“A Tale Preserved in a Museum”: The Long-Awaited Discovery of Genitocotyle necromnemos n. sp. (Trematoda: Opecoelidae) from the Cardinal Fish Apogon imberbis (Linnaeus) in the Western Mediterranean
Fatima-Zohra Zedam, Chahinez Bouguerche, Fadila Tazerouti

TL;DR
A new species of trematode, Genitocotyle necromnemos, was discovered using museum specimens and new collections, clarifying a long-standing taxonomic issue.
Contribution
The discovery and description of Genitocotyle necromnemos n. sp., resolving taxonomic ambiguity through museum and new specimens.
Findings
Genitocotyle necromnemos n. sp. is morphologically distinct with larger body size and more numerous eggs.
The new species clarifies the identity of museum specimens collected in 1951.
The study emphasizes the role of museum collections in resolving taxonomic uncertainties.
Abstract
Opecoelids, a diverse group of digenean trematodes, pose a taxonomic challenge due to morphological similarities. The genus Genitocotyle is little studied with only five recognized species. Recent phylogenetic advancements have refined Opecoelidae classification, recognizing 15 subfamilies, including opecoelins, distinguished by the absence of a cirrus sac and a canalicular seminal receptacle. Within this subfamily, the genus Genitocotyle remains understudied, with only five recognized species. The aim of this study is to describe a new species and to clarify a long-standing taxonomic ambiguity. Two historical specimens of Genitocotyle from Apogon imberbis off Madeira, preserved in the Natural History Museum, London, were examined alongside newly collected specimens from Algerian waters. Comparative morphological analyses, including illustrations and morphometric measurements, were…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
