Early-life adversity alters adult nucleus incertus neurons: implications for neuronal mechanisms of increased stress and compulsive behavior vulnerability
Anna Gugula, Patryk Sambak, Aleksandra Trenk, Sylwia Drabik, Aleksandra Nogaj, Zbigniew Soltys, Andrew L. Gundlach, Anna Blasiak

TL;DR
Early-life stress changes brain cells in adult rats, increasing vulnerability to stress and compulsive behaviors.
Contribution
This study reveals how early-life adversity alters the nucleus incertus, a brain region linked to stress and reward.
Findings
ELS reduced acute stress responsiveness and caused dendritic shrinkage in nucleus incertus neurons.
ELS increased CRHR1 and TrkA mRNA expression in adult nucleus incertus neurons.
Synaptic transmission and electrophysiological properties of NI neurons were altered by ELS.
Abstract
Early-life stress (ELS) arising from physical and emotional abuse disrupts normal brain development and impairs hypothalamic–pituitary–adrenal axis function, increasing the risk of psychopathological disorders and compulsive behaviors in adulthood. However, the underlying neural mechanisms remain unclear. The brainstem nucleus incertus (NI) is a highly stress-sensitive locus, involved in behavioral activation and stress-induced reward (food/alcohol) seeking, but its sensitivity to ELS remains unexplored. We used neonatal maternal separation stress in rats as a model for ELS and examined its impact on stress-related mRNA and neuropeptide expression in the NI, using fluorescent in situ hybridization and immunohistochemistry, respectively. Using whole-cell, patch-clamp recordings we determined the influence of ELS on the synaptic activity, excitability, and electrophysiological properties…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
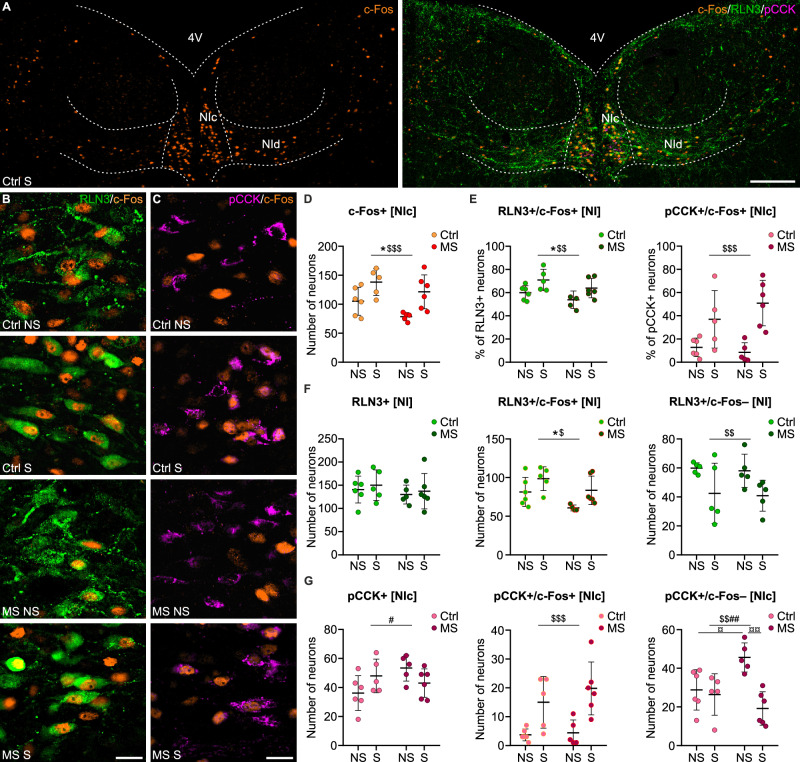 Figure 1
Figure 1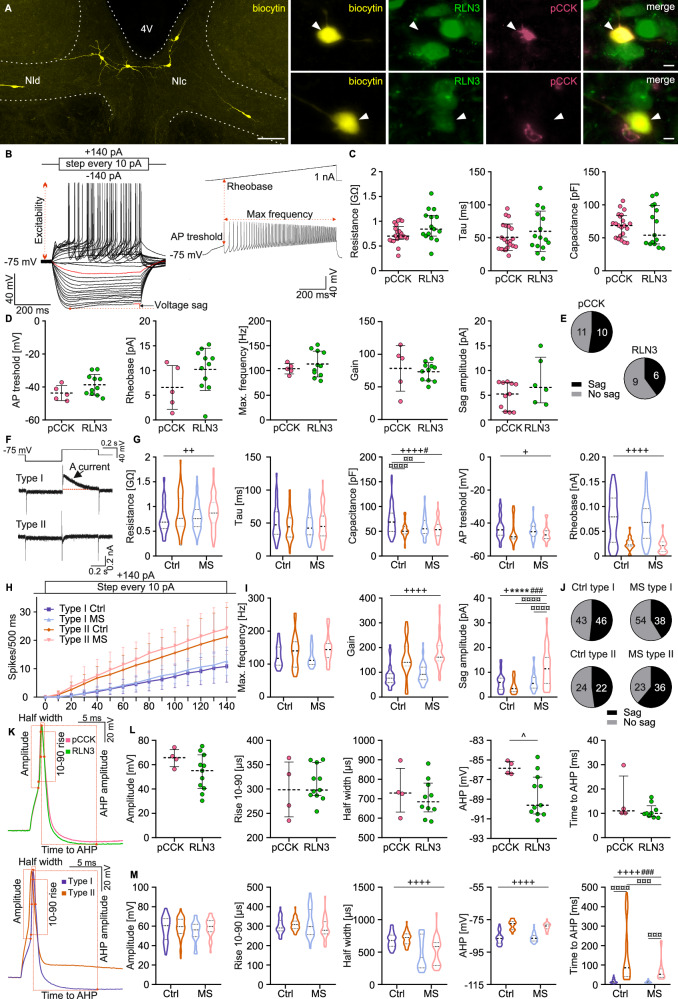 Figure 2
Figure 2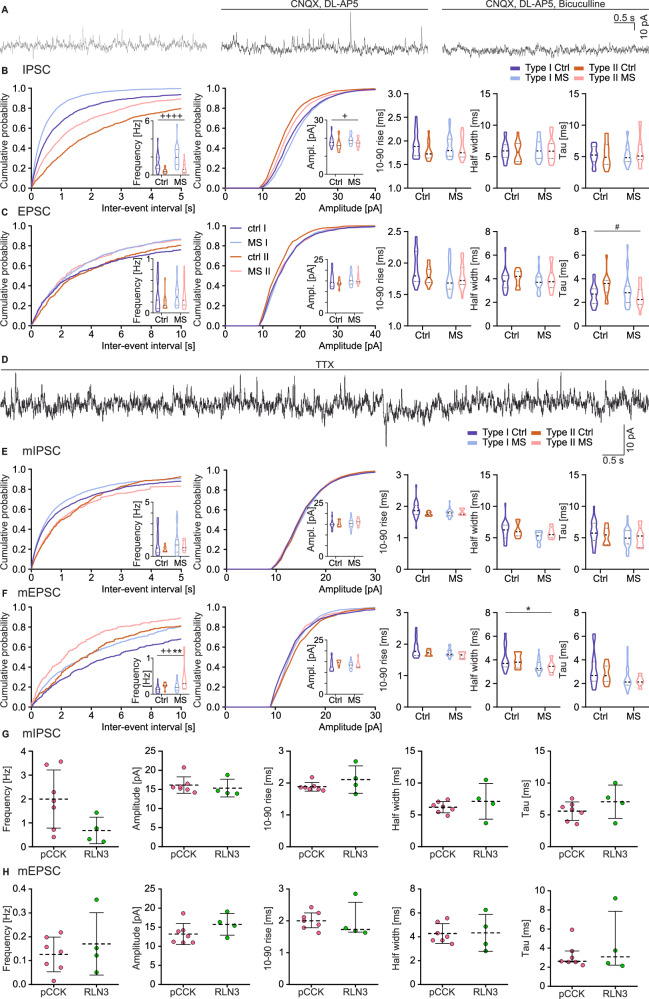 Figure 3
Figure 3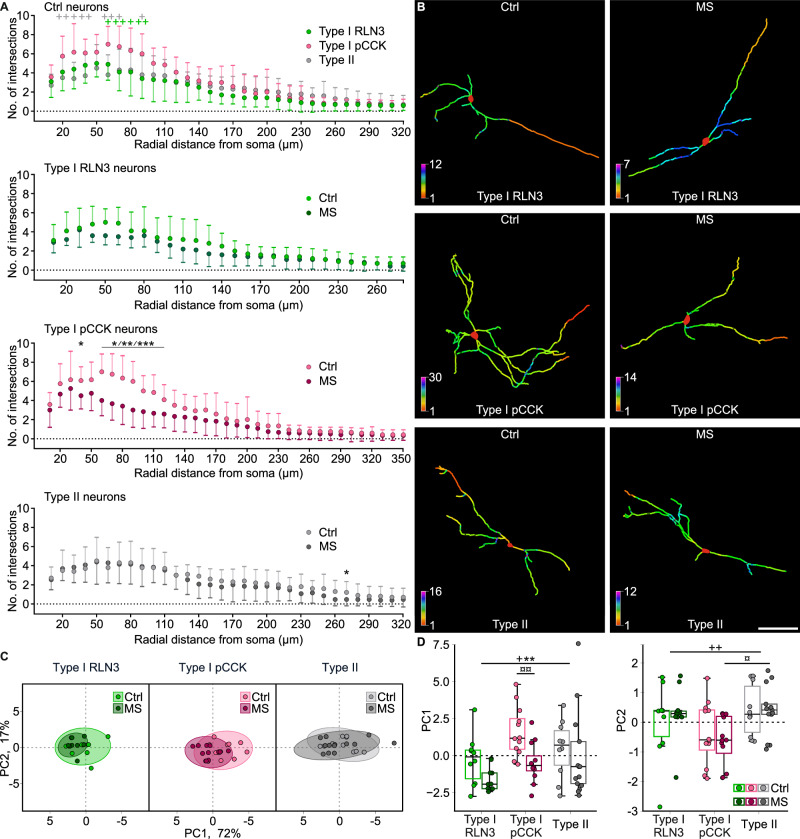 Figure 4
Figure 4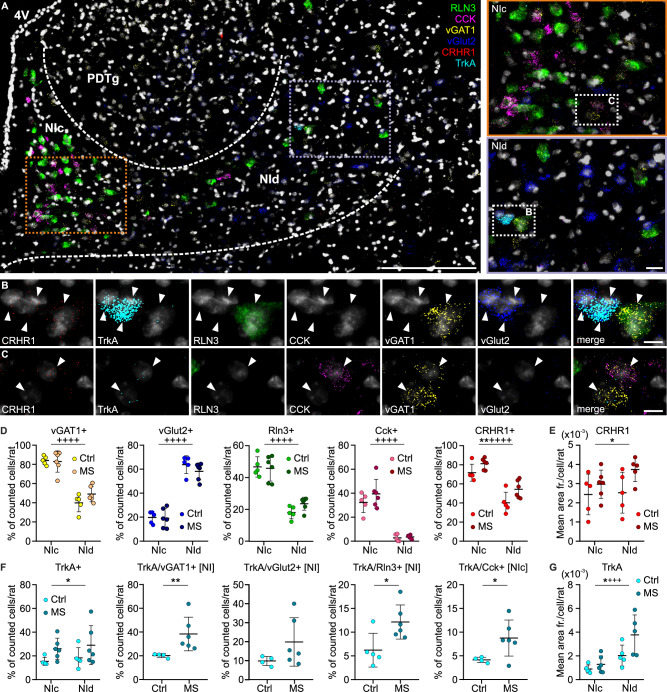 Figure 5
Figure 5- —https://doi.org/10.13039/501100004281Narodowe Centrum Nauki (National Science Centre)
- —European Union’s Horizon Europe research and innovation program under Marie Sklodowska-Curie grant agreement No 101086247 - PsyCoMed Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsStress Responses and Cortisol · Neuroendocrine regulation and behavior · Neuroscience of respiration and sleep
Introduction
