Bibliometric analysis combined with visualization on universal trends and hot topics of LOX family in human diseases: 1995 to 2025
Tingting Yang, Heng Li, Wenxuan Zhou, Ning Zhang, Zhenyu Tian, Heming Wang, Yuyan Feng, Yunguang Chen, Zhen Wang

TL;DR
This paper analyzes 30 years of research on the LOX family of enzymes, showing their role in diseases and highlighting trends in scientific interest and potential future directions.
Contribution
The study provides a comprehensive bibliometric and visual analysis of LOX research trends from 1995 to 2025.
Findings
There were 9,261 LOX-related publications identified, with a significant increase in the past decade.
The United States, China, and Japan led in publication output, with Harvard and Boston Universities being highly productive.
LOX research is linked to cancer, fibrosis, and cardiovascular diseases, with potential for new diagnostics and treatments.
Abstract
Lysyl oxidase (LOX) is crucial for modifying collagen and elastin, thereby preserving tissue integrity. Aberrant LOX activity has been associated with a multitude of health disorders, including cutaneous, pulmonary, fibrotic, cardiovascular diseases, and cancer. In cancer, LOX can either promote or inhibit tumor development, and its expression level is closely correlated with patient prognosis. This research utilized data retrieved from the Web of Science Core Collection on May 30, 2025. The search strategies were crafted to target LOX – related terms while excluding irrelevant ones, and the data were limited to English – language articles. Over the past 30 years, 9261 LOX – related publications were identified. The number of articles exhibited an upward trend, especially in the past decade. The United States, China, and Japan were the leading countries in terms of publication output,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
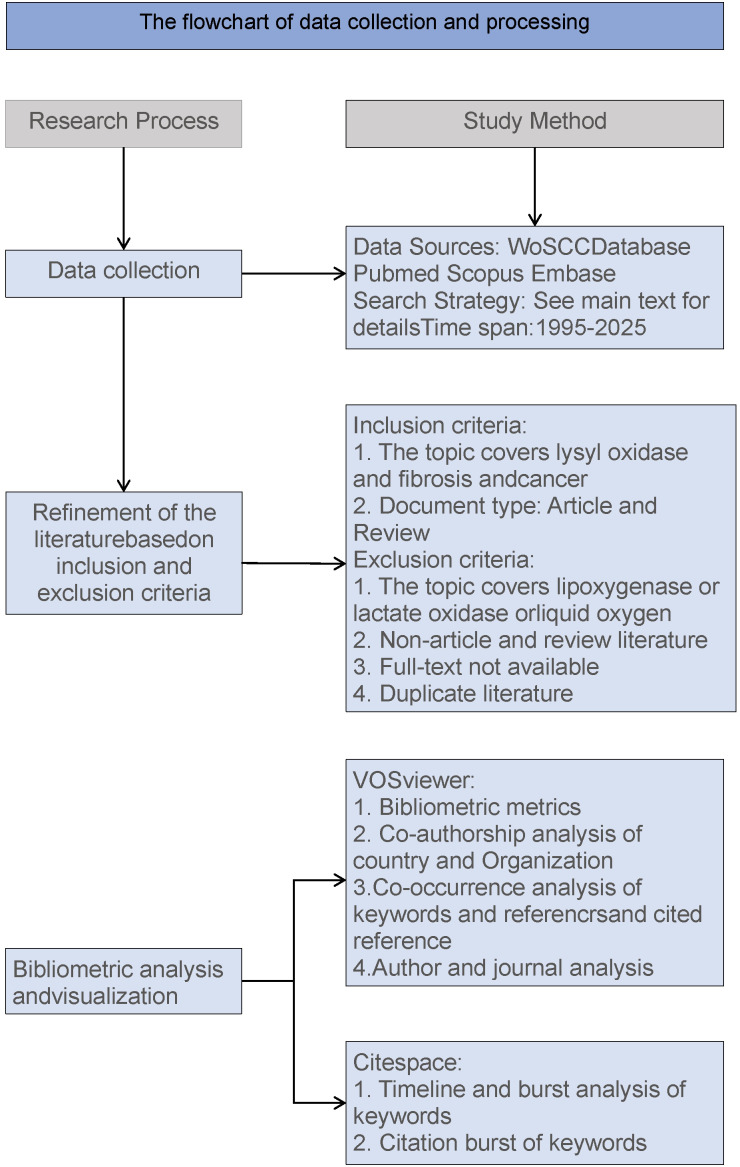 Figure 1
Figure 1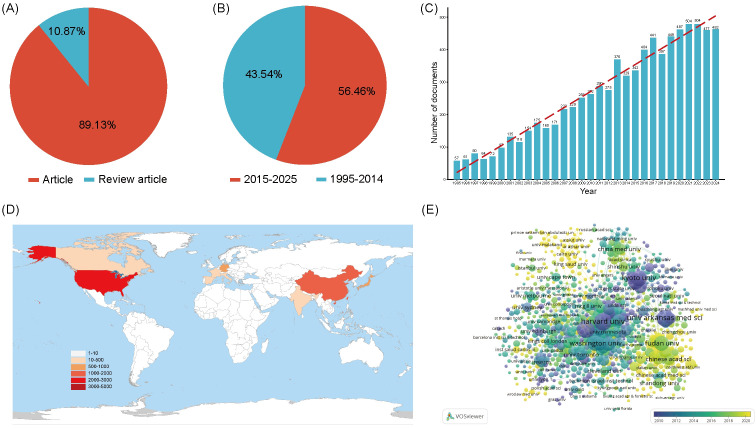 Figure 2
Figure 2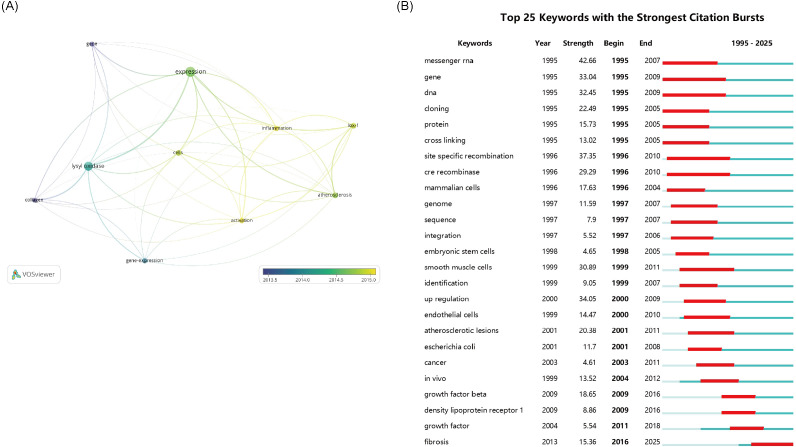 Figure 3
Figure 3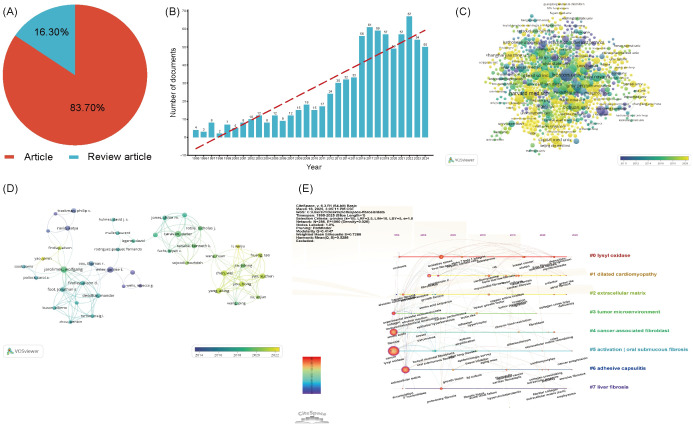 Figure 4
Figure 4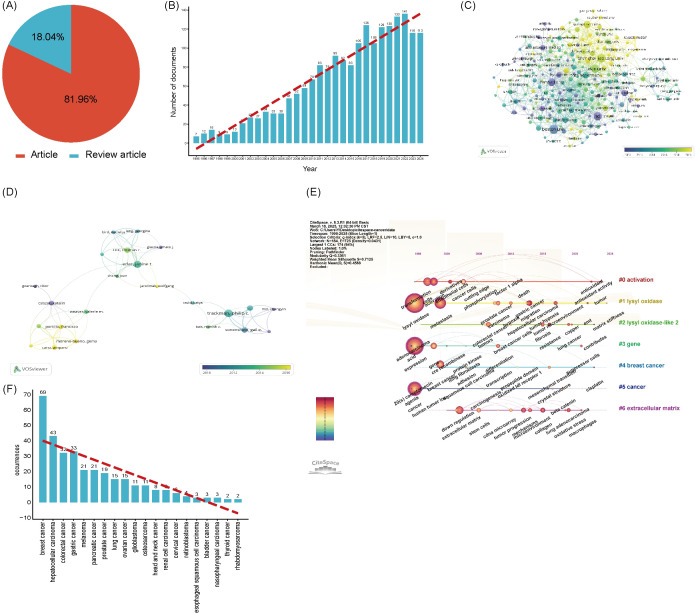 Figure 5
Figure 5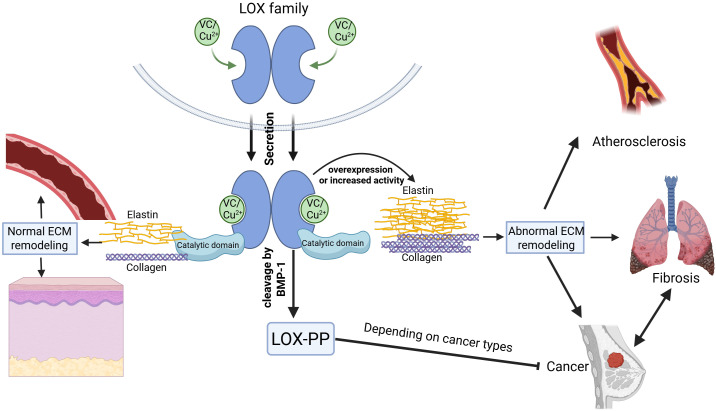 Figure 6
Figure 6| Gene Name | Alias | Primary Expressing Tissues/Cells | Core FunctionLOX |
|---|---|---|---|
