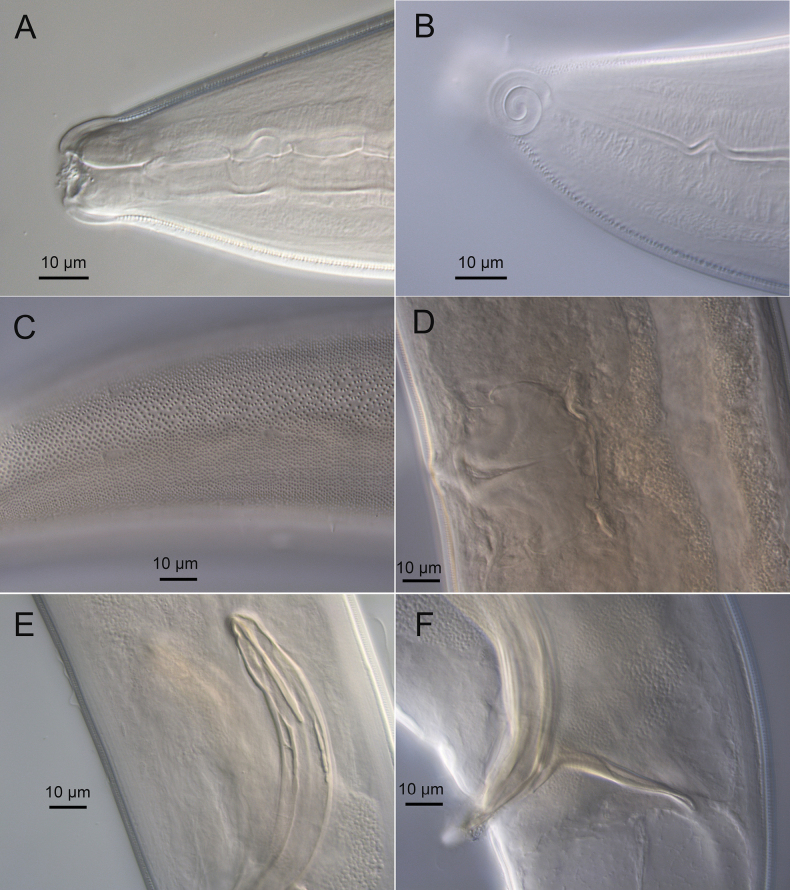 Figure 8
Figure 8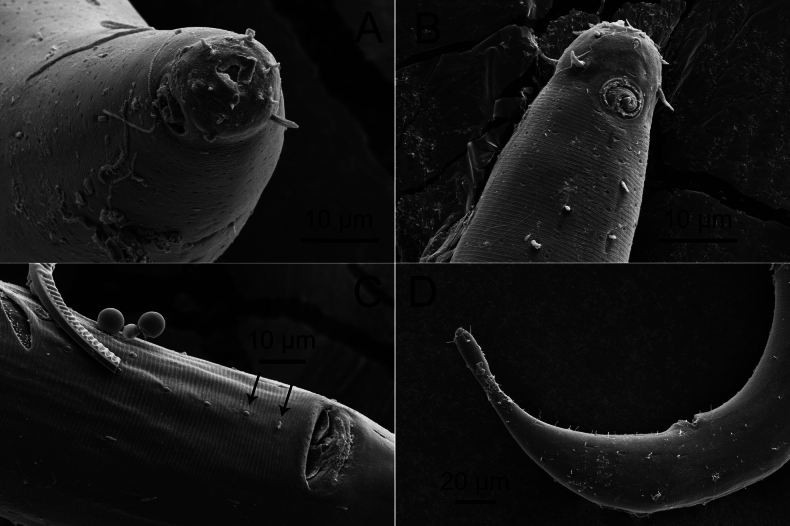 Figure 9
Figure 9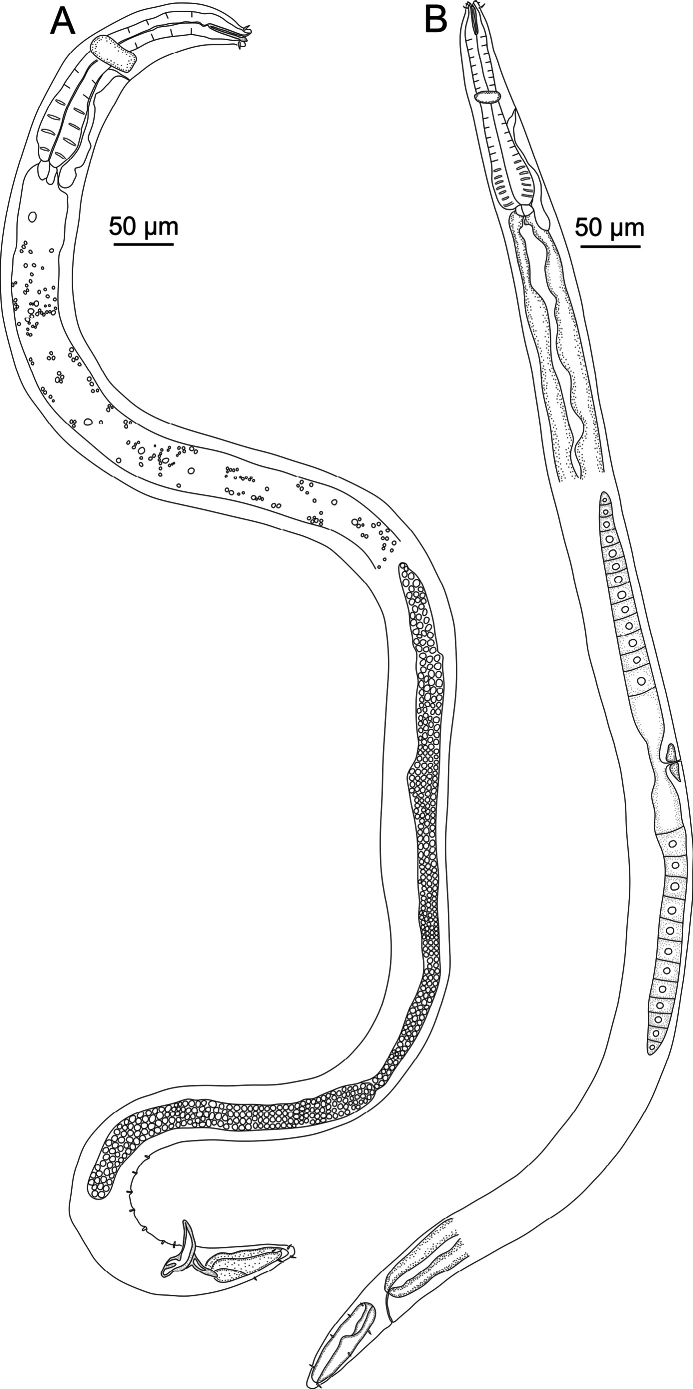 Figure 10
Figure 10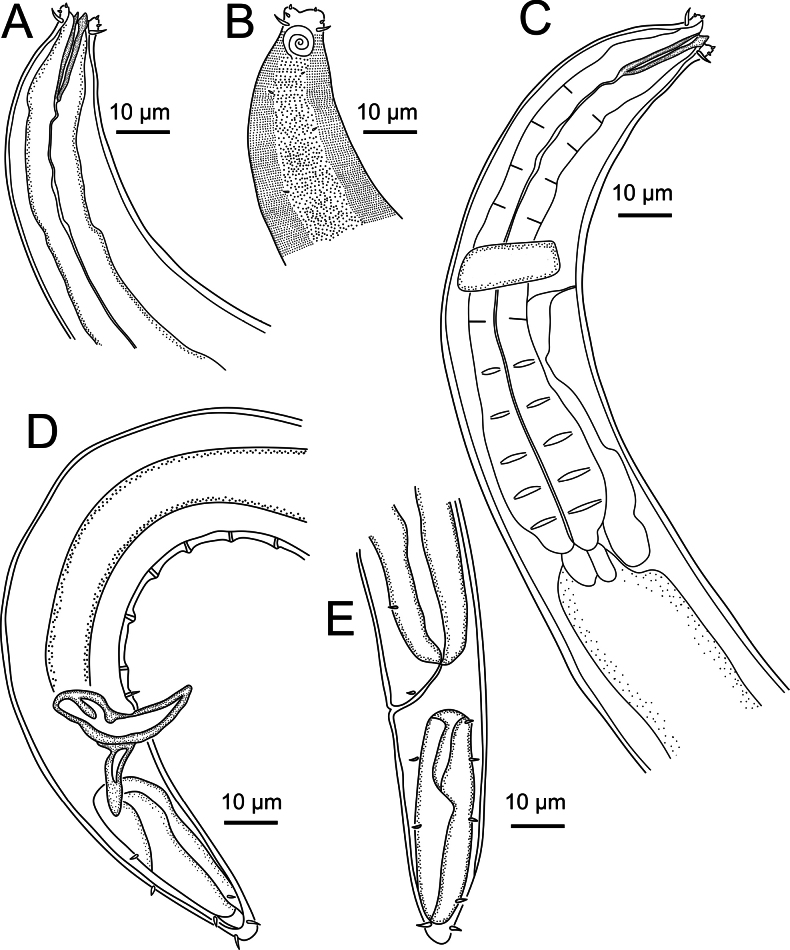 Figure 11
Figure 11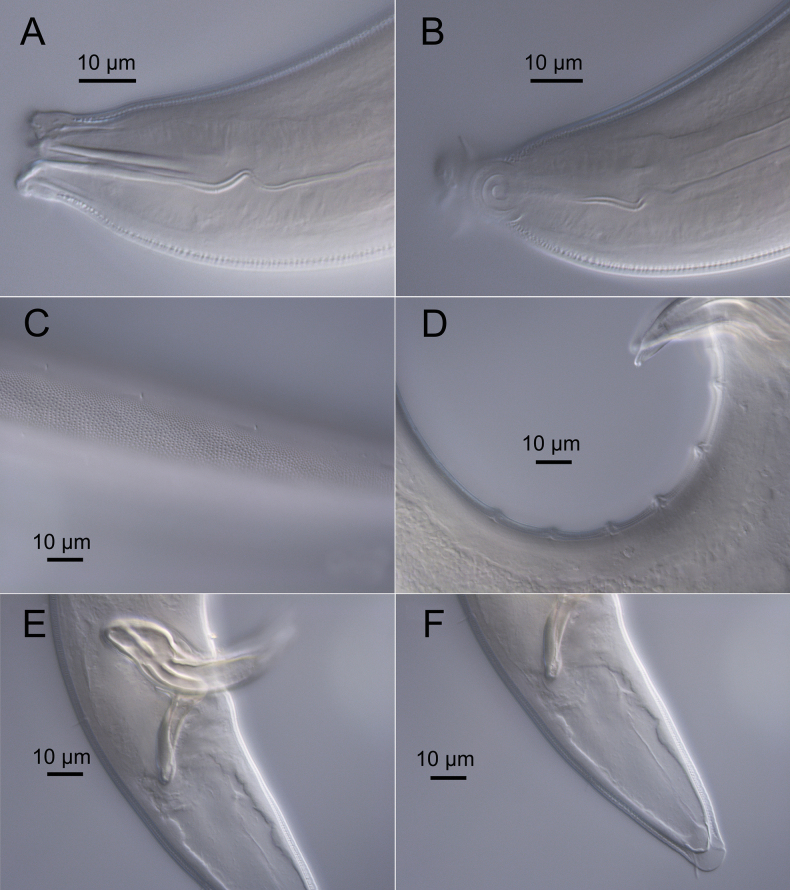 Figure 12
Figure 12 Figure 13
Figure 13| Station | Latitude, Longitude | Region | Date | Depth (m) |
|---|---|---|---|---|
| DA-04 |
| Antarctic Peninsula | 28 Dec. 2022 | 574 |
| C4-12 |
| Cosmonauts Sea | 22 Jan. 2021 | 1321 |
| C4-11 |
| Cosmonauts Sea | 18 Jan. 2021 | 2085 |
| C4-10 |
| Cosmonauts Sea | 20 Jan. 2021 | 2444 |
| C5-07 |
| Cosmonauts Sea | 20 Feb. 2022 | 2096 |
| C2-13 |
| Cosmonauts Sea | 26 Jan. 2021 | 1186 |
| M10D |
| Prydz Bay | 31 Dec. 2020 | 578 |
| MF1 |
| Prydz Bay | 1 Jan. 2021 | 310 |
| A2-2 |
| Amundsen Sea | 7 Feb. 2022 | 457 |
| A11-00 |
| Amundsen Sea | 5 Feb. 2022 | 773 |
| Male holotype | Male paratypes | Female paratypes | |
|---|---|---|---|
| Number of specimens | 1 | 4 | 3 |
| L | 2230 | 1898 (1786–2009) | 2123 (2032–2224) |
| a | 35 | 29 (27–31) | 35 (29–39) |
| b | 9 | 8 (7–8) | 8 |
| c | 18 | 16 (15–18) | 18 (17–20) |
| c′ | 2.4 | 2.2 (1.9–2.4) | 2.5 (2.4–2.6) |
| Head diameter at cephalic setae | 14 | 16 (15–18) | 15 (14–16) |
| Length of cephalic setae | 4 | 3 | 3 (3–4) |
| Amphid height | 13 | 12 (12–14) | 11 (11–12) |
| Amphid width | 12 | 12 (10–12) | 11 (10–11) |
| Amphid width/ | 66 | 62 (54–68) | 51 (48–54) |
| Amphid from anterior end | 9 | 9 (8–10) | 10 (9–12) |
| Nerve ring from anterior end | 142 | 109 (94–125) | 112 (102–128) |
| Nerve ring | 45 | 44 (41–47) | 43 (41–45) |
| Excretory pore from anterior end | 154 | 143 (126–162) | 166 (149–184) |
| Pharynx length | 254 | 250 (239–261) | 264 (254–269) |
| Pharynx bulb | 54 | 55 (50–59) | 53 (52–55) |
| Pharyngeal bulb diameter | 30 | 34 (33–35) | 38 (37–39) |
| Maximum body diameter | 63 | 65 (59–73) | 62 (57–69) |
| Spicule length | 76 | 76 (71–79) | - |
| Gubernacular apophyses length | 26 | 24 (21–29) | - |
| Number of precloacal supplements | 17 | 16 or 17 | - |
| Anal or cloacal body diameter | 50 | 53 (50–58) | 46 (46–47) |
| Tail length | 122 | 116 (110–124) | 116 (113–118) |
| V | - | - | 1046 (1005–1082) |
| V% | - | - | 49 (49–50) |
| Vulva body diameter | - | - | 61 (57–69) |
| Male holotype | Male paratypes | Female paratypes | |
|---|---|---|---|
| Number of specimens | 1 | 3 | 3 |
| L | 3093 | 2888 (2628–3085) | 3176 (2910–3613) |
| a | 30 | 28 (23–31) | 27 (24–33) |
| b | 9 | 8 (8) | 8 (8) |
| c | 16 | 17 (16–18) | 17 (16–18) |
| c′ | 2.7 | 2 (1.8–2.1) | 2.3 (2.1–2.4) |
| Head diameter at cephalic setae | 20 | 21 (18–23) | 24 (22–26) |
| Length of cephalic setae | 4 | 5 (4–6) | 5 (4–5) |
| Amphid height | 12 | 12 (12–13) | 12 (11–12) |
| Amphid width | 13 | 12 (10–13) | 12 (12–13) |
| Amphid width/ | 54 | 48 (38–54) | 46 (44–47) |
| Amphid from anterior end | 10 | 10 (9–11) | 11 (9–12) |
| Nerve ring from anterior end | 139 | 158 (146–178) | 168 (134–226) |
| Nerve ring | 69 | 76 (71–84) | 76 (74–78) |
| Excretory pore from anterior end | 196 | 219 (199–230) | 232 (199–268) |
| Pharynx length | 341 | 347 (311–367) | 390 (355–443) |