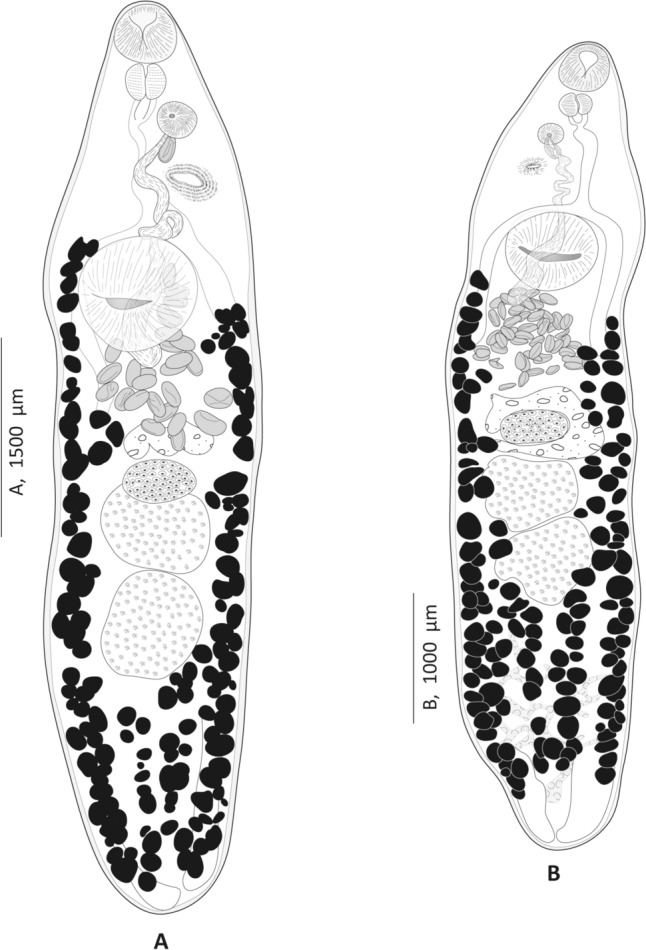 Figure 1
Figure 1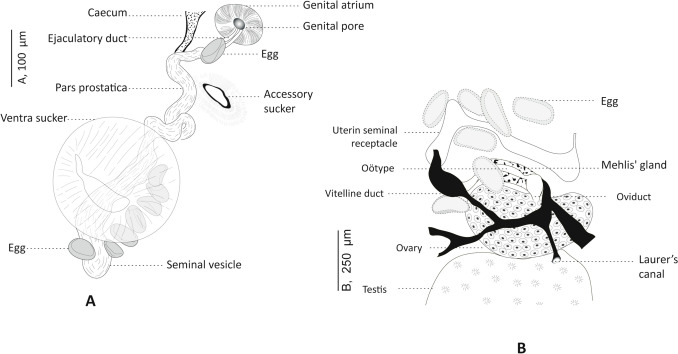 Figure 2
Figure 2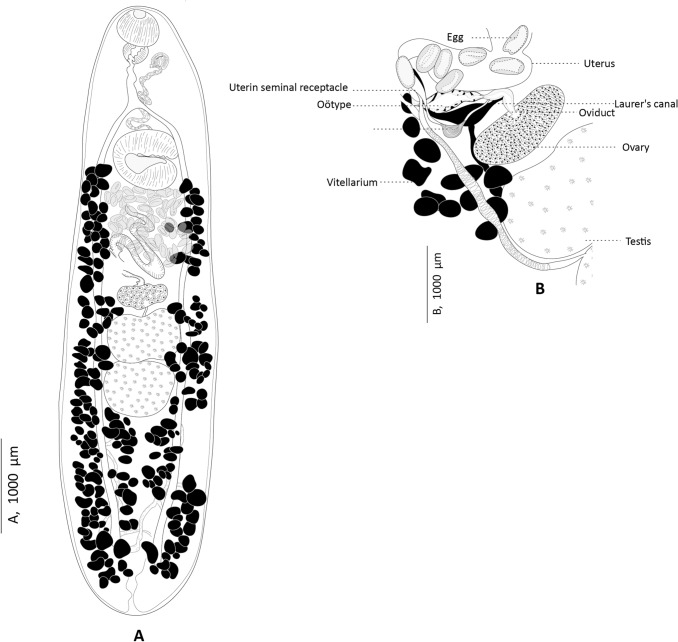 Figure 3
Figure 3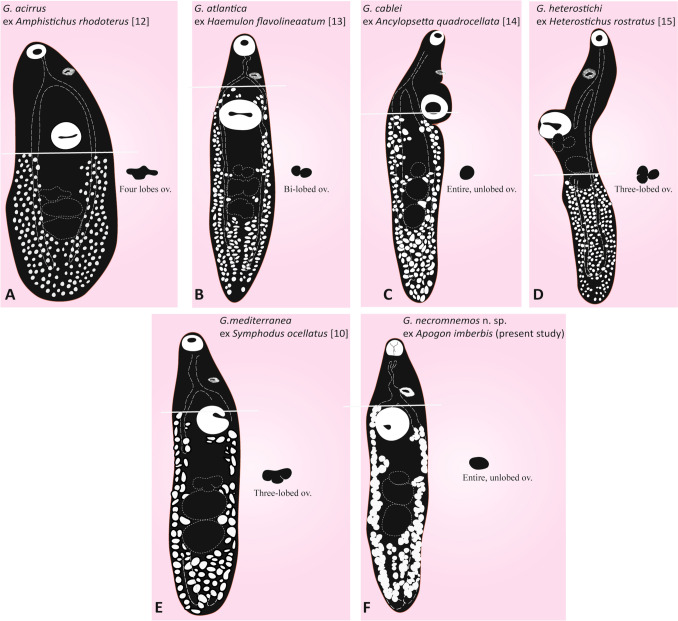 Figure 4
Figure 4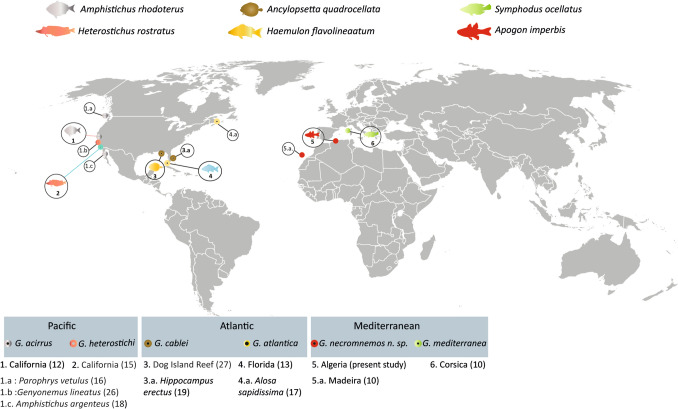 Figure 5
Figure 5- —Swedish Museum of Natural History
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasite Biology and Host Interactions · Helminth infection and control · Parasites and Host Interactions
Introduction
The opecoelids belong to one of the most taxonomically confusing groups of digeneans [1]. The family Opecoelidae Ozaki, 1925 comprises a diverse group of trematodes that parasitize marine and freshwater fishes [2]. Although the opecoelids are a large and evolutionarily derived group, the morphology of the adults is not very diverse or specialized [3]. Since the early 1980 s, the organization of Opecoelidae has been primarily based on classification hypothesis with four-subfamilies (see Martin [4]), with a lack of an appropriate subfamily classification [5]. In recent years, a new subfamily classification for Opecoelidae has been developed that reflects the phylogeny and provides practical taxonomic subdivisions. The proposed classification includes at least 15 subfamilies, based on phylogenetic relationships, adult morphology, and life-cycle ecology [3, 6–8].