| LOX/LOXL0 ( | LOX | Fibroblasts, endothelial cells, etc. | A classical lysyl oxidase that catalyzes the cross-linking of collagen and elastin to maintain ECM stability. |
| LOXL1 ( | LOX-like 1 | Skin, lungs, mammary glands, etc. | Involved in ECM cross-linking, regulating tissue fibrosis and tumor metastasis, including inhibiting breast cancer cell migration. |
| LOXL2 ( | LOX-like 2 | Tumor cells, fibroblasts | Promotes tumor angiogenesis, invasion, and metastasis, highly expressed in lung and ovarian cancers, associated with fibrosis. |
| LOXL3 ( | LOX-like 3 | Embryonic tissues, placenta, skin | Participates in embryonic development and skin homeostasis, potentially related to hair development. |
| LOXL4 ( | LOX-like 4 | Kidneys, placenta, digestive tract | Regulates kidney development and injury repair, potentially involved in ECM remodeling and fibrotic processes. |
| Rank | Country | Documents | Citations | Total link strength |
|---|---|---|---|---|
| 1 | USA | 2944 | 173096 | 1623 |
| 2 | China | 1698 | 36064 | 495 |
| 3 | Japan | 849 | 37539 | 429 |
| 4 | Germany | 576 | 30399 | 610 |
| 5 | France | 369 | 17462 | 344 |
| 6 | England | 364 | 19964 | 458 |
| 7 | Italy | 298 | 13353 | 294 |
| 8 | Canada | 292 | 12751 | 296 |
| 9 | India | 292 | 5453 | 150 |
| 10 | Spain | 225 | 12823 | 198 |
| Rank | Organization | Country | Documents | Citations | Total link strength |
|---|---|---|---|---|---|
| 1 | Harvard University | USA | 135 | 16998 | 264 |
| 2 | Boston University | USA | 133 | 7231 | 136 |
| 3 | University of Arkansas for Medical Sciences | USA | 128 | 7800 | 202 |
| 4 | Kyoto University | Japan | 110 | 7780 | 150 |
| 5 | National Cardiovascular Center | Japan | 106 | 9028 | 167 |
| 6 | Fudan University | China | 99 | 2869 | 84 |
| 7 | Washington University in St. Louis | USA | 98 | 4344 | 168 |
| 8 | Shanghai Jiao Tong University | China | 93 | 1952 | 106 |
| 9 | University of Michigan | USA | 82 | 7109 | 151 |
| 10 | University of Tokyo | Japan | 80 | 4276 | 121 |
| 11 | University of California, San Diego | USA | 78 | 4796 | 116 |
| 12 | Johns Hopkins University | USA | 75 | 4958 | 111 |
| 13 | University of Pennsylvania | USA | 75 | 10433 | 152 |
| 14 | Baylor College of Medicine | USA | 67 | 5721 | 138 |
| 15 | Sun Yat-sen University | China | 66 | 1754 | 83 |
| 16 | University of California, San Francisco | USA | 66 | 14282 | 160 |
| 17 | University of California, San Francisco | USA | 64 | 7360 | 125 |