| Pharynx bulb | 99 | 94 (87–104) | 99 (94–102) |
| Pharyngeal bulb diameter | 62 | 61 (57–64) | 68 (62–74) |
| Maximum body diameter | 102 | 105 (84–136) | 117 (110–127) |
| Spicule length | 108 | 107 (103–110) | - |
| Gubernacular apophyses length | 47 | 44 (42–45) | - |
| Number of precloacal supplements | 21 | 20 or 21 | - |
| Anal or cloacal body diameter | 70 | 88 (84–94) | 84 (83–86) |
| Tail length | 190 | 174 (169–182) | 191 (184–199) |
| V | - | - | 1609 (1441–1870) |
| V% | - | - | 51 (48–52) |
| Vulva body diameter | - | - | 119 (117–120) |
| Males | Female | |
|---|---|---|
| Number of specimens | 3 | 1 |
| L | 1618–1760 | 1341 |
| a | 20–24 | 25 |
| b | 7-8 | 6 |
| c | 18–21 | 15 |
| c′ | 2 | 3 |
| Head diameter at cephalic setae | 14–16 | 13 |
| Length of cephalic setae | 4–5 | 5 |
| Amphid height | 11–12 | 9 |
| Amphid width | 11–12 | 8 |
| Amphid width/ | 62–69 | 54 |
| Amphid from anterior end | 7 | 5 |
| Nerve ring from anterior end | 96–109 | 76 |
| Nerve ring | 50–53 | 36 |
| Excretory pore from anterior end | 130–134 | 131 |
| Pharynx length | 226–231 | 211 |
| Pharynx bulb | 56–82 | 47 |
| Pharyngeal bulb diameter | 37–60 | 33 |
| Maximum body diameter | 71–85 | 54 |
| Spicule length | 54–58 | - |
| Gubernacular apophyses length | 25–27 | - |
| Anal or cloacal body diameter | 42–43 | 36 |
| Tail length | 80–94 | 89 |
| V | - | 678 |
| V% | - | 51 |
| Vulva body diameter | - | 54 |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 0.0044 | 0.1464 | 0.0044 | 0.0015 | 0.1252 | 0.1118 | 0.1426 | 0.1426 | 0.1349 | 0.1040 | 0.2177 | 0.2505 | 0.2582 | 0.2582 | ||
| 2 | 0.1079 | 0.1252 | 0.0000 | 0.0983 | 0.0559 | 0.0809 | 0.1310 | 0.1310 | 0.1310 | 0.1214 | 0.2235 | 0.2678 | 0.2177 | 0.2177 | ||
| 3 | 0.0116 | 0.0073 | 0.0073 | 0.0102 | 0.0087 | 0.0116 | 0.0058 | 0.0058 | 0.0087 | 0.1561 | 0.2216 | 0.2563 | 0.2158 | 0.2158 | ||
| 4 | 0.1021 | 0.0520 | 0.1233 | 0.0906 | 0.0405 | 0.0771 | 0.1118 | 0.1118 | 0.1195 | 0.1137 | 0.2274 | 0.2543 | 0.2023 | 0.2023 | ||
| 5 | 0.0347 | 0.0029 | 0.1291 | 0.0029 | 0.1156 | 0.0867 | 0.1387 | 0.1387 | 0.1426 | 0.1002 | 0.2100 | 0.2524 | 0.2524 | 0.2524 | ||
| 6 | 0.0087 | 0.0044 | 0.1195 | 0.0044 | 0.0073 | 0.0944 | 0.1214 | 0.1214 | 0.1252 | 0.1329 | 0.2274 | 0.2640 | 0.1965 | 0.1965 | ||
| 7 | 0.0087 | 0.0073 | 0.1233 | 0.0073 | 0.0073 | 0.0116 | 0.1195 | 0.1195 | 0.1272 | 0.1040 | 0.2351 | 0.2717 | 0.2312 | 0.2312 | ||
| 8 | 0.0116 | 0.0073 | 0.0925 | 0.0073 | 0.0102 | 0.0087 | 0.0116 | 0.0000 | 0.0385 | 0.1541 | 0.2370 | 0.2601 | 0.2447 | 0.2447 | ||
| 9 | 0.0116 | 0.0073 | 0.0925 | 0.0073 | 0.0102 | 0.0087 | 0.0116 | 0.0000 | 0.0385 | 0.1541 | 0.2370 | 0.2601 | 0.2447 | 0.2447 | ||
| 10 | 0.0116 | 0.0102 | 0.0809 | 0.0102 | 0.0102 | 0.0116 | 0.0087 | 0.0058 | 0.0058 | 0.1618 | 0.2351 | 0.2640 | 0.2274 | 0.2274 | ||
| 11 | 0.0102 | 0.0087 | 0.0160 | 0.0087 | 0.0087 | 0.0116 | 0.0131 | 0.0131 | 0.0131 | 0.0131 | 0.2216 | 0.2582 | 0.2331 | 0.2331 | ||
| 12 | 0.0247 | 0.0203 | 0.0247 | 0.0203 | 0.0233 | 0.0233 | 0.0218 | 0.0218 | 0.0218 | 0.0247 | 0.0203 | 0.2832 | 0.2678 | 0.2678 | ||
| 13 | 0.0305 | 0.0320 | 0.0349 | 0.0320 | 0.0291 | 0.0363 | 0.0334 | 0.0363 | 0.0363 | 0.0363 | 0.0378 | 0.0465 | 0.3295 | 0.3295 | ||