The subfamily Opecoelinae Ozaki, 1925 is defined for species with reduced or absent cirrus sac and lacking a canalicular seminal receptacle [1]. It currently comprises 22 recognized genera [9]. Despite the general morphological uniformity, genera within this subfamily differ primarily by variations in the gut structure of the respective species (many exhibit configurations other than blind caeca), the shape and characteristics of the ventral sucker (e.g. the presence of papillae or a stalked form), and the distribution of the vitelline follicles [1]. Three genera, Anisoporus Ozaki, 1928, Opecoeloides Odhner, 1928 and Genitocotyle Park, 1937, in which the corresponding species are characterised by an unusual accessory sucker located between the genital atrium and the ventral sucker. The main differences between these genera lie in the termination of the intestinal caeca. In Anisoporus spp. the caeca open through a single anus, whereas in Opecoeloides spp. they connect to the base of the excretory vesicle and form a uroproct. In contrast, the caeca of Genitocotyle spp. are blind-ended [10].
To date, Genitocotyle comprises only five species [11]: (i). G. acirrus Park, 1937, the type species described from the stomach and upper intestine of the redtail surfperch Amphistichus rhodoterus (Agassiz) (Embiotocidae) off California (Eastern Central Pacific) [12]; (ii) G. atlantica Manter, 1947, described from the intestine of various fish families, off Florida (Western-Central Atlantic) [13]; (iii) G. cablei Park, 1937, found in the intestine of the ocellated flounder Ancylopsetta quadrocellata Gill (Paralichthyidae), off Dog Island Reef, Florida (Western-Central Atlantic) [14]; (iv) G. heterostichi Montgomery, 1957 described from the intestine of the giant kelpfish Heterostichus rostratus Girard (Clinidae) off La Jolla, California (Eastern Central Pacific) [15]; and (v) G. mediterranea Bartoli, Gibson and Riutort, 1994 described from the intestine of the ocellated wrasse Symphodus ocellatus (L.) (Labridae) off Corsica, France (Western Mediterranean) [10].
Genitocotyle spp. are distinguished by several morphological criteria, including the distribution of vitelline follicles, the extent of their yolk fields and the dimensions and shape of specific organs such as ovary and testes. Additional distinguishing features include the number of eggs, the presence of elongated seminal vesicles, and the length of the post-testicular region. These characteristics are sufficient to distinguish the species within this genus and help to characterise new species within this genus [10, 12–19]
Only one species of Genitocotyle has been recorded from the Mediterranean Sea—G. mediterranea [10]. However, two Genitocotyle specimens from a cardinal fish, Apogon imberbis (Apogonidae) (originated from Madeira, but died in 1951 in the aquarium of London Zoo), preserved in the Natural History Museum, London (NHMUK), showed similarities with G. mediterranea, but had notable differences, such as a larger size, more numerous eggs, a longer post-testicular zone and a seminal vesicle apparently restricted to the forebody [10]. Their conspecificity with G. mediterranea remained uncertain and required fresh material for confirmation [10]. Herein, we address the long-standing enigma of these digeneans. Although they were suspected and published as potentially new species, they remained undescribed due to limitations. We formally describe them as a new species of Genitocotyle based on the study of the museum specimens and freshly collected specimens from A. imberbis off Algeria.