| 18 | Chinese Academy of Sciences | China | 63 | 1922 | 77 |
| 19 | Vanderbilt University | USA | 63 | 3719 | 95 |
| 20 | China Medical University | China | 62 | 1557 | 177 |
| Rank | Country | Documents | Citations | Total link strength |
|---|---|---|---|---|
| 1 | USA | 335 | 23710 | 195 |
| 2 | China | 150 | 3702 | 54 |
| 3 | Germany | 59 | 4562 | 78 |
| 4 | Japan | 54 | 2660 | 41 |
| 5 | England | 50 | 3114 | 72 |
| 6 | France | 42 | 2372 | 48 |
| 7 | Spain | 41 | 2486 | 39 |
| 8 | Canada | 37 | 1841 | 40 |
| 9 | Australia | 33 | 2103 | 40 |
| 10 | Netherlands | 26 | 1288 | 28 |
| Rank | Organization | Country | Documents | Citations |
|---|---|---|---|---|
| 1 | Boston University | USA | 23 | 1160 |
| 2 | Harvard Medical School | USA | 19 | 1168 |
| 3 | University of Navarra | Spain | 14 | 1210 |
| 4 | University of Pennsylvania | USA | 14 | 5251 |
| 5 | Sichuan University | China | 13 | 151 |
| 6 | University of California, San Francisco | USA | 12 | 3724 |
| 7 | Massachusetts General Hospital | USA | 11 | 468 |
| 8 | National Taiwan University | Taiwan, China | 11 | 1534 |
| 9 | University of Oslo | Norway | 11 | 412 |
| 10 | Capital Medical University | China | 10 | 178 |
| 11 | Gilead Sciences, Inc. | USA | 10 | 1074 |
| 12 | Johns Hopkins University | USA | 10 | 570 |
| 13 | Medical University of South Carolina | USA | 10 | 687 |
| 14 | Pharmaxis Ltd | Australia | 10 | 359 |
| 15 | Shanghai Jiao Tong University | China | 10 | 167 |
| 16 | Technion – Israel Institute of Technology | Israel | 10 | 1163 |
| 17 | University of Kansas | USA | 10 | 346 |
| 18 | University of Michigan | USA | 10 | 1368 |
| 19 | University of Sydney | Australia | 10 | 829 |
| 20 | University of Arkansas for Medical Sciences | USA | 9 | 316 |
| Rank | Co-cited reference | Author | Publication date | Journal | Citations | Total link strength |
|---|---|---|---|---|---|---|
| 1 | Allosteric inhibition of lysyl oxidase-like-2 impedes the development of a pathologic microenvironment | Barry-Hamilton V | 2010 | Nature Medicine | 111 | 423 |
| 2 | Lysyl oxidase-like-1 dysregulation in hypertrophic cardiomyopathy | López B | 2010 | American Journal of Physiology-Heart and Circulatory Physiology | 87 | 197 |
| 3 | Lysyl oxidase: properties, regulation and multiple functions in biology biology” | Kagan HM | 2003 | Journal of Cellular Biochemistry | 85 | 259 |