| 14 | 0.0378 | 0.0363 | 0.0407 | 0.0363 | 0.0363 | 0.0349 | 0.0378 | 0.0363 | 0.0363 | 0.0378 | 0.0422 | 0.0480 | 0.0596 | 0.0000 | ||
| 15 | 0.0276 | 0.0262 | 0.0305 | 0.0262 | 0.0262 | 0.0247 | 0.0276 | 0.0262 | 0.0262 | 0.0276 | 0.0320 | 0.0378 | 0.0494 | 0.0116 | ||
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine Biology and Ecology Research · Crustacean biology and ecology · Isotope Analysis in Ecology
Introduction
The family Comesomatidae Filipjev, 1918 is found from the intertidal zone and shallow sea areas to the deep sea, showing high dominance in marine sediments (Jensen 1979). The genus Sabatieria de Rouville, 1903 is one of the most common genera in Antarctica (Baishnab et al. 2023). Free-living marine nematodes have been used as indicators of biological, environmental, or physical disturbance, and Sabatieria is among the most tolerant genera to environmentally damaging factors (Ridall and Ingels 2021). Eighty-two valid Sabatieria species have been described to date; they are divided into five species groups, including the praedatrix group (50 species), the armata group (7 species), the pulchra group (9 species), the celtica group (7 species), the ornata group (7 species) and two ungrouped species (Platt 1985; Yang et al. 2019; Fu et al. 2023; Leduc and Zhao 2023). Praedatrix species group: most species with lateral cuticle differentiation of larger, more widely spaced punctations and amphids with three turns, but two or four turns may also occur. Spicules without central lamella distinct from the internal projection from proximal end. Simple tubular or pore-like supplements and straight gubernacular apophyses. This large group is relatively loosely defined, and unlike the other groups it is not characterized by any autapomorphic features. Armata species group: similar to praedatrix group except for the elongated cephalic setae (> 1.7 corresponding body diameter) and mostly slender bodies (a > 65). Amphids usually with three turns but four turns may also occur. Simple tubular supplements. Pulchra species group: pairs of short and relatively stout cervical setae present. Amphids with three to four turns. Five to nine conspicuous precloacal supplements. Gubernaculum with median pieces. Celtica species group: amphids with three turns. Body relatively large and stout. All have curved gubernacular apophyses, and conspicuous precloacal supplements. Ornata species group: amphids usually with three turns. Similar to celtica group except for the presence of a posterior group of more closely situated precloacal supplements (Platt 1985).
Studies on nematodes in the Southern Ocean began at the end of the 19^th^ century (von Linstow 1891). To date, more than 160 species of marine nematodes have been described; however, data on marine nematode taxonomic information in the Southern Ocean is still fragmentary (Allgén 1959; Platt 1983; Ingels et al. 2014; Leduc 2014; Leduc 2016; Shimada et al. 2017, 2019, 2021; Fu et al. 2023). Technological advances, including scanning microscopy and analysis of molecular data, have enhanced our understanding of biodiversity and the phylogenetic position of marine nematodes (Fonseca et al. 2017). In the present study, we describe two new Sabatieria species and a new record of Hopperiabeaglense Chen & Vincx, 1998 from the Southern Ocean using morphological and molecular data of 18S rRNA and D2-D3 expansion fragments of 28S rRNA. The new species and new record were found during a marine nematode diversity study resulting from the 37^th^ and 38^th^ Chinese Antarctic Research Expedition.