Materials and Methods
From 2020 to 2024, a total of 218 specimens of A. imberbis were obtained from local fishermen in Dellys and Tamentfoust, Algeria, Western Mediterranean. Fish were transferred to the laboratory immediately after capture and identified using keys [20]. The gastrointestinal tract was removed and examined for trematodes using a stereomicroscope (Carl Zeiss^™^ 2000 Stereomicroscope, Germany) for the presence of helminths. Trematodes were removed alive using fine dissecting needles and heat-killed. Due to the need for detailed morphological comparison, all Genitocotyle specimens were flattened during preparation and then fixed in 70% ethanol with Bouin’s fluid [10]. While this method is not ideal for preservation [21], it was necessary to facilitate accurate comparison of key morphological features across species. Whole mounts were stained with boracic carmine, dehydrated through a graded ethanol series (70, 96 and 100%), cleared in clove oil, and mounted in Canada balsam [22]. Drawings were made with the aid of a Leitz microscope (USTHB, Algeria) and a Nikon Eclipse i80 microscope with DIC (differential interference contrast) (SMNH, Sweden) equipped with a drawing tube, scanned and redrawn with Adobe Illustrator (CS5). Measurements of whole-mounts are in micrometres and indicated as the range followed by mean values in parentheses. Additionally, two specimens of Genitocotyle sp. from the intestine of A. imberbis off Madeira (NHMUK Coll. No. 1980.7.17.228) [10] were examined for comparison. To compare morphological and anatomical features across Genitocotyle spp., figures in the global literature were extracted from published PDF files following Bouguerche et al. [23]. The following abbreviations are used: SMNH, Swedish Museum of Natural History, Stockholm, Sweden; NHMUK, Natural History Museum, London, UK.
Results
Family Opecoelidae Ozaki, 1925
Subfamily Opecoelinae Ozaki, 1925
Genus Genitocotyle Park, 1937
Genitocotyle necromnemos n. sp. (Figs. 1, 2, 3)Fig. 1Genitocotyle necromnemos n. sp. from the intestine of Apogon imberbis, off Algeria. A Body, ventral view. B Body, dorsal viewFig. 2Genitocotyle necromnemos n. sp. from the intestine of Apogon imberbis, off Algeria. A Ventral view of terminal genitalia. B Dorsal view of the female reproductive systemFig. 3Genitocotyle necromnemos n. sp. from the intestine of Apogon imberbis, off Madeira, historical specimens from the Natural History Museum (NHM 1980.7.17. 228). A Body, dorsal view. B Ventral view of the female reproductive system
Synonyms: Genitocotyle sp. sensu Bartoli, Gibson [10].
Type-host: Apogon imberbis (Kurtiformes: Apogonidae), the cardinal fish.
Type-locality: Dellys, 36.912984° N, 3.928383° E, Algeria, Western Mediterranean.
Other localities: Tamentfoust, 36.803675° N, 3.231279° E, Algeria, Western Mediterranean and Madeira, Eastern Central Atlantic [10].
Site in host: Intestine.
Infection details: Prevalence: 21.6 %; mean abundance: 0.22.; mean intensity: 3.04.
Type specimens: Holotype (SMNH Type-9988) and five paratypes (SMNH Type-9989–9993) deposited in the Type collections of the Swedish Museum of Natural History (SMNH), Stockholm, Sweden. 10 paratypes (LBEIG-ApGn-L7-ApGn-L16) deposited in the collections of Laboratoire de Biodiversité et Environnement, Interactions—Génomes, Université des Sciences et de la Technologie Houari Boumediene (USTHB), Algiers, Algeria.
Vouchers: Two specimens of Genitocotyle sp. ex Apogon imberbis off Madeira [NHMUK Coll. No. 1980.7.17.228].
ZooBank registration: urn:lsid:zoobank.org:pub:B2E79107-1C21-4A27-9EC1-2453C9DA42A2.
Etymology: The species name “necromnemos” is derived from two Greek words: necro-, meaning “dead” or “from death,” referencing the long-standing mystery surrounding these specimens that originated off Madeira and died in 1951 in the aquarium of London Zoo) and preserved the Natural History Museum (London); and -m**nemos, meaning"memory"alluding to the historical nature of these specimens, which had remained undescribed and largely forgotten until their rediscovery and formal description in this study.