Early-life stress (ELS) arising from physical and emotional abuse, disrupts normal brain development and impairs hypothalamic–pituitary–adrenal (HPA) axis function, and a large body of evidence has shown that children who experience such adversities have an elevated risk of developing psychopathological disorders in adulthood [1–3]. Moreover, a common consequence of experiencing ELS is increased susceptibility to compulsive behaviors, such as compulsive food consumption and addiction to psychoactive substances [4–6]. Furthermore, ELS increases stress sensitivity in adulthood and exposure to severe stressors throughout life can markedly increase the likelihood of developing both psychopathological disorders and substance abuse in individuals who have experienced ELS [7–12]. However, although the consequences of adverse experiences in early life associated with the development of mental disorders and substance abuse have been widely described, the underlying neural mechanisms are still largely unknown.
An area of the brain that is currently unexplored in the context of ELS sensitivity, is the brainstem nucleus incertus (NI), a highly stress-sensitive and conserved component of an ascending arousal network, described in human [13], macaque [14], rat [15], mouse [16] and fish [17] brain. The NI contains heterogeneous populations of mainly GABA, but also glutamate, neurons, that give rise to ascending forebrain projections, and it is the major source of the neuropeptide, relaxin-3 (RLN3) in the brain [15, 18–21]. Considerable pharmacological and functional data indicate an important role of the NI and RLN3 in stress responses and behavioral activation [15, 22–29], food intake control [30–32], compulsive behaviors, such as binge eating [33–35], and stress-induced reward seeking [36–38]. Human and animal studies have shown that NI neurons abundantly express stress-related corticotropin-releasing hormone receptor type 1 (CRHR1), and virtually all RLN3-positive NI neurons co-express CRHR1 [27, 39, 40]. RLN3 and its cognate receptor, RXFP3 (relaxin-family peptide 3 receptor), control stress-induced alcohol preference in mice [38] and reinstatement of alcohol seeking in rats [36, 37, 41, 42], via CRHR1 signaling in the NI [27, 36]. Importantly, the potent effects of ELS on the expression of CRH and CRHR1, as well as the tight link between this influence and increased susceptibility to developing substance abuse have been repeatedly demonstrated in humans and animals [43, 44], yet the possible influence of ELS on NI neurons, including CRHR1 expression, remains unknown. Notably, in addition to RLN3, NI neurons synthesize other neuropeptides, including cholecystokinin (CCK) [15, 45]. Our previous studies have demonstrated that CCK is synthesized by a separate neuronal population to that synthesizing RLN3, but the vulnerability of NI CCK neurons to different stressors, including ELS and acute stress remains to be elucidated.