| 4 | Lysyl oxidase: properties, regulation and multiple functions in biology | Smith-Mungo LI | 1998 | Matrix Biology | 71 | 180 |
| 5 | Lysyl oxidase inhibition ameliorates angiotensin II-induced cardiac remodeling | Liu SB | 2016 | FASEB Journal | 60 | 192 |
| 6 | Lysyl oxidase-like 2 is a key mediator in pancreatic cancer progression | Ikenaga N | 2017 | Gut | 57 | 220 |
| 7 | Abnormal deposition of collagen around hepatocytes in Wilson’s disease is associated with hepatocyte specific expression of lysyl oxidase and lysyl oxidase like protein-2 | Vadasz Z | 2005 | Journal of Hepatology | 53 | 218 |
| 8 | The tumour microenvironment: a novel target for cancer therapy | Barker HE | 2012 | Nature Reviews Cancer | 50 | 220 |
| 9 | Lysyl oxidase: an oxidative enzyme and effector of cell function | Lucero HA | 2006 | Cellular and Molecular Life Sciences | 50 | 223 |
| 10 | Matrix crosslinking forces tumor progression by enhancing integrin signaling | Levental KR | 2009 | Cell | 49 | 169 |
| 11 | Lysyl oxidases: a novel multifunctional amine oxidase family | Csiszar K | 2001 | Progress in Nucleic Acid Research and Molecular Biology | 46 | 189 |
| 12 | A molecular signature for metastasis in human solid cancers | Peinado H | 2005 | EMBO Journal | 46 | 202 |
| 13 | Targeting lysyl oxidase (LOX) overcomes chemotherapy resistance in triple negative breast cancer | Yang J | 2016 | Nature Communications | 42 | 141 |
| 14 | Comparative analysis of lysyl oxidase (LOX) family expression in carcinogenesis | Aumiller V | 2017 | Scientific Reports | 40 | 199 |
| 15 | A fluorometric assay for detection of lysyl oxidase enzyme activity in biological samples | Palamakumbura AH | 2002 | Analytical Biochemistry | 40 | 155 |
| 16 | The hypoxic cancer secretome induces pre-metastatic bone lesions through lysyl oxidase | Cox TR | 2013 | Cancer Research | 39 | 133 |
| 17 | Hypoxia-induced lysyl oxidase is a critical mediator of bone marrow cell recruitment to the neovasculature | Erler JT | 2009 | Cancer Cell | 39 | 141 |
| 18 | The expression of lysyl oxidase in oral squamous cell carcinoma | Trivedy C | 1999 | Journal of Oral Pathology and Medicine | 38 | 36 |
| 19 | Regulation of lysyl oxidase activity in cultured bovine aortic smooth muscle cells | Kagan HM | 1991 | American Journal of Respiratory Cell and Molecular Biology | 37 | 69 |