Description
[Based on 20 whole- flattened gravid specimens; measurements in Table 1].Table 1. Morphomotry of Genitocotyle species. ^1^ Two specimens, previously annotated as Genitocotyle sp. from Apogon imberbis off Madeira, from the Natural History Museum. London [NHM 1980. 7. 17. 228]. ^2^ Other hosts are Carapus bermudensis (Carapidae), Malacoctenus macropus, (Labrisomidae), Opistognathus sp. (Opistognathidae), Syngnathus robertsi (Syngnathidae) (Manter, 1947)G. necromnemos n. sp.G. mediterraneaG. cableiG. heterostichiG. acirrusG. atlanticaHostApogon imberbis(Apogonidae)Symphodus ocellatus(Labridae)Ancylopsetta quadrocellata(Paralichthyidae)Heterostichus rostratus(Clinidae)Amphistichus rhodoterus(Embiotocidae)Haemulon**flavolineaatum^2^(Haemulidae)LocalityDellys, Tamentfoust Algeria, WMMaderia, Portugal, NEACorsica, France, WMDog Island Reef, WCALa Jolla, California, ECPDillon’s Beach, California, ECPTortugas, Florida, WCANo. of specimens2021021030Reference.Present studyPresent study^1^Bartoli et al. [10]Nahhas and Short [14]Montgomery [15]Park [12]Manter [13]Body L.1121–3408 (1994, n = 20)1939, 2425746–2201 (1307)2700–29301190–17302580 (2030–3480)892–1530Body W.222–723 (407, n = 20)457, 500213–421 (284)567–600170–2808700 (7500–1130)280–345Forebody280–789 (450, n = 20)400, 410266–576 (364)–310–530–Hindbody658–2378 (1337, n = 20)1310, 1790522–1396 (816)––––Oral sucker L.81–152 (117, n = 20)130, 13374–108 (89)165–18580–902000 (1700–2400)0.064–0.1 64–120Oral sucker W.75–174 (119, n = 20)140, 17380–128 (101)70–90Ventral sucker L.149–353 (221, n = 20)210, 254130–217 (170)268–294140–1703700 (3400–4200)96–200Ventral sucker W.141–334 (219, n = 20)230, 269130–217 (169)140–170Prepharynx L.9–27 (16, n = 15)11–39 (17)Accessory sucker L.27–79 (37, n = 20)90, 8343–98 (64)70–791800 (1600–1900)Accessory sucker W.7–60 (42, n =20)100, 95Pharynx L.51–147 (74, n = 20)80, 8046–72 (55)15530–601400 (110–180)48–54Pharynx W.49–128 (82, n = 20)78, 8056–93 (71)5043–48Oesophagus L.85–276 (186, n = 15)–91–228 (129)294–3604–5 times pharynx L.260 (150–380)Genital atrium–30–80 (51)Anterior testis L.85–374 (192, n = 20)129, 14598–228 (144)232–309210 (140–300)Anterior testis W.118–470 (246, n = 20)222, 240130–304 (203)360 (290–530)Posterior tests L.79–401 (206, n = 20)147, 19898–250 (160)290 (170–410)Posterior tests W.126–497 (244, n = 20)231, 218130–271 (193)360 (230–470)Ovary L.48–213 (93, n = 20)75, 7950–108 (74)155–180140 (60–200)Ovary W.61–250 (147, n = 20)162, 197102–228 (152)260 (200–380)Number of egg7–39 (18)6–12Egg L.41–61 (52, n = 18)46, 5049–59 (53)56–6466–7070–8052–58Egg W.20–45 (30, n = 18)28, 2924–32 (28)31–3645–5530–4026–30Ventral sucker to ovary142–644 (311, n = 20)310, 450109–325 (169)Ventral sucker to anterior testis190–863 (414, n = 20)400, 550130–434 (239)Post–testicular L.262–856 (511, n = 20)620, 840174–477 (279)510–1020Sucker-ratio1:0.53–0.52 (0.51)1:1.67–2.12 (1.91)1:1.54–1.621:1.8–1.911:1.45–1.8 (1.5)Oral sucker/pharynx ratio1:1.53–1.35 (0.34)1:0.53–0.69 (0.62)Forebody/hindbody ratio1:0.42–0.26 (0.34)1:1.71–2.54 (2.20)Forebody, L. of body ratio1:0.25–0.23 (0.24)1:3.2–4 (3.7)Forebody/Body L. (%)25–24% (23; n = 20)36–26 (28%)26–31%1/4 body L. (25%)Hindbody/Body L. (%)59–70% (67; n = 20)70–63 (62%)––Ventral sucker to ovary/Body L.13–19% (16; n = 20)14–15 (13%)Ventral sucker to anterior testis/Body L. (%)17–25% (21; n = 20)17–20 (18%)Post–testicular L./Body L. (%)23–25% (26; n = 20)23–22 (21%)43–59 (51%)All Measurements are in micrometres and indicated as the range followed by mean values in parentheses for some speciesWM Western Mediterranean, NEA Northeast Atlantic, WCA Western-Central Atlantic, ECP Eastern Central Pacific, L Length, W Width*Diameter
Body elongate, dorsoventrally flattened, widest at level of ventral sucker, rounded posteriorly. Forebody and hindbody occupying 23–25 (24; n = 20) and 59–70 (64; n = 20) % of body length, respectively. Post-testicular region shorter than forebody. Tegument thick, unarmed. Oral sucker ventro-subterminal. Ventral sucker located at limit of anterior and middle thirds of body, rounded, larger than oral sucker; ventral sucker to oral sucker length ratio 2:1.8–2.3 (2; n = 20), width ratio 1:1.87–1.91 (1.89; n = 20).