Material and methods
Sampling and sample processing
Sampling was conducted in the Antarctic Peninsula, the Cosmonauts Sea, Prydz Bay and Amundsen Sea during the 37^th^ and 38^th^ Chinese National Antarctic Research Expedition in the summer of 2021 and 2022 (Fig. 1, Table 1). As mentioned in our previous paper (Fu et al. 2023), the nematodes were sampled in the field, sorted, and slide-mounted in the laboratory.
Map of sampling stations in the Southern Ocean. Green circles indicate sampling stations and red text indicates station names.
The observations were conducted using differential interference contrast microscopy (LEICA DM6B), and line drawings were made with a camera lucida. For scanning electron microscopy (SEM), specimens were placed in a 2% osmium tetroxide solution overnight, then dehydrated through a graded ethanol series, and critical point drying. Finally, individuals were mounted onto stubs before coating with gold using a sputter coater. Observations were made using a focused ion beam SEM (Helios 5 UC) at Xiamen University.
All measurements are given in μm and curved structures measured along the arc. All holotypes and paratypes are deposited in the Marine Biological Sample Museum of the Chinese Offshore Investigation and Assessment (MBSMCOIA) at the Third Institute of Oceanography, Ministry of Natural Resources, China.
Abbreviations are as follows: a, body length/maximum body diameter; b, body length/pharynx length; c, body length/tail length; c’, tail length/cloacal body diameter; cbd, corresponding body diameter; L, total body length; n, number of specimens; V, vulva distance from anterior end of body; V%, V/total body length × 100.
DNA extraction, PCR amplification, and sequencing
Sediment samples used for molecular analysis were stored at -20 °C. Specimens of the two new species and one record were mounted on separate temporary slides to be photographed for morphological identification. The DNA extraction of each specimen was done using the Animal Genomic DNA Quick Extraction Kit for PCR analysis produced by Beyotime (Shanghai, China). Two fragments of 18S rRNA and 28S rRNA genes were amplified. For 18S rRNA of Sabatieriacosmonautae sp. nov. and Sabatieriacrassilonga sp. nov., 18SFi (5’-TGAATAACTACGCCGATCGCA-3’), and the reverse primer 18Sri (5’-CGAGCTTATGACCCACACTTACTG-3’) were designed using Primer Premier 5. For 28S rRNA, D2A (5′-ACAAGTACCGTGAGGGAAAGT-3′) and the reverse primer D3B (5′-TGCGAAGGAACCAGCTACTA-3′) were used following Nunn (1992). For both 18S rRNA and 28S rRNA, we used the 25-μL polymerase chain reaction (PCR) based on the method of our previous paper (Fu et al. 2023). Sequencing reactions were performed by Sangon Biotech (Shanghai) Co., Ltd. (Shanghai, China).