| 20 | Lysyl Oxidase Is Essential for Normal Development and Function of the Respiratory System and for the Integrity of Elastic and Collagen Fibers in Various Tissues | Mäki JM | 2002 | Circulation | 37 | 150 |
| Rank | Country | Documents | Citations | Total link strength |
|---|---|---|---|---|
| 1 | USA | 701 | 52026 | 394 |
| 2 | China | 506 | 13145 | 135 |
| 3 | Japan | 174 | 8375 | 83 |
| 4 | Germany | 105 | 4509 | 111 |
| 5 | England | 97 | 7200 | 127 |
| 6 | France | 87 | 5791 | 78 |
| 7 | India | 81 | 1754 | 28 |
| 8 | South korea | 79 | 3073 | 29 |
| 9 | Italy | 62 | 2730 | 49 |
| 10 | Canada | 54 | 3871 | 55 |
| Rank | Organization | Country | Documents | Citations | Total link strength |
|---|---|---|---|---|---|
| 1 | Boston University | USA | 48 | 1904 | 19 |
| 2 | Shanghai Jiao Tong University | China | 44 | 1191 | 21 |
| 3 | Fudan University | China | 42 | 1550 | 20 |
| 4 | National Cancer Institute | USA | 41 | 2158 | 16 |
| 5 | Harvard University | USA | 35 | 4355 | 26 |
| 6 | Sun Yat-sen University | China | 32 | 745 | 22 |
| 7 | Johns Hopkins University | USA | 26 | 2892 | 12 |
| 8 | The University of Tokyo | Japan | 24 | 1400 | 6 |
| 9 | Stanford University | USA | 22 | 5099 | 21 |
| 10 | Harvard Medical School | USA | 20 | 545 | 16 |
| 11 | Kyoto University | Japan | 20 | 1440 | 6 |
| 12 | University of Arkansas for Medical Sciences | USA | 20 | 1654 | 7 |
| 13 | University of California, San Francisco | USA | 20 | 8626 | 25 |
| 14 | University of Hawaii | USA | 20 | 1800 | 6 |
| 15 | The University of Texas MD Anderson Cancer Center | USA | 20 | 1634 | 22 |
| 16 | Northwestern University | USA | 19 | 1066 | 23 |
| 17 | University of Copenhagen | Denmark | 19 | 2349 | 13 |
| 18 | Yonsei University | South Korea | 19 | 544 | 10 |
| 19 | Central South University | China | 18 | 298 | 15 |
| 20 | Institut National de la Santé et de la Recherche Médicale | France | 18 | 1022 | 16 |
| Rank | Co-cited reference | Author | Publication date | Journal | Citations | Total link strength |
|---|---|---|---|---|---|---|
| 1 | Lysyl oxidase-related protein-1 promotes tumor fibrosis and progression | Erler JT | 2006 | Nature | 330 | 1831 |
| 2 | Lysyl oxidase-like 2 (LOXL2) modulates tumor-associated macrophage polarization and promotes breast cancer metastasis | Erler JT | 2009 | Cancer Cell | 210 | 1189 |
| 3 | Tumor-secreted LOXL2 activates fibroblasts through FAK signaling | Kagan HM | 2003 | Journal of Cellular Biochemistry | 178 | 1069 |