The detrimental consequences of ELS also involve disturbances in nerve-growth factor (NGF)-mediated tropomyosin receptor kinase A (TrkA) signaling [46, 47]. NGF/TrkA signaling, in addition to its canonical trophic actions, is strongly implicated in stress responses, and exerts stimulatory actions on the HPA axis. Furthermore, NGF levels are sensitive to stressful events [48–50], and NGF/TrkA signaling is strongly implicated in alcohol and other drug use disorders [51, 52], but the underlying neuronal mechanisms are not well understood. Notably, NI neurons in the rat abundantly express TrkA [53, 54], yet to our knowledge, there is no data regarding the influence of stress on TrkA expression in the brainstem, including the NI.
Therefore, in the current study, we examined the influence of neonatal maternal separation (MS), a well-documented animal model of ELS, on the expression of specific neuropeptide and stress-related receptor mRNA species in GABA and glutamate NI neurons in male rats. We also examined the impact of MS on the synaptic activity, excitability, and membrane electrophysiological properties of defined NI neurons, and the consequences for NI neuronal morphology. Finally, we determined the influence of MS on the sensitivity of NI neurons to acute stress in adulthood.
Materials and methods
Animals and maternal separation procedure
All procedures were approved by the Second Local Institutional Animal Care and Use Committee (Krakow, Poland) and conducted in accordance with the EU Directive 2010/63/EU on the protection of animals used for scientific purposes. Male Sprague-Dawley rats used in all experimental procedures were obtained from the Institute of Zoology and Biomedical Research Animal Facility, Jagiellonian University, Krakow. Rats were housed under standard conditions (12:12 light/dark cycle, lights on 08:00–20:00, temperature 22–23 °C) with ad libitum access to food and water. The MS procedure was performed between postnatal days (PND) 2–14, during which pups were separated from their dams for 3 h daily, as previously described [55]. Control (Ctrl) rats remained undisturbed. Pups were weaned on PND 28 and housed in family groups until use in further experimental procedures.
Acute stress and c-Fos expression in neurochemically-defined nucleus incertus neurons
On PND 55–56, rats were subjected to 30 min restraint stress in transparent plastic bag restrainers (control stressed, Ctrl S, n = 5; MS stressed, MS S, n = 6). Non-stressed Ctrl rats remained in transport cages in the experimental room for 30 min (Ctrl NS, n = 6; MS NS, n = 5). One hour post-stress, rats were deeply anesthetized and transcardially perfused with phosphate-buffered saline (PBS) followed by 4% phosphate-buffered formaldehyde. Brains were collected, post-fixed overnight and cryoprotected in 30% sucrose solution in PBS, before sectioning into 45 µm coronal slices for immunohistochemistry. Sections were immunostained against c-Fos, RLN3, and pro-cholecystokinin (pCCK), using fluorescent secondary antibodies. Images were acquired with an Axio Imager M2 fluorescence microscope (Zeiss, Gottingen, Germany), and neurons immunopositive for RLN3 (RLN3+) and pro-CCK (pCCK+), co-expressing c-Fos protein (c-Fos+) or without such co-expression (c-Fos–) were quantified.
Electrophysiological patch clamp recordings
A total of 38 male rats (PND 35–56) were used in electrophysiological experiments (Ctrl, n = 17; MS, n = 21). Whole-cell recordings were performed as previously described [45]. A series of electrophysiological tests was conducted under voltage-clamp and current-clamp conditions. Subsequently, spontaneous postsynaptic currents (sPSCs) were recorded under voltage-clamp (holding potential: –50 mV). To examine miniature postsynaptic currents (mPSCs), a separate group of neurons was recorded in the presence of tetrodotoxin (TTX, 0.5 μM). Both sPSCs and mPSCs were detected offline and analyzed using Mini Analysis software (Synaptosoft Inc., Fort Lee, NJ, USA). After the recordings, slices were fixed in 4% formaldehyde and incubated with primary mouse and rabbit antibodies against RLN3 and pCCK, respectively. Then, slices were incubated with secondary antibodies: Alexa Fluor 647 donkey anti-rabbit, Alexa Fluor 488 donkey anti-mouse, and ExtrAvidin-Cy3, followed by mounting with Fluoroshield containing DAPI.
Dendritic tracing and morphological analysis
After electrophysiological recordings, slices containing biocytin-filled neurons were immunostained for RLN3 and pCCK and imaged using a Zeiss LSM 780 confocal microscope. Selected well-stained neurons (RLN3: Ctrl, n = 10, and MS, n = 10; pCCK: pCCK, Ctrl, n = 12, and MS, n = 12) were subjected to dendritic tracing, used for Sholl analysis [56] and acquisition of topological parameters of the dendritic tree.
Multiplex fluorescent in situ hybridization
Multiplex RNAscope HiPlex in situ hybridization (Advanced Cell Diagnostics (ACD), Hayward, CA, USA) was performed using fresh-frozen NI-containing brain sections (16 µm) from adult rats (PND 51–57; Ctrl, n = 5; MS, n = 7), following the manufacturer’s user manual as described [15]. Probes targeted RLN3, CCK, vGAT1, CRHR1, vGlut2, and TrkA mRNA. Matching ROIs located in NIc and NId were imaged using the fluorescence microscope, and mRNA-expressing NI cells were counted. CRHR1 and TrkA mRNA expression levels (area fraction) per cell were quantified.
Statistical analysis
Data were analyzed using Prism 8 (GraphPad) and R software. Normality was tested with the Shapiro–Wilk test, and variance homogeneity with Levene’s test. Parametric data were analyzed using unpaired t-tests or ANOVA with a post-hoc Tukey’s test; non-parametric data were analyzed with a Mann–Whitney test or type III two-way ANOVA with White’s correction. Outliers were removed using ROUT (Q = 1%). Data analysis was performed by experimenters blinded to the treatment conditions.
More details of used procedures are included in the Supplementary Materials.
Results
MS and restraint stress differentially affected c-Fos expression in NI neurons
In studies to determine the impact of MS on the sensitivity of NI neurons to acute stress in adulthood, rats (8 weeks old) from control (Ctrl) and MS cohorts were divided into groups that were subjected to restraint stress (S) or not (NS). Activation of relaxin-3 (RLN3+) and pro-cholecystokinin (pCCK+) positive NI neurons was assessed in these groups by the presence of c-Fos protein. RLN3+ and pCCK+ neurons with (+) and without (–) c-Fos were counted (RLN3+/c-Fos+, pCCK+/c-Fos+ and RLN3+/c-Fos–, pCCK+/c-Fos–) (Fig. 1A–C). A semi-automated analysis of binarized signal from NI-located c-Fos+ nuclei was also performed.Fig. 1. Maternal separation and restraint stress differentially modify the number of c-Fos-expressing NI neurons.A Representative images of c-Fos protein expression in NI. Scale bar: 200 μm. Magnified NIc area with RLN3- (B) and pCCK-immunopositive (C) NI neurons expressing c-Fos in all groups of rats. Scale bar: 20 μm. D The number of all NIc c-Fos+ cells was increased by restraint stress and decreased by MS under both conditions (NS and S). E The proportions of c-Fos+ neurons within RLN3+ NI cells (left) and pCCK+ NIc cells (right) were increased by restraint stress in both NI neuronal populations but affected (decreased) by MS only in the RLN3+ population. F, G MS and restraint stress differentially influenced the number of neurochemically defined c-Fos+ and c-Fos– NI/NIc neurons but did not significantly affect total numbers of RLN3+ or pCCK+ cells in these areas. Statistical significance in (D–G) was determined using two-way ANOVA with a post-hoc Tukey test: *MS effect, $ restraint stress effect, # interaction of MS and restraint stress, ¤ post-hoc. The number of each symbol indicates the level of statistical significance: *(p < 0.05), **(p < 0.01), ***(p < 0.001). See Supplementary Table 1 for details. 4V 4th ventricle, NI nucleus incertus, NIc nucleus incertus pars compacta, NId nucleus incertus pars dissipata, MS maternal separation, pCCK pro-cholecystokinin, RLN3 relaxin-3.