Sequence alignment and phylogenetic analyses
The DNA sequences of Sabatieriacosmonautae sp. nov. (18S rRNA and 28S rRNA), Sabatieriacrassilonga sp. nov. (18S rRNA and 28S rRNA) and Hopperiabeaglense were deposited in GenBank and the accession numbers are PP693307 and PP693308, PP693305 and PP693306, PP693309 and PP693358, respectively. Sequences used in the phylogenetic analysis were based on a single specimen of three species from three genera in the family Axonolaimidae, Filipjev, 1918 (outgroup) and 13 species from five genera in the family Comesomatidae, which were longer than 900 bp for 18S rRNA and 600 bp for 28S rRNA.
PhyloSuite (Zhang et al. 2020) was utilized to conduct the analyses using the following programs: sequences were aligned using MAFFT (Katoh et al. 2002) with default parameters; TrimAl v. 1.2 (Capella-Gutiérrez et al. 2009) was used to remove ambiguously aligned sequences with default settings; and ModelFinder (Kalyaanamoorthy et al. 2017) was used to select the best-fit model based on the corrected Akaike information criterion (AICc) (Sugiura 1978).
Phylogenetic analyses were carried out using Bayesian inference (BI) methods and maximum likelihood (ML) methods. Bayesian inference phylogenies were inferred using MrBayes v.3.2.6 (Ronquist et al. 2012). The best-fit model for the two new species and one new record for 18S rRNA was found to be GTR+F+I+G4, with two parallel runs for 2000000 generations; for 28S rRNA, it was under GTR+F+G4 with two parallel runs for 2000000 generations. Maximum likelihood phylogenies were performed using IQ-TREE (Nguyen et al. 2015). The best-fit model for the two new species and one new record for 18S rRNA was found to be under TIM2e+I+I+R2 for 10000 bootstraps; for 28S rRNA, it was under TIM3e+R2 for 10000 bootstraps. The results were viewed and embellished in iTOL v.6 (Letunic and Bork 2016). Genetic distances (p-distance) over sequence pairs were calculated in MEGA X (Kumar et al. 2018).
Results
Systematics
Family Comesomatidae Filipjev, 1918
Subfamily Sabatieriinae Filipjev, 1934
Sabatieria
Taxon classificationAnimaliaAraeolaimidaComesomatidae
Genus
de Rouville, 1903
4F42972F-7F22-5AFE-8608-1D79294AEF1D
Diagnosis
(modified from Fonseca and Bezerra (2014)). Cuticle with transverse punctation, lateral differentiation of larger regular or irregular punctations may occur. Cephalic sensilla in three distinct circles, cephalic setae longer than the outer labial setae. Anterior buccal cavity cup-shaped, posterior portion narrow, weakly cuticularized. Amphid multi-spiral. Apophyses usually directed dorso-caudally or caudally. Precloacal supplements usually present.
Sabatieria
cosmonautae
sp. nov.
Taxon classificationAnimaliaAraeolaimidaComesomatidae
07A15E44-0C79-530F-B4F9-CC26CB4A8587
https://zoobank.org/A4EFBA85-06CF-4C0E-9900-218F34095E86
Material examined.
Holotype: • male, collected in the Cosmonauts Sea; 67.1217°S, 44.3743°E; depth 1321 m; collected on 22 Jan. 2021; Jianfeng Mou leg.; total organic carbon 0.22%; sand, 50.73%; clay, 7.62%; silt 41.11%; mean particle diameter 4.49 mm; on slide no. MBSMCOIA-C4-12-0-1-2. Paratypes: • four males and three females, collected in the Cosmonauts Sea; 65.3262°S–67.1217°S, 44.3743°E–49.9987°E; depth 1321–2444 m; collected on 18 Jan. 2021–22 Jan. 2021, Feb. 2022; Jianfeng Mou leg.; total organic carbon 0.22%–0.61%; sand, 4.78%–50.73%; clay, 7.62%–23.70%; silt, 41.11%–71.51%; mean particle diameter 4.49–6.77 mm; males on slide no. MBSMCOIA-C5-7-0-2-2, MBSMCOIA-C4-10-0-2-2, MBSMCOIA-C4-11-2-4 and MBSMCOIA-C4-12-0-1; females on slide no. MBSMCOIA-C4-11-2-4.
Measurements.
All measurement data are given in Table 2.
Description.
Male. Body short, narrowing gradually towards both extremities. Cuticle with transverse rows of punctations starting from amphid to near tail tip. Lateral differentiation with slightly sparse punctations. Cuticle striations distinct on cuticle surface using SEM (Fig. 5). Short somatic setae sparsely present on entire body, ~2 μm long. Cephalic region with distinct constriction at level of cephalic setae. Anterior sensilla in three crowns: six small inner labial papillae, six outer labial setae (2 μm long), and four cephalic setae (3 μm long or 0.15–0.25 corresponding body diameter long). Anterior buccal cavity cup-shaped, 5–6 μm wide, posterior portion narrow. Spiral amphidial fovea with three turns, 10–12 μm in diameter or 54–68% of corresponding body diameter, anterior border located at the level of cephalic setae. Pharynx surrounds half of anterior buccal cavity, gradually broadening posteriorly and forming a weak posterior bulb. Nerve ring situated at 39–56% of anterior pharynx. Secretory-excretory gland located posterior to pharynx. Secretory-excretory pore located posterior to nerve ring, 126–162 μm from the anterior end. Cardia small, completely surrounded by intestinal tissue. Reproductive system diorchic, anterior outstretched testis to the left of intestine and posterior outstretched testis to the right of the intestine. Spicules arcuate, equal, 1.2–1.5 cloacal body diameters long, proximal part of spicule with central cuticularized projection (lamella) extending from proximal end to approximately half of spicule. Gubernaculum with straight dorso-caudally directed apophyses. One precloacal seta 2 μm long. Sixteen or seventeen precloacal supplements, in the form of small pores, with distance between the anterior eight supplements (12–21 μm) irregularly distributed, posterior eight supplements with distance between adjacent supplements (5–13 μm) increasing towards anterior (Fig. 3D). Conico-cylindrical tail 1.9–2.4 cloacal body diameters long bearing three terminal setae, with very slightly swollen tip. Cylindrical part ~1/5 of total tail length. Three caudal glands present. Spinneret present.
Sabatieriacosmonautae sp. nov. A. Entire male holotype; B. Entire female paratype.
Sabatieriacosmonautae sp. nov. A. Male holotype anterior body region; B. Surface view of male holotype anterior body region showing amphidial fovea and cephalic sensilla; C. Male holotype pharyngeal body region; D. Male holotype posterior body region, showing precloacal supplements (arrow); E. Female paratype posterior body region.
Sabatieriacosmonautae sp. nov. Light micrographs. A. Male paratype anterior body region showing the buccal cavity; B. Surface view of female paratype anterior body region showing amphids; C. Lateral punctations of male paratype anterior body region; D. Vulva; E. Male paratype posterior body region; F. Male paratype tail.