| 4 | The tumour microenvironment: a novel target for cancer therapy | Levental KR | 2009 | Cell | 177 | 1147 |
| 5 | Allosteric inhibition of lysyl oxidase-like-2 impedes the development of a pathologic microenvironment | Barker HE | 2012 | Nature Reviews Cancer | 175 | 1049 |
| 6 | Chromosomal localization of the lysyl oxidase gene (LOX) in mouse and human | Kirschmann DA | 2002 | Cancer Research | 155 | 1147 |
| 7 | The lysyl oxidase-like 2 enzyme regulates tumor angiogenesis and is a potential therapeutic target | Peinado H | 2005 | EMBO Journal | 129 | 1017 |
| 8 | Lysyl oxidases: a novel multifunctional amine oxidase family | Barry-Hamilton V | 2010 | Nature Medicine | 126 | 919 |
| 9 | Hypoxia-induced lysyl oxidase is a critical mediator of bone marrow cell recruitment to form the pre-metastatic niche | Lucero HA | 2006 | Cellular and Molecular Life Sciences | 110 | 755 |
| 10 | Lysyl oxidase is essential for hypoxia-induced metastasis | Csiszar K | 2001 | Progress in Nucleic Acid Research and Molecular Biology | 111 | 786 |
| 11 | Lysyl oxidase: properties, specificity, and biological roles inside and outside of the cell | Payne SL | 2005 | Cancer Research | 101 | 785 |
| 12 | A molecular role for lysyl oxidase in breast cancer invasion | Payne SL | 2007 | Journal of Cellular Biochemistry | 98 | 769 |
| 13 | Matrix crosslinking forces tumor progression by enhancing integrin signaling | Smith-Mungo LI | 1998 | Matrix Biology | 90 | 556 |
| 14 | Lysyl oxidase: an oxidative enzyme and effector of cell function | Cox TR | 2013 | Cancer Research | 83 | 533 |
| 15 | Lysyl oxidase regulates breast cancer cell migration and adhesion through a hydrogen peroxide-mediated mechanism | Peinado H | 2008 | Cancer Research | 83 | 709 |
| 16 | Lysyl oxidase regulates tumor cell survival through Akt-dependent and -independent mechanisms | Baker AM | 2011 | Journal of the National Cancer Institute | 79 | 604 |
| 17 | Lysyl oxidase-like 2 as a new regulator of epithelial-mesenchymal transition | Barker HE | 2011 | Cancer Research | 77 | 617 |
| 18 | LOXL2-mediated matrix remodeling in metastasis and mammary gland involution | Peng L | 2009 | Carcinogenesis | 77 | 650 |
| 19 | Lysyl oxidase activates cancer stromal cells and promotes gastric cancer progression | Akiri G | 2003 | Cancer Research | 76 | 566 |