Both MS and restraint stress altered the total number of c-Fos-expressing cells in the NI pars compacta (NIc). Expression of c-Fos in NIc was increased by restraint and decreased by MS in both groups (Ctrl S and MS S; Fig. 1D, Supplementary Table 1). An effect of restraint stress was also observed in the NI pars dissipata (NId), and when the whole NI area was analyzed (Supplementary Fig. 1a).
Cell counting and subsequent ANOVA revealed that the percentage of c-Fos+ cells within the total RLN3+ NI neuronal population was increased by restraint stress in both the Ctrl and MS groups, and MS decreased their responsiveness regardless of the acute stress condition (Fig. 1E, Supplementary Table 1). A similar effect was observed on the number of RLN3+/c-Fos+ cells (Fig. 1F). Concurrently, MS or restraint stress did not affect the total number of RLN3+ neurons in the NI (Fig. 1F, Supplementary Figure 1c, Supplementary Table 1). The effects of restraint stress were significant in NIc but not NId, with respect to the percentage of RLN3+/c-Fos+ cells, and only in NIc, with respect to their number (Supplementary Fig. 1b, d, Supplementary Table 1).
Counting of pCCK+ neurons was performed only in NIc, as CCK in the NI is synthesized almost exclusively by neurons located in this area [15]. Similar to the effect on RLN3+ cells, ANOVA indicated that restraint stress induced an increase in the percentage of c-Fos+ neurons within the total pCCK+ NIc population, with no effect of MS on their responsiveness (Fig. 1E, Supplementary Table 1). These results were reflected in the number of pCCK+/c-Fos+ neurons (Fig. 1G, Supplementary Table 1). Notably, an interaction of restraint stress and MS was detected in the total number of pCCK+ NIc cells (Fig. 1G, Supplementary Table 1), indicating that MS altered the baseline number of pCCK+ neurons and modified the influence of acute stress on pCCK synthesis. The altered number of pCCK+ neurons under the examined conditions, along with the lack of influence of MS on the number of activated pCCK+ neurons (pCCK+/c-Fos+), was reflected in the ANOVA results, which revealed a significant interaction of restraint stress with MS in relation to the number of non-activated pCCK+ neurons (pCCK+/c-Fos–). Post-hoc analysis revealed that MS produced a significant increase in the number of pCCK+/c-Fos– neurons, and restraint stress in the MS group reversed this effect (Fig. 1G, Supplementary Table 1).
Notably, restraint stress altered the proportions of c-Fos+ and c-Fos– pCCK+ neurons, towards a higher level of c-Fos expression. The effect was significant in both the Ctrl and MS groups, but more pronounced in the latter (Supplementary Fig. 1f, Supplementary Table 2).
MS altered the electrophysiology of adult NI neurons
MS altered the active and passive membrane properties of NI neurons
In studies to investigate the impact of MS on the passive and active membrane properties of NI neurons, a series of ex vivo whole-cell patch-clamp experiments was conducted in current clamp mode (holding potential –75 mV). Immunohistochemical procedures combined with anti-biocytin staining, that followed these electrophysiological experiments, allowed the categorization of examined neurons based on their spatial distribution (NIc and NId) and peptide content (RLN3+ NI neurons and pCCK+ NI neurons) (Fig. 2A).Fig. 2. Influence of electrophysiological type and MS on passive and active neural properties of NI neurons.A Immunohistochemical localization of recorded neurons in the NI, filled with biocytin (left side, scale bar: 100 μm), and fluorescent projection images of biocytin-filled pCCK+ (top) or RLN3+ (bottom) NI neurons, indicated by arrowheads (scale bar: 20 μm). B Current step stimulation protocol (top left) and voltage responses of an exemplary NI neuron (bottom left), illustrating the measured neural properties. Neuronal excitability (input-output relationship) was assessed by counting the number of APs evoked by depolarizing current pulses (0–140 pA). Membrane resistance, time constant, and capacitance were measured from the voltage response to a –20 pA hyperpolarizing current pulse. Resistance was calculated by dividing the voltage by the current, according to the ohmic current-voltage relationship; the time constant was measured in the charging phase of the voltage response, and capacitance was calculated by dividing the resistance and time constant. Voltage sag was measured from the voltage response to a –140 pA hyperpolarizing current step. Current ramp stimulation protocol (top right) and voltage response of the exemplary NI neuron (bottom right) illustrating the measured properties. The AP threshold and rheobase were measured when the first AP occurred; the maximal spiking frequency was calculated using the minimal inter-spike interval. C, D Strip charts comparing membrane properties of pCCK+ (pink) and RLN3+ (green) NI neurons. E Pie charts illustrating the proportions of pCCK+ and RLN3+ neurons that were characterized by the occurrence and lack of sag. F Voltage clamp protocol used to test the presence of the potassium A-type current (top). Current response of a type I NI neuron showing visible A-type current (middle) and a type II NI neuron lacking A-type current (bottom). G Violin charts illustrating ANOVA comparison of active and passive neural properties of type I and type II NI neurons under control and MS conditions. H Current step stimulation protocol (top) and number of APs evoked in response to the stimulation (bottom). The slope of the regression was considered as the gain. I Violin charts illustrating ANOVA comparison of active neural properties of type I and type II NI neurons in control and MS conditions. J Pie charts illustrating the proportions of neurons that were characterized by the occurrence and lack of sag in control and MS rats. K Exemplary waveform of AP with comparison between pCCK and RLN3 neurons (top), and type I and type II NI neurons (bottom), illustrating the measured properties. L Strip charts representing the comparison between membrane properties of pCCK+ (pink) and RLN3+ (green) NI neurons. M Violin charts representing ANOVA comparisons between type I and type II NI neurons under control and MS conditions. The following symbols were used for statistical significance determined using two-way ANOVA with a post-hoc Tukey test: *MS effect, +neuronal type effect, # interaction, ¤ post-hoc; or ^t-tests. The number of each symbol indicates the level of statistical significance: *(p < 0.05), **(p < 0.01), ***(p < 0.001), ****(p < 0.0001). See Supplementary Tables 3, 5 and 6 for details.