Sabatieriacosmonautae sp. nov. Scanning electron micrographs. A. Male anterior body region showing cephalic sensilla; B. Male posterior body region.
Female. Similar to males. Reproductive system with two opposed, outstretched ovaries, with anterior ovary to the left of intestine and posterior ovary to the right of intestine. Spermatheca present. Vulva at 49–50% of body length from anterior. Granular vaginal glands present. Three caudal glands present.
Diagnosis and relationships.
Sabatieriacosmonautae sp. nov. is characterized by body length 1786–2230 μm, short cephalic setae, 0.15–0.25 corresponding body diameter long; cuticle with transverse rows of punctations starting from amphid to near tail tip; lateral differentiation with sparse punctations; spiral amphidial fovea with three turns; males with arcuate spicules, 1.2–1.5 cloacal body diameters long, straight dorso-caudally directed gubernacular apophyses, and with 16 or 17 precloacal supplements, in the form of small pores; Conico-cylindrical tail 1.9–2.4 cloacal body diameters long in males and 2.4–2.6 cloacal body diameters long in females.
Sabatieriacosmonautae sp. nov. belongs to the praedatrix group: amphids with three turns, the presence of straight gubernacular apophyses and with 16 or 17 pore-like precloacal supplements. There are 50 valid species in this group. Within the praedatrix group, S. cosmonautae sp. nov. most closely resembles S.ancudiana Wieser, 1954, S.granifer Wieser, 1954, S.intermissa Wieser, 1954, S.lawsi Platt, 1983, S.parabyssalis Wieser, 1954 as well as S.praedatrix de Man, 1907, which are all characterized by having nearly 17 precloacal supplements. In addition, the new species differs from S.ancudiana in the stouter body shape (a = 27–35 vs 42–65 in S.ancudiana), shorter cephalic setae (0.15–0.25 vs 0.60–0.70 cbd in S.ancudiana) and shorter tail (1.9–2.4 vs 3.5 cloacal body diameters in S.ancudiana), from S.granifer in the shorter tail (1.9–2.4 vs 3.3–4.8 cloacal body diameters in S.granifer), from S.intermissa in the stouter body shape (a = 27–35 vs 40–48 in S.intermissa), shorter cephalic setae (0.15–0.25 vs 0.70–0.90 cbd in S.intermissa), shorter spicules (1.2–1.5 vs 2.0 cloacal body diameters in S.intermissa) and shorter tail (1.9–2.4 vs 3.5 cloacal body diameters in S.intermissa), from S.lawsi in the shorter tail (1.9–2.4 vs 3.1–3.8 cloacal body diameters in S.lawsi), from S.parabyssalis in the shorter cephalic setae (0.15–0.25 vs 0.70 cbd in S.parabyssalis), and shorter tail (1.9–2.4 vs 3.8–4.3 cloacal body diameters in S.parabyssalis), and from S.praedatrix in the much shorter tail (1.9–2.4 vs 4.0–4.5 cloacal body diameters in S.praedatrix).
This new species is also similar to S.bathycopia Leduc, 2013 in the celtica group by the relative cephalic setae length, relative spicule length and the value of c'. The new species differs from S.bathycopia by the value of c' (1.9–2.4 vs 3.6–4.3 in S.bathycopia), the structure of spicules (without velum or swollen distal portion vs with velum and swollen distal portion in S.bathycopia), and the gubernaculum shape (straight vs curved in S.bathycopia).
Etymology.
This species is named after the type locality belonging to the Cosmonauts Sea.
Sabatieria
crassilonga
sp. nov.
Taxon classificationAnimaliaAraeolaimidaComesomatidae
5C595496-BF9F-566D-B9E1-712B93EFCC7F
https://zoobank.org/5F2C0763-5B31-4041-91C7-CD08224DA181
Material examined.
Holotype: • male, collected in the Amundsen Sea, 72.7167°S, 124.5031°W; depth 457 m; collected on 7 Feb. 2022; Jianfeng Mou leg.; muddy sediment; on slide no. MBSMCOIA-A2-2-2-5. Paratypes: • three males and three females, collected in the Cosmonauts Sea, Prydz Bay and Amundsen Sea; 66.5038°S–73.9433°S, 33.2992°E–112.3525°W; depth 310-2085 m; collected on 1 Jan. 2021–22 Jan. 2021, Feb. 2022; Jianfeng Mou leg.; total organic carbon 0.12%–0.61%; sand, 4.78%–55.92%; clay, 7.62%–23.70%; silt, 35.58%–71.51%; mean particle diameter 4.36–6.77 mm; males on slide no. MBSMCOIA-C2-13-2-5-3, MBSMCOIA-A11-00-5-10-3, and MBSMCOIA-MF1-2-2; females on slide no. MBSMCOIA-MF1-2-2, MBSMCOIA-C4-11-2-4, and MBSMCOIA-C4-12-0-1.
Measurements.
All measurement data are given in Table 3.
Description.
Male. Body long (2628–3093 μm long), quite stout (maximum body width 84–136 μm), narrowing gradually towards both extremities. Cuticle with lateral differentiation of coarser, irregularly spaced punctations starting from amphid to near tail tip. Cuticle striations appearing distinct on cuticle surface under SEM (Fig. 9). Short somatic setae sparsely present on entire body, ~3 μm long. Cephalic region with distinct constriction at level of cephalic setae. Anterior sensilla in three crowns: six small inner labial papillae, six outer labial setae (3 μm long), and four cephalic setae (4–6 μm long or 0.20–0.26 corresponding body diameter long). Anterior buccal cavity cup-shaped, 4–7 μm wide, posterior portion narrow. Spiral amphidial fovea with 2.5 turns, 10–13 μm in diameter or 38–54% of corresponding body diameter, anterior border located at the level of cephalic setae. Pharynx surrounds half of anterior buccal cavity, gradually broadening posteriorly and forming a weak posterior bulb. Nerve ring situated at 40–49% of anterior pharynx. Secretory-excretory gland located posterior to pharynx. Secretory-excretory pore located posterior to nerve ring, 196–230 μm from the anterior end. Cardia small, partially surrounded by intestinal tissue. Reproductive system diorchic, anterior outstretched testis to the left of intestine and posterior outstretched testis to the left or right of the intestine. Spicules arcuate, equal, 1.2–1.5 cloacal body diameters long, proximal part of spicule with central cuticularized projection (lamella) extending from proximal end to ~1/3 of spicule. Gubernaculum with a long straight dorso-caudal apophysis, 42–47 μm long. One precloacal seta 3 μm long, distinct using SEM (Fig. 9C). Twenty or twenty-one precloacal supplements, in the form of small pores, the distance between seven posterior-most supplements more or less equal (10–12 μm), anterior nine supplements with distance between adjacent supplements (12–36 μm) increasing towards anterior (Figs 7, 9). Conico-cylindrical tail 1.8–2.7 cloacal body diameters long with swollen tip bearing three terminal setae. Cylindrical part ~1/5 of total tail length. Three caudal glands present. Spinneret present.