Accessory sucker present, without a limiting membrane, approximately midway between genital atrium and ventral sucker; aperture transverse, elliptical with visible radiating muscles. Pre-pharynx short. Pharynx large, ovoid. Esophagus long, thin-walled, bifurcating at level of anterior margin of ventral sucker. Caeca thick-walled, terminating blindly near posterior end of body.
Testes 2, tandem, entire, post-ovarian and located in mid-hindbody. Cirrus-sac absent. Vasa efferentia uniting to form vas deferens at base of seminal vesicle. Seminal vesicle long, tubular, oval or contoured, extending posteriorly to ventral sucker. Pars prostatica cylindrical. Genital atrium spherical, thick-walled, halfway between pre-pharynx and accessory sucker. Ejaculatory duct long, opening into genital atrium.
Ovary median, unlobed, pre-testicular, transversely elongated. Oviduct short. Laurer's canal connected to oviduct, opening dorsally to ovary. Oötype voluminous and surrounded by Mehlis’ gland, pre-ovarian. Canalicular seminal receptacle absent. Seminal receptacle uterine present. Uterus pre-ovarian, sinuous, extending to anterior edge of ventral sucker, ending with metraterm; metraterm thick-walled, opening into genital atrium. Eggs ovoid, few in numbers in uterus (around 30). Vitellarium follicular, vitelline follicles numerous and voluminous; ventrally, vitelline fields extending from anterior edge of acetabulum to posterior end of body, merging in post-testicular area; dorsally, vitelline fields extending from posterior edge of ventral sucker to two thirds of post-testicular area; transverse vitelline ducts situated in post-testicular region.
Remark
Two specimens of Genitocotyle sp. collected from A. imberbis off Madeira (see above) show similarities with G. mediterranea as noted by Bartoli and Gibson [10], but show notable differences, such as larger size, more numerous eggs in the uterus, a longer post-testicular zone and a seminal vesicle apparently restricted to the forebody. In this study, we re-examined these two specimens, and our analysis presented through illustrations and measurements confirms their conspecificity (Fig. 3; Table 1). Notably, both populations of Genitocotyle ex A. imberbis (museum specimens off Madeira and newly collected specimens off Algeria) share the extent vitellarium, supporting their conspecificity and classification as the same species, described here as G. necromnemos n. sp.
Discussion
To date, there are four nominal species of Genitocotyle : G. acirrus and G. heterostichi, occur in the Eastern Central Pacific [12, 18, 25, 26]; G. cablei and G. atlantica occurring in the Western-Central Atlantic [13, 17, 19, 27]; and G. mediterranea known only from Mediterranean waters [10]. Morphologically, G. necromnemos n. sp. ex A. imberbis off Algeria can be distinguished from G. acirrus in that the vitelline follicles extend into the forebody (vs. not surpassing the level of ventral sucker) and by the possession of an unlobed ovary (vs. lobed, with three lobes). Genitocotyle necromnemos n. sp. is readily distinguished from G. atlantica in having an unlobed ovary (vs. distinctly bilobed ovary in G. atlantica). Additionally, G. atlantica is readily differentiated from G. necromnemos n. sp. by the testes being wider than they are long and their outline is irregular or lobed, the post-testicular distance being approximately equal to or slightly greater than forebody length, and by having an ovoid ventral sucker. Genitocotyle cablei is similar to G. necromnemos n. sp. in that the ovary is unlobed, but differs in the extension of vitelline follicles (from the posterior end of the body to the posterior margin of the ventral sucker in *G. cablei *vs. extending to the anterior margin of the ventral sucker in G. necromnemos n. sp.), and by the post-testicular distance being longer than the length of the forebody in G. cablei. Genitocotyle heterostichi is easily distinguished from Genitocotyle necromnemos n. sp. in that the vitelline follicles reach the anterior margin of the posterior testis, an ovary with three lobes of ovoid shape, in having a greater post-testicular length, about 1/2 of the body, greater than forebody length, and by the seminal vesicle that does not extend posteriorly to the acetabulum.