NI neurons can be divided into electrophysiological types based on presence (type I) or absence (type II) of an A-type potassium current (Fig. 2F) [19]. Our previous studies have shown that both RLN3 and CCK cells are predominantly type I NI neurons [15, 45]. In the current study, a series of electrophysiological tests allowed for further characterization of RLN3 and CCK neurons (Fig. 2B–E, Supplementary Table 3). Results revealed no significant differences in the active or passive membrane properties of pCCK+ and RLN3+ cells, suggesting homogeneity of the electrophysiological properties of type I NI neurons.
ANOVA and further analysis of the results obtained revealed that in both control and MS groups, there were significant differences between type I and type II NI neurons in the majority of tested membrane properties (i.e., resistance, capacitance, action potential (AP) threshold, rheobase and neuronal gain). Importantly, MS significantly decreased the membrane capacitance in type I neurons and caused an increase in sag amplitude in type II NI neurons (Fig. 2H–J, Supplementary Fig. 2e, Supplementary Tables 5 and 6). Additional analysis was conducted to verify whether MS affected NI neurons differently depending on their electrophysiological type (I/II), localization (NIc/NId) and neurochemical nature (i.e., presence or absence of RLN3 or pCCK). Type I NI neurons localized in NIc were particularly sensitive to MS, as MS increased resistance, decreased capacitance and increased neuronal gain in these neurons (Supplementary Fig. 2b–d) The results also revealed a significant effect of MS on the sag amplitude, AP maximum frequency and neuronal gain of NIc RLN3 neurons, and on the capacitance of pCCK neurons (Supplementary Fig. 2). MS also influenced the time constant (tau) of type II neurons in NId (Supplementary Fig. 2b). Details of the statistical analysis are reported in Supplementary Table 7.
MS altered the action potential properties of NI neurons
In studies to assess the AP properties of NI neurons and the possible influence of MS on AP shape, the membrane potential of recorded cells was held at –75 mV, and a single depolarizing current pulse exceeding the excitability threshold was applied (0.75 nA, 2 ms). Analysis of AP properties revealed similarities between pCCK+ and RLN3+ neurons in all but one tested feature, AHP amplitude, which was higher in RLN3 cells (Fig. 2K, L, Supplementary Table 3).
For the ANOVA of AHP-related properties, only the slow AHP (sAHP, occurring 25–572 ms after AP peak), present in both type I and type II NI neurons, was considered. Fast AHP (fAHP, occurring 1–9 ms after AP peak), detected only in a fraction of type II neurons, was not included in this analysis. ANOVA revealed that type I and II NI neurons differ in terms of AP half-width, amplitude, and time to the AHP, regardless of the procedure (Ctrl/MS) to which the tested rats were subjected (Fig. 2K, M, Supplementary Table 5). Notably, MS decreased the time to AHP in an electrophysiological type-dependent manner, particularly impacting type II neurons (Fig. 2M). In-depth analysis designed to indicate the possible influence of MS on AP properties of NI neurons differing in terms of electrophysiological type, neuron localization and presence/absence of the different neuropeptides revealed specific shortening of AP (shorter AP half-width) in NIc type II neurons (Supplementary Fig. 2f). Within this analysis, the fast AHP and slow AHP of type II NI neurons were considered, and notably MS caused fast AHP to occur more rapidly in type II NI neurons localized in NIc but did not affect the slow AHP (Supplementary Fig. 2g). Details of the statistical analysis are reported in Supplementary Table 7.
MS modified synaptic activity within the NI
Synaptic activity within the NI was assessed by analyzing the 200 s long recordings in the voltage clamp configuration (holding potential –50 mV). Under these conditions, both outward and inward postsynaptic currents (PSCs) were observed, and their dependence on GABAergic and glutamatergic signaling was confirmed using respective receptor antagonists (Fig. 3A), indicating that these are inhibitory PSC (IPSCs) and excitatory PSCs (EPSCs), respectively. The spontaneous postsynaptic currents (sPSCs) were recorded in standard ACSF, and frequency, amplitude, 10–90 rise, half-width and tau were evaluated for recorded inward and outward PSCs. ANOVA revealed that type I NI neurons were characterized by higher frequency, as well as higher amplitude sIPSCs than type II neurons in both examined groups (Fig. 3B, Supplementary Table 8). Notably, MS impacted NI neurons in a type-dependent manner, as it produced an increase in the tau value in type I and decreased this value in type II NI neuron EPSCs (Fig. 3C, Supplementary Table 8). Further analysis revealed that MS caused a decrease in rise time in type I NIc neurons. In the case of type II neurons, MS caused a decrease in the EPSC tau value only in NIc (Supplementary Figure 3d, Supplementary Table 9).Fig. 3. Spontaneous and miniature synaptic inputs to NI neurons.A Exemplary trace of NI spontaneous synaptic events recorded in voltage clamp at –50 mV holding potential (left), along with traces illustrating the abolition of inward currents with glutamatergic receptors antagonists (middle) and abolition of both inward and outward PSC with glutamatergic and GABAergic receptors antagonists (right). B Cumulative distribution histograms of the sIPSC (bin: 10 ms), strip chart illustrating the comparison between type I and II NI neurons in the control group and after MS (right-hand side), and strip charts representing comparison between type I and type II NI neurons in control and MS groups. C Cumulative distribution histograms of the sEPSC (bin: 10 ms), strip chart illustrating the comparison between type I and II NI neurons in the control group and after MS (right-hand side) and strip charts representing a comparison between type I and type II NI neurons in control and MS groups. D Exemplary trace of NI miniature synaptic events recorded in voltage clamp at –50 mV holding potential, with TTX added to the ACSF. E Cumulative distribution histograms of the mIPSC (bin: 10 ms), strip chart illustrating the comparison between type I and II NI neurons in the control group and after MS (right-hand side) and strip charts representing comparison between type I and type II NI neurons in the control and MS groups. F Cumulative distribution histograms of the mEPSC (bin: 10 ms), strip chart illustrating the comparison between type I and II NI neurons in the control group and after MS (right-hand side) and strip charts representing a comparison between type I and type II NI neurons in control and MS groups. G Strip charts representing a comparison between mIPSC properties of the pCCK+ (pink) and RLN3+ (green) NI neurons. H Strip charts representing comparison between mEPSC properties of the pCCK+ (pink) and RLN3+ (green) NI neurons. The following symbols were used for statistical significance determined using two-way ANOVA with a post-hoc Tukey test: *MS effect, +neuronal type effect, #interaction. The number of each symbol indicates the level of statistical significance: *(p < 0.05), **(p < 0.01), ****(p < 0.0001). Please see Supplementary Tables 4 and 8 for details.
Miniature PSCs (mPSCs) were recorded from NI neurons in the presence of TTX in ACSF (Fig. 3D). ANOVA revealed a significantly higher frequency of mEPSC in type II than in type I NI neurons in both Ctrl and MS groups (Fig. 3F, Supplementary Table 8). In contrast, no differences were observed for mIPSC (Fig. 3E, Supplementary Table 8). Importantly, MS caused an increase in the frequency and a decrease in the half-width of the mEPSCs in both types of studied neurons (Fig. 3F, Supplementary Table 8). Additional analysis revealed that pCCK+ and RLN3 + NI neurons did not differ in the properties of their synaptic inputs in the Ctrl group (Fig. 3G, H, Supplementary Table 4) and that MS increased the rise time of mIPSCs while decreasing the half-width and tau values in type I NIc neurons, regardless of their neuropeptide content. Moreover, MS increased the frequency of mEPSCs while decreasing their rise time, half-width, and tau values in type I NIc neurons, independent of neuropeptide content (Supplementary Fig. 3e–h, Supplementary Table 9). In positively-stained cells, MS affected synaptic activity mainly in pCCK+ neurons, as it led to a decrease in the rise time and half-width of mIPSCs, as well as a decrease in the rise time, half-width, and tau of the mEPSCs recorded from NIc pCCK+ neurons (Supplementary Fig 3g, h). MS also produced an increase in the frequency of mIPSCs in NIc RLN3+ neurons, indicating that MS influences both inhibitory and excitatory synaptic transmission in pCCK+ and only inhibitory synaptic transmission in RLN3+ neurons (Supplementary Fig. 3e). Details of the statistical analysis are provided in Supplementary Table 9.