Sabatieriacrassilonga sp. nov. A. Entire male holotype; B. Entire female paratype.
Sabatieriacrassilonga sp. nov. A. Male holotype anterior body region; B. Surface view of male holotype anterior body region showing amphidial fovea and cephalic sensilla; C. Male holotype pharyngeal body region; D. Male holotype posterior body region, showing precloacal supplements (arrow); E. Female paratype posterior body region.
Sabatieriacrassilonga sp. nov. Light micrographs. A. Male paratype anterior part showing buccal cavity; B. Surface view of male paratype anterior body region showing amphids; C. Lateral punctations of female paratype anterior body region; D. Female paratype vulva; E. Male paratype spicule; F. Male paratype gubernaculum.
Sabatieriacrassilonga sp. nov. Scanning electron micrographs. A. Male anterior part showing cephalic sensilla and amphids; B. Male anterior part showing amphids; C. Male posterior part showing cloaca, precloacal seta (right arrow) and precloacal supplements (left arrow); D. Male posterior body region.
Female. Similar to males. Reproductive system with two opposed, outstretched ovaries, with anterior ovary to the left of intestine and posterior ovary to the left or right of intestine. Spermatheca present. Vulva at 48–52% of body length from anterior. Granular vaginal glands present. Three caudal glands present.
Diagnosis and relationships.
Sabatieriacrassilonga sp. nov. is characterized by relatively stout body (a = 23–33); long body length 2628–3093 μm in males and 2910–3613 μm in females; short cephalic setae (4–6 μm long, 0.20–0.26 corresponding body diameter long); cuticle with lateral differentiation of coarser and irregularly spaced punctations extending from anterior edge of amphid to the tail region; amphidial fovea spiral with 2.5 turns; males with spicules 1.2–1.5 cloacal body diameters long. Twenty or twenty-one fine precloacal supplements. Gubernaculum with a long straight dorso-caudal apophysis, 42–47 μm long. Conico-cylindrical tail 1.8–2.7 cloacal body diameters long in male and 2.1–2.4 cloacal body diameters long in female.
Sabatieriacrassilonga sp. nov. belongs to the praedatrix group based on amphids with three turns, the presence of pore-like supplements and straight apophysis. An amphid with 2¼, 2½, 2¾, or 3 turns are all considered as three turns (Platt 1985). Within the praedatrix group, Sabatieriacrassilonga sp. nov. most resembles S.alata Warwick, 1973, S.coomansi Chen & Vincx, 1999, S.major Yang, Guo, Chen & Lin, 2019, S.triplex Wieser, 1954, and S.palmaris Fadeeva & Belogurov, 1984 in having long body length (2600–4012 μm). The new species can be differentiated from S.alata by the ratio of a (23–31 vs 36–53 in S.alata) and shorter tail (2.1–2.4 vs 5.5 cloacal body diameters in S.alata), from S.coomansi by the number of precloacal supplements (20 or 21 vs 23–26 in S.coomansi) and shorter tail (2.1–2.4 vs 3.3–3.6 cloacal body diameters in S.coomansi), from S.major by the shorter body length (2888–3613 μm vs 3879–4255 μm in S.major) and the lower value of c’ (1.8–2.7 vs 2.9–3.7 in S.major), from S.triplex by the ratio of a (23–31 vs 57 in S.triplex), length of the cephalic setae (0.20–0.26 vs 0.50 cbd in S.triplex) and shorter tail (2.1–2.4 vs 3.7 cloacal body diameters in S.triplex), and from S.palmaris by the ratio of a (23–31 vs 20–22 in S.palmaris) and the ratio of c (16–18 vs 9–11 in S.palmaris).
Sabatieriacrassilonga sp. nov. is also similar to S.kelletti Platt, 1983 in the celtica group but differs from the latter by the shorter cephalic setae (0.20–0.26 vs 0.42–0.47 cbd in S.kelletti), lower number of precloacal supplements (20 or 21 vs 21–27 in S.kelletti) and the lower value of c’ (1.8–2.7 vs 3.6–4.3 in S.kelletti).
Etymology.
The species is named due to its stout and long body, which derived from Latin crassa (= thick, fat, stout) and longa (= long).
Hopperia
beaglense
Taxon classificationAnimaliaAraeolaimidaComesomatidae
Chen & Vincx, 1998
E6619A56-CB32-56FB-AF11-B153E5D0178B
Material examined.
• Three males, collected in the Prydz Bay, 68.2833°S, 75.5827°E; depth 578 m; collected on 31 Dec. 2020; Jianfeng Mou leg.; total organic carbon 1.37%; sand, 7.87%; clay, 16.77%; silt 75.37%; mean particle diameter 6.20 μm; on slide no. MBSMCOIA-M10D-3-5-10, MBSMCOIA-M10D-1-2-5-4 and MBSMCOIA-M10D-1-5-10-1; female specimen was collected from Antarctic Peninsula, 61.0046°S, 54.6971°W; depth of 574 m; collected on 28 Dec. 2022; Jianfeng Mou leg.; muddy sediment; on slide no. MBSMCOIA-DA-04-2-5-1.
Measurements.
All measurement data are given in Table 4.
Description.
Male. Body long (1618–1760 μm long), narrowing gradually towards both extremities. Cuticle with transverse rows of punctations starting from amphid to near tail tip. Lateral differentiation with larger, irregularly spaced punctations. Short somatic setae sparsely present on entire body, ~3 μm long. Cephalic region with distinct constriction at level of cephalic setae. Anterior sensilla in three crowns: six small inner labial papillae, six outer labial setae (2 μm long), and four cephalic setae (4–5 μm long or 0.30–0.33 corresponding body diameter long). Anterior buccal cavity cup-shaped, and the posterior portion cylindrical. Three cuticularized pointed teeth present at the junction between the two compartments of buccal cavity. Spiral amphidial fovea with 3.5 turns, 11–12 μm in diameter or 62–69% of corresponding body diameter, anterior border located at the level of cephalic setae. Pharynx cylindrical, gradually broadening posteriorly and forming a weak posterior bulb. Nerve ring situated at 42–47% of anterior pharynx. Secretory-excretory gland located posterior to pharynx. Secretory-excretory pore located slightly posterior to nerve ring, 96–109 μm from the anterior end. Cardia small, partially surrounded by intestinal tissue. Reproductive system diorchic, anterior outstretched testis to the left or right of intestine and posterior outstretched testis to the right of the intestine. Spicules arcuate, equal, 1.3–1.4 cloacal body diameters long, proximal part of spicule with central cuticularized projection (lamella) extending from proximal end to ~1/4 of spicule. Gubernaculum with slightly curved dorso-caudal apophyses, 25–27 μm long. One precloacal seta 3 μm long. Seven tubular precloacal supplements. Conical tail 1.8–2.2 cloacal body diameters long, two terminal setae (4 μm long). Three caudal glands present.