Genitocotyle mediterranea resembles G. necromnemos n. sp. in that the vitelline follicles extend from the posterior end of the body to the anterior margin of the ventral sucker. Genitocotyle mediterranea differs from G. necromnemos n. sp. in having an ovary distinctly or indistinctly three-lobed (vs. unlobed in G. necromnemos n. sp.), having a transversely elongate ovary; median lobe posteriorly oriented and post-testicular length, about 1/4 of the body, less than the length of the forebody.
A comparative diagram of six Genitocotyle species (Fig. 4) suggests that the distribution of the vitelline follicles is a useful character to distinguish Genitocotyle species.
In G. acirrus, the vitelline follicles reach as far as the posterior edge of the ventral sucker (Fig. 4A). In G. atlantica, the vitelline follicles extend to the intestinal bifurcation (Fig. 4B). In G. cablei, the vitelline follicles reach the posterior edge of the ventral sucker (Fig. 4C). In G. heterostichi, the vitelline follicles extend to the posterior testis (Fig. 4D). In G. mediterranea, the vitelline follicles extend to the anterior edge of the acetabulum (Fig. 4E). In G. necromnemos n. sp., the vitelline follicles extend to the intestinal bifurcation (Fig. 4F).Fig. 4. Diagram showing the differences among Genitocotyle species in distribution of the vitelline follicles and shape of the ovary. A, Genitocotyle acirrus Park, 1937 ex Amphistichus rhodoterus [12]. B, Genitocotyle atlantica Manter, 1947 ex Haemulon flavolineaatum [13]. C, Genitocotyle cablei Nahhas and Short, 1965 ex *Ancylopsetta quadrocellata *[14]. D, *Genitocotyle heterostichi *ex Heterostichus rostratus Montgomery, 1957 [15]. E, Genitocotyle mediterranea Bartoli, Gibson and Riutort, 1994 ex Symphodus ocellatus [10]. F, Genitocotyle necromnemos n. sp. ex Apogon imberbis (present study)
We also tried to test the usefulness of the organization and shape of the ovary, and in G. acirrus, the ovary is clearly or indistinctly four-lobed (Fig. 4A). In G. atlantica, the ovary is bi-lobed (Fig. 4B). In G. cablei, the ovary is rounded and not lobed (Fig. 4C). In G. heterostichi, the ovary is ovoid with three lobes (Fig. 4D). In G. mediterranea, the ovary is distinctly or indistinctly three-lobed (Fig. 4E). In G. necromnemos n. sp., the ovary is ovoid and unlobed (Fig. 4F). These differences in the extent of the vitelline follicles and the morphology of the ovary highlight the usefulness of these features in distinguishing among Genitocotyle spp.
Conclusion
In this study, we described G. necromnemos n. sp., a new species of digenean based on the re-examination of museum specimens and freshly collected material from Apogon imberbis off Algeria. Our findings confirm that this species is distinct from other members of the genus Genitocotyle based on key morphological features, including the unlobed ovary and the extension of the vitelline follicles. The comparative analysis of these specimens, alongside other described species, highlights the importance of the distribution of the vitelline follicles and the organization of the ovary as distinguishing characters within the genus. This study also confirms G. necromnemos n. sp. as the first Genitocotyle species described from an apogonid host and provides a clearer understanding of the diversity within this genus. Further research on the ecological and zoogeographical aspects of this species will contribute to a more comprehensive understanding of Genitocotyle species’ host specificity and distribution (Fig. 5).Fig. 5. Geographic distribution of Genitocotyle species including the type hosts and type localities. Numbers in bold indicate type hosts mentioned in the original descriptions; numbers followed by letters represent secondary records. All references for the records are given in parentheses
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wo RMS (2025) Opecoelinae Ozaki, 1925. https://www.marinespecies.org/aphia.php?p=taxdetails&id=725010 Accessed 24 Mar 2025
- 2Wo RMS (2025 a) Genitocotyle Park, 1937. https://www.marinespecies.org/aphia.php?p=taxdetails&id=108527 Accessed 24 Mar 2025