Overall, our electrophysiological recording revealed that MS significantly altered the neurophysiological properties of NI neurons, in an electrophysiological type-dependent manner, affecting capacitance, resistance, and synaptic activity. Type I neurons exhibited reduced capacitance and increased neuronal gain, while type II neurons displayed changes in AP dynamics, including increased sag amplitude and faster AHP. MS also influenced synaptic transmission, altering excitatory and inhibitory activity in specific neuron subtypes. The effect of MS also related to the neuron peptide content and location within the NI (as detailed in the Supplementary Materials).
MS produced type-dependent dendritic shrinkage of NI neurons
Following the electrophysiological experiments, dendritic tracing of biocytin-filled type I RLN3+ , type I pCCK+ and type II NI neurons was performed to investigate the possible influence of MS on the morphology of these cells. On the basis of the resultant data, Sholl and additional analyses of dendritic tree topological parameters were performed.
Two-way ANOVA and post-hoc tests revealed that in control rats type I pCCK+ neurons had the most complex dendritic tree among the tested groups, with more intersections in its proximal part (Fig. 4A, B, Supplementary Fig. 4, Supplementary Tables 10, 11). Interestingly, type I pCCK+ neurons were the most sensitive to MS, and MS caused significant shrinkage of their dendritic tree in its proximal and middle parts (Fig. 4A, B, Supplementary Table 11). Additional analysis revealed that this effect was also reflected in the decreased maximal number of all intersections and their sum: MS lowered these parameters, exclusively in pCCK+ neurons (Supplementary Fig. 4, Supplementary Table 10). Concurrently, MS had no marked effect on type I RLN3+ and type II cells (Fig. 4A, B, Supplementary Fig. 4, Supplementary Tables 10, 11).Fig. 4. Maternal separation caused a selective dendritic shrinkage of pCCK-immunopositive NI neurons.A Sholl analysis of 3D-reconstructed control and MS NI cells, illustrating differences between the complexity of studied types of control neurons, with significantly more complex type I pCCK+ cells (top panel), exclusively affected by MS (lower middle panel). The x-axis range varies with the radial range of dendrites within a given cell type. B Representative skeletal 3D reconstructions of defined NI neurons from control and MS groups. Colors indicate the number of dendritic intersections with virtual Sholl spheres. Note a pronounced MS-induced dendritic atrophy of type I pCCK+ neurons. Scale bar: 100 μm. C, D Principal component analysis of measured topological parameters of the dendritic tree (see Supplementary Tables 12–14 for details). C Scatter plots of the first two principal components – a comparison of control and MS neurons of all studied types. Loadings of parameters used in the PCA are listed in Supplementary Table 15. D Statistical analysis of PC1 and PC2, indicating the most complex dendritic tree in the case of type I pCCK+ neurons and its MS-induced selective complexity decrease. Statistical significance in (A) and (D) was determined using two-way ANOVA with a post-hoc Tukey test: *MS effect, +neuronal type effect, ¤post-hoc. The number of each symbol indicates the level of statistical significance: *(p < 0.05), **(p < 0.01), ***(p < 0.001). The “+” symbol colors in (A) represent the difference between type I pCCK+ cells and type I RLN3+ (green) or type II (gray) cells. Please see Supplementary Tables 11 and 16 for details.
Topological parameters describing the shape of the dendritic tree, i.e., the number of (1) primary dendrites, (2) bifurcations, (3) branches and (4) dendritic tips, and (5) the total dendritic length, and (6) the maximal branch order, were analyzed using principal component analysis (PCA), in order to reduce the number of highly correlated features (Supplementary Tables 12, 13). The first two principal components (PCs), explaining 89% of the observed variance (Fig. 4C), were considered for further analysis. Parameters (2–5) were highly and positively correlated with PC1, whereas parameter (1) was negatively correlated with PC2 (Supplementary Table 14; see Supplementary Table 15 for loadings). Two-way ANOVA applied to PC1 revealed an effect of both MS and neuronal type, and a post-hoc test revealed a significant MS-induced decrease in complexity that was selective for type I pCCK+ neurons (Fig. 4D, Supplementary Tables 12, 16). In the case of PC2, a significant effect of neuronal type was observed, but post-hoc comparisons revealed significant differences only between type I pCCK+ and type II neuron dendritic trees in the MS group, with the former more complex than the latter (Fig. 4D, Supplementary Tables 12, 16).
MS altered the expression of stress-related receptor mRNA in the NI
In order to verify the influence of MS on the expression of mRNA encoding key neuropeptides (RLN3, CCK), classical markers for inhibitory and excitatory neurons (vGAT1, vGlut2), and stress-related receptors (CRHR1, TrkA), multiplex fluorescence in situ hybridization was conducted using coronal sections throughout the rostrocaudal NI. Cells from two ROIs; NIc and NId, were included in the analysis (Fig. 5A–C). No differences between the total number of counted control and MS neurons were observed (Supplementary Table 17).Fig. 5. Expression of different mRNA species in control and MS rats.A Representative image of NI neurons expressing RLN3 (green), CCK (magenta), vGAT1 (yellow), vGlut2 (blue), CRHR1 (red) and TrkA (cyan) mRNAs. Scale bar: 200 μm. Enlarged sections of NIc (orange rectangle) and NId (purple rectangle) selected for cell counting are displayed on the right. Scale bar: 20 μm. B, C A series of images illustrating exemplary NI neurons (white arrowheads) expressing each mRNA examined with DAPI-stained nuclei (light-gray). Scale bars: 10 μm. D The percentage of counted cells expressing the examined mRNAs per rat in both groups. E The mean area fraction of CRHR1-immunofluorescent dots per cell. F The percentage of counted cells expressing TrkA mRNA in control and MS rats, and TrkA in combination with other studied mRNA species. G The mean area fraction of TrkA-immunofluorescent dots calculated per single NI cell. Statistical significance was determined by two-way ANOVA: *MS effect, +localization effect; *t-test (with Welch correction if needed) or Mann–Whitney test. The number of each symbol indicates the level of statistical significance: *(p < 0.05), **(p < 0.01), ***(p < 0.001), ****(p < 0.0001). Please see Supplementary Tables 17–22 for details. 4V 4th ventricle, NIc nucleus incertus pars compacta, NId nucleus incertus pars dissipata, PDTg posterodorsal tegmental nucleus, RLN3 relaxin-3, CCK cholecystokinin, CRHR1 corticotropin-releasing hormone receptor type 1, TrkA tropomyosin receptor kinase A, vGAT1 vesicular γ-aminobutyric acid (GABA) transporter, vGlut2 vesicular glutamate transporter.