Hopperiabeaglense Chen & Vincx, 1998. A. Entire male; B. Entire female.
Hopperiabeaglense Chen & Vincx, 1998. A. Male anterior body region; B. Surface view of male anterior body region showing amphidial fovea and cephalic sensilla; C. Male pharyngeal body region; D. Male posterior body region; E. Female posterior body region.
Female. Similar to males, but shorter than males. Reproductive system with two opposed, outstretched ovaries, with anterior ovary to the left of intestine and posterior ovary to the right of intestine. Vulva at 51% of body length from anterior. Granular vaginal glands present. Three caudal glands present.
Hopperiabeaglense Chen & Vincx, 1998. Light micrographs. A. Male anterior part showing buccal cavity; B. Surface view of male anterior body region showing amphids; C. Lateral punctations of female anterior body region; D. Male posterior part showing precloacal supplements; E. Male spicular apparatus; F. Female tail.
Remarks.
Hopperiabeaglense was first described by Chen and Vincx (1998) from the Strait of Magellan and the Beagle Channel, based on specimens collected at 100–110 m water depth with muddy sediment. The present specimens were collected from muddy sediment and at greater water depths (574–578 m). This species differs from all other Hopperia species by the conical tail tip. The present specimens are similar to the description by Chen and Vincx (1998), and there are some differences in the value of a (20–25 in present specimens vs 30–41 in the Chilean specimens), maximum body diameter in males (71–85 μm in present specimens vs 41–49 μm in the Chilean specimens), and the present specimens lack the supplement-like structure situated halfway down the ventral side of the tail. This species was also recorded in Kaikoura Canyon, New Zealand (Leduc 2012). So, H.beaglense has a circumpolar distribution.
Molecular analysis
The ML topology trees based on concatenated 18S + 28S rRNA sequences are similar to the BI topology, and only the BI trees are shown (Fig. 13). According to tree topology, 13 species from five genera within the family Comesomatidae, including Setosabatieria Platt, 1985, Comesoma Bastian, 1865, Sabatieria, Hopperia Vitiello, 1969 and Dorylaimopsis Ditlevsen, 1918, form a well-supported monophyletic clade (100% posterior probability and 100% bootstrap value). Genus Sabatieria is not shown as a monophyletic clade which is in accordance with Fu et al. (2019) and Fu et al. (2023) based on 18S rRNA and 28S rRNA sequences. Sabatieriacosmonautae sp. nov. was sister to Sabatieriabrevicaudata Fu, Zhang, Leduc, Mou & Lin, 2023 (100% posterior probability and 98% bootstrap value). Sabatieriacrassilonga sp. nov. was sister to the group of Sabatieriaarticulata Fu, Leduc & Zhao, 2019, Sabatieriamultipora Fu, Zhang, Leduc, Mou & Lin, 2023 and Dorylaimopsis sp. with high support (100% posterior probability and 97% bootstrap value). Hopperiabeaglense was sister to the clade consisting of Sabatieriapunctata (Kreis, 1924) and Sabatieriapulchra (Schneider, 1906) Riemann, 1970 with high support (100% posterior probability and 100% bootstrap value). Table 5 provides the pairwise p-distances for 18S and 28S rRNA sequences. Within the Comesomatidae, genetic distances between Sabatieriacosmonautae sp. nov. and other species ranged from 0.87% to 10.79% for 18S rRNA sequences, and from 0.15% to 25.82% for 28S rRNA; genetic distances between Sabatieriacrassilonga sp. nov. and other species ranged from 0.29% to 10.79% for 18S rRNA sequences, and from 0 to 26.78% for 28S rRNA sequences. Relationship analysis based on 18S rRNA sometimes contradicts the results from 28S rRNA (Liang et al. 2024), which makes determining the relationships among nematodes even more complex. Phylogenetic analysis based on concatenated 18S + 28S rRNA sequences might give more hints to solve relationships among nematodes. Besides accurate morphological identification, phylogenetic relationships based on concatenated rRNA sequence analysis requires obtaining multiple gene fragments from the same specimen. Due to the limited availability of gene data from the same specimen in GenBank, we could not confirm whether concatenated 18S + 28S rRNA sequence analysis is superior to the analysis of a single gene fragment. DNA barcoding is aiding in species identification nowadays, and more nuclear and mitochondrial genes are needed to obtain more reliable results in phylogenetic relationships of nematodes.
Bayesian inference tree of the families Comesomatidae and Axonolaimidae (outgroup) based on concatenated 18S + 28S rRNA sequences. Posterior probability on the left and bootstrap values on the right are given for the corresponding clades. Sequences obtained in this study are shown in bold. The scale bar represents the number of substitutions per site.
Supplementary Material
XML Treatment for Sabatieria
XML Treatment for Sabatieria cosmonautae
XML Treatment for Sabatieria crassilonga
XML Treatment for Hopperia beaglense
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Allgén CA (1959) Further Zoological Results of the Swedish Antarctic Expedition 1901–1903. Vol. V. No. 2. Freeliving Marine Nematodes. Kungl. Boktryckeriet P.A.Norstedt & Söner, Stockholm, 293 pp.
- 2Baishnab SS Shahir A Mandal S Tripathy SC (2023) Unveiling the meiobenthic community structure of Prydz bay, Antarctica during austral summer. Deep-sea Research. Part I, Oceanographic Research Papers 199: 104109. 10.1016/j.dsr.2023.104109 · doi ↗
- 3Bastian HC (1865) Monograph of the Anguillulidae, or Free Nematoids, Marine, Land, and Freshwater; with Descriptions of 100 New Species. Transactions of the Linnean Society of London 25(Part II): 73–184. 10.1111/j.1096-3642.1865.tb 00179.x · doi ↗
- 4Capella-Gutiérrez S Silla-Martínez JM Gabaldón T (2009) Trim Al: A tool for automated alignment trimming in large-scale phylogenetic analyses.Bioinformatics (Oxford, England)25(15): 1972–1973. 10.1093/bioinformatics/btp 34819505945 PMC 2712344 · doi ↗ · pubmed ↗
- 5Chen GT Vincx M (1998) Nematodes from the strait of Magellan and the Beagle channel (Chile): Description of four new species of the Comesomatidae.Hydrobiologia 379(1–3): 97–110. 10.1023/A:1003494127181 · doi ↗
- 6Chen GT Vincx M (1999) Nematodes from the Strait of Magellan and the Beagle Channel (Chile): the genus Sabatieria (Comesomatidae: Nematoda) with the description of Sabatieriacoomansi n. sp.Hydrobiologia 405: 95–116. 10.1023/A:1003752619953 · doi ↗
- 7de Man JG (1907) Sur quelques espèces nouvelles ou peu connues de nématodes libres habitant les côtes de la Zélande.Mémoires de la Société Zoologique de France 20: 33–90.
- 8de Rouville E (1903) Enumération des Nématodes libres du canal des Bourdigues (Cette).Comptes Rendus des Seances de la Société de Biologie et de ses Filiales 55: 1527–1529.