Counting of RLN3-mRNA-positive (Rln3+) and CCK-mRNA-positive (Cck+) cells, followed by two-way ANOVA revealed a significant NIc/NId localization effect, without an MS effect (Fig. 5D, Supplementary Table 18). A similar effect was observed for NI neurons expressing vGAT1 mRNA (vGAT1+) and vGlut2 mRNA (vGlut2+) (Fig. 5D, Supplementary Table 18). Almost all Rln3+ and Cck+ cells were vGAT1-mRNA-positive, and if any expressed vGlut2 mRNA, there was also co-expression of vGAT1 mRNA, as previously reported [15, 45].
CRHR1 mRNA (CRHR1+) was observed to be co-expressed with vGAT1+, vGlut2+, Rln3+ and Cck+ mRNA (Fig. 5B, C and Supplementary Table 19). Importantly, a significant MS effect was observed for the NI cells expressing CRHR1 mRNA, indicating an increased number of CRHR1+ neurons, regardless of the NI area (Supplementary Table 19). When individual subtypes of CRHR1+ cells were analyzed, only in the case of CRHR1+/vGAT1+ NId neurons was a significant MS effect detected (Fig. 5D, Supplementary Table 19). Moreover, in rats subjected to MS, an increase in the level of CRHR1 mRNA within individual NI neurons was observed, as the mean area fraction of CRHR1 mRNA-related immunofluorescent dots per cell was higher in MS rats (Fig. 5E, Supplementary Table 20).
TrkA mRNA was found to be co-expressed with vGAT1+, vGlut2+, Rln3+ and Cck+ mRNA (Fig. 5B, C and Supplementary Table 21). Importantly, MS caused an increase in the number of NI cells expressing TrkA mRNA (TrkA+, Fig. 5F, Supplementary Table 18), specifically in vGAT1+, Rln3+ and Cck+ neurons, but not in TrkA+/vGlut2+ neurons (Supplementary Table 21). The increase in the number of TrkA+/vGAT1+ and TrkA+/Rln3+ neurons was observed in the whole NI area, while the number of TrkA+/Cck+ cells increased specifically in NIc (the main area of CCK synthesis; Fig. 5F, Supplementary Table 21). Notably, the mean area fraction of TrkA mRNA-related immunofluorescent dots calculated per individual NI neuron was also increased in MS rats and generally higher in NId (Fig. 5G, Supplementary Table 20).
Discussion
In the current study we demonstrated that the NI, an important element of the central stress response system implicated in stress induced food intake and alcohol seeking, is highly sensitive to ELS. Specifically, we observed a decrease in the basal activity of NI RLN3 neurons in adult male rats exposed to ELS produced by MS between PND2–14. Additionally, we observed that ELS leads to an increase in the number of pCCK-positive neurons within the NI, coinciding with a change in their acute stress responsiveness in adulthood. Concurrently, we observed that ELS produced a severe impairment in pCCK neuron dendritic tree structure. We also demonstrated that ELS modulated the electrophysiological membrane properties, and synaptic transmission of NI neurons, in a cell-type-specific manner. Our data also revealed that ELS increased levels of mRNA encoding CRHR1 and TrkA mRNAs in the NI of adult male rats.
ELS and stress-responsiveness of NI neurons
NI consists mainly of GABAergic projection neurons that innervate a wide range of forebrain structures [57, 58], and has been implicated in the control of contextual memory, arousal, and responses to stress [22, 23, 27, 59]. Here we demonstrated that NI neurons are sensitive to ELS, and that early-life disturbances may alter the functioning of NI RLN3 and CCK neurons. Notably, we revealed that both these neuronal populations reacted to acute restraint stress with elevated activation, but ELS affected them differently. In the case of RLN3 neurons, ELS did not affect their number but significantly decreased the number of activated RLN3 cells in the adult brain, both under control conditions and after acute stress. These findings indicate a potential dysfunction of RLN3 neurons in rats that experienced MS and suggest their dampened stress responsiveness.
Notably, MS had a distinct effect on pCCK neurons, significantly increasing their overall number. However, despite this increase, acute stress activated a comparable number of pCCK neurons in both MS-exposed and control rats. This effect suggests that while MS alters the baseline number of detectable pCCK neurons, it does not influence their responsiveness to acute stress. The MS-induced increase in the pCCK neuron count may indicate a disrupted influence of acute stress on the pCCK-to-CCK transition in adult rats that experienced ELS. In neurons, CCK synthesis is regulated by multiple factors, with prohormone convertase 2 (PC2) being the primary enzyme responsible for this conversion [60]. Previous studies have demonstrated that PC2 expression is sensitive to hormonal fluctuations and various stressors [61–63], therefore dysregulation of PC2 may contribute to the observed alterations in pCCK neuron numbers in the present study. Consequently, the MS-induced increase in detected pCCK neurons may not correspond to a greater number of neurons producing the mature form of CCK, potentially reflecting a compensatory mechanism rather than a functional enhancement of CCK neurotransmission.
As the current study is the first to directly link pCCK NI neurons with neurogenic stress, further investigations are needed to elucidate the underlying mechanisms and the precise functional and behavioral implications. Unlike pCCK NI neurons, RLN3 NI neurons have been proven to be reactive to various stressors in rats, including acute restraint, as indicated by elevated c-Fos expression [39, 64]. Furthermore, both c-Fos expression and electrical activity were reported to be elevated in RLN3 NI neurons after intracerebroventricular (ICV) CRH injection [27, 39], and the excitatory action of CRH in the NI was blocked by a CRHR1 antagonist under ex vivo conditions [27], indicating a direct role of CRHR1 in CRH action in the rat NI. Our current results are consistent with these reports and confirm activation of NI neurons by acute stress.
It is notable that, although still limited, the available literature indicates sex differences in the sensitivity of the NI and the RLN3/RXFP3 system to stress. For example, central administration of RLN3 has been shown to induce sex-specific differences in the activation of the HPA axis, including greater activation of CRH neurons in the paraventricular hypothalamic nucleus (PVN) in male rats than in female rats [65, 66]. Additionally, increased RLN3 expression in the NI of female rats has been associated with reduced HPA axis activation in response to stress [30, 31]. These findings formed the basis for selecting males for the current study, but it has been shown that ELS may differentially affect the nervous system in males and females in both animal models and in humans [67]. Therefore, future studies should investigate the influence of ELS on the NI in females to better understand the mechanisms underlying sex differences in stress regulation and their potential therapeutic implications. Also, future studies should further explore the NI-originating input to the HPA axis, including the innervation of the PVN and RLN3/CCK receptor expression by CRH-producing neurons, to better understand the role of the NI in stress regulation.
ELS-related functional and structural alterations in the NI: possible mechanisms and consequences
The RLN3 and CCK NI neurons are mostly separate populations but are both type I NI cells [15], and our current data indicate that the passive and active membrane properties of RLN3 and CCK NI neurons are very similar. Importantly, we observed that ELS influenced the physiology of NI neurons in a specific fashion. While most of the passive and active membrane properties of NI neurons remained unchanged in adult rats exposed to ELS, the amplitude of the voltage sag, a characteristic of activating the hyperpolarization-activated cyclic nucleotide-gated (HCN) channel current (Ih), was significantly altered in an electrophysiological- and neurochemical-dependent manner. We observed that MS significantly reduced the sag amplitude in type I RLN3 NI neurons but increased it in type II neurons. Activation of HCN channels can exert dual effects, either excitatory or inhibitory, depending on the cell type [68, 69]. It can lead to membrane depolarization and an increase in neuronal excitability or, by increasing membrane conductance, to attenuation of membrane potential alternations, reduction of sensitivity to incoming stimuli and a decrease in postsynaptic currents summation [70]. In the current study, we observed that the sag amplitude in RLN3 NI neurons was significantly decreased by ELS experience, which was accompanied by an increase in neuronal gain in these cells. This indicates a possible ELS-induced decrease in the expression of HCN channels (or a lower level of their activation) in RLN3 neurons, with a consequent enhancement of the synaptic input summation capacity, increased excitability, and possible increased reactivity to stress-related stimuli. This mechanism may be associated with the involvement of the RLN3/RXFP3 system in stress-induced compulsive behaviors, such as binge alcohol and food intake [33, 34, 38]. Concurrently, ELS caused a significant increase in the sag amplitude of NIc type II neurons, that was also accompanied by an increase in the neuronal gain. These data, together with the ELS-induced increase in maximal firing frequency and shortened duration of the action potential, indicate a possible higher responsiveness of NIc type II neurons to incoming salient stimuli, in adult rats exposed to maltreatment during development. Notably, our previous data indicated that type I and II NI neurons belong to different neuronal circuits [15, 20], and here we demonstrated the ELS sensitivity of both neuronal types, which strongly implies the potential adverse impact of maltreatment during early life stages on distinct neuronal systems, that are under NI control.
The alterations in the electrophysiological properties of NI neurons induced by ELS, were accompanied by selective and pronounced dendritic shrinkage of type I CCK NI neurons. It has been previously demonstrated that ELS can dramatically alter neuronal morphology, and rodents experiencing ELS exhibit decreased dendritic arborization in the prefrontal cortex and hippocampus [71, 72], accompanied by modification of synaptic signaling [71, 73]. In accordance with these data, the ELS-induced shrinkage of CCK neuron dendritic trees was accompanied by significantly altered kinetics of both excitatory and inhibitory synaptic currents, indicating a postsynaptic origin of the observed changes. These findings reflect important neuroanatomical and functional consequences of ELS on the NI, which may underlie behavioral impairments related to NI-controlled functions, including reduced spatial memory, enhanced threat learning, and the increased vulnerability to substance abuse observed in humans and animals following this kind of stress [6, 74, 75].
Possible role of NI CCK neurons in stress responses
Very little is known about the precise function of NI CCK neurons, which we have shown are highly sensitive to ELS. Notably, an early study of CCK neurons that were described as located in the “caudal dorsal raphe nucleus” were shown to influence behavioral components of stress-adaptation responses and to impact the HPA axis response to chronic stress [76]. However, closer examination revealed that these neurons were in fact NI CCK neurons, and in our recent report we demonstrated that, together with RLN3 neurons, NI CCK cells form the NI–MS pathway [15], which implies their possible involvement in theta rhythm, spatial navigation, arousal, and motivation-related behavior control [23, 77–80].
Importantly, existing research has demonstrated a substantial involvement of CCK neurons in the regulation of substance abuse behavior, including alcohol abuse, through their heightened sensitivity to various stress factors, including ELS [81–83]. Similarly, several studies have demonstrated that the NI and the RLN3/RXFP3 system contribute to alcohol preference in rodents [27, 84]. Pretreatment with CRHR1 antagonists reduced alcohol seeking in mice and rats [85, 86], and CRHR1 signaling within the NI was strongly implicated in this process, as intra-NI CRHR1 antagonist injections strongly attenuated stress-induced reinstatement of alcohol seeking [36]. Numerous studies also report an increase in drug abuse vulnerability in human adults who experienced ELS [6]. In the current study, ELS significantly increased the number of NI CRHR1 mRNA-expressing neurons and the level of mRNA expression per cell, in accordance with reports that ELS disrupts the normal development of the CRH/CRHR1 system and leads to a net reinforcement of HPA axis activity in humans and rodents [44]. Thus, current data suggest the possible involvement of ELS-induced changes in NI CCK neuron anatomy and physiology, as well as CRHR1 expression in the NI, in the increased susceptibility to substances of abuse in adult individuals who have experienced childhood neglect.
Implications of ELS-induced TrkA mRNA dynamics in NI neurons
In the current study we also demonstrated that ELS increased the number of TrkA mRNA-expressing neurons and the level of TrkA mRNA per cell in the NI. Previous study reported that TrkA is selectively co-expressed with RLN3 in rat NI neurons [54], but co-expression with other neurotransmitters was not tested. Here, we demonstrated that TrkA mRNA was expressed by GABA/RLN3 and GABA/CCK neurons, as well as glutamate NI neurons. Notably however, we observed that the ELS-induced upregulation of TrkA mRNA occurred only in GABA, and not glutamate NI neurons, indicating a possible functional distinction between these neuronal subpopulations. Indeed, a recent study reported that in pontine central gray (PCG), of which NI is an integral part, GABA and glutamate neurons encode positive and negative valence, respectively, and that activation of these two populations resulted in preference (GABA) and avoidance behavior (glutamate) [87]. Our findings indicate that NI GABAergic and glutamatergic neuronal populations exhibit differential susceptibility to ELS, which may contribute to the atypical processing of valence-related information, a phenomenon documented in subjects with a history of ELS [88].
Several studies have shown that alcohol dependence is associated with alterations in plasma levels of NGF [89] and after alcohol withdrawal a rapid increase in the serum NGF concentration was observed [90–92]. It was also shown that ELS causes an increase in the level of NGF in the hippocampus, cerebral cortex and hypothalamus [93, 94], leading to impaired synaptic plasticity and behavioral deficits [95]. These data suggest that anxiety resulting from both ELS and alcohol use and withdrawal triggers NGF release. Notably, NGF increases TrkA expression [96], and our current data indicate that TrkA mRNA in the NI is increased by ELS, which together with an increase in CRHR1 mRNA expression, may translate into an altered sensitivity of the NI to stress in adulthood. Moreover, the increased basal expression of TrkA resulting from ELS may contribute to the augmentation of alcohol withdrawal-induced anxiety and promote stress-induced relapse of its consumption.
Additionally, ELS-induced upregulation of TrkA mRNA expression may be associated with enhanced protective mechanisms within the stress-sensitive NI neurons, as an established function of NGF is to facilitate neuronal survival [97]. However, the maladaptive or recurrent activation of NGF synthesis during the early postnatal period may impact stress sensitivity in adulthood and heighten susceptibility to stress-related psychopathology [47].
Concluding remarks
Collectively, the current findings advance our understanding of the neural mechanisms underlying ELS-related neuronal impairments, and together with previous data indicating the NI as a structure critically involved in contextual memory control [22] and stress-induced reinstatement of alcohol seeking [36, 41, 42], position the NI as a possible node linking ELS with compromised stress responses and greater vulnerability to substance abuse.
Supplementary information
Supplementary Materials Supplementary Table 9
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sholl DA. Dendritic organization in the neurons of the visual and motor cortices of the cat. J Anat 1953;87:387–406.PMC 124462213117757 · pubmed ↗