| 20 | Lysyl oxidase: properties, regulation, and multiple functions in biology | Contente S | 1990 | Science | 72 | 445 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial metabolism and enzyme function · Medical and Biological Ozone Research
Introduction
The Lysyl oxidase (LOX) family of enzymes plays indispensable roles in the cross-linking and remodeling of the extracellular matrix (ECM), and participates in the post-translational modification of collagen and elastin (1). In mammals, the LOX family typically consists of four or five members: LOX, LOXL1, LOXL2, LOXL3, and LOXL4, depending on the nomenclature differences in the literature regarding LOX and LOXL0. Structurally, they are made up of three separate domains: an amino-terminal signal peptide that facilitates its secretion, a central catalytic domain that performs the oxidative deamination of lysine residues in collagens, and a carboxy-terminal regulatory domain (2–6). They share common features such as dependence on copper ions (Cu^2+^) and vitamin C as cofactors to catalyze the oxidation of lysine/hydroxylysine to aldehyde for ECM cross-linking. This unique structure enables LOX family to act as a critical regulator of tissue integrity, facilitating the formation of cross - linked collagen fibers that are essential for skin elasticity, vessel stability, and the strength of connective tissues (7–10)However, they differ in substrate preference, cellular signaling regulation, and expression during developmental stages. LOX primarily acts on collagen and elastin, while LOXLs can target a broader range of matrix proteins. LOXLs, especially LOXL2, can regulate signaling pathways in an enzyme-independent manner, whereas LOX function is more dependent on enzymatic activity. LOX is continuously expressed in adult tissues, while LOXLs are more active during embryonic development or tissue repair. Based on the characteristics of each LOX family member, we have compiled and described the LOX family members according to their molecular weight, primary tissue/cell expression, and core functions in Table 1.
LOX family of enzymes activity is crucial for preserving the structural integrity and mechanical properties of tissues like the skin, lungs, blood vessels, and other connective tissues (3–6, 8, 11). LOX facilitates the oxidation of lysine residues in proteins, leading to the formation of covalent cross-links essential for tissue elasticity and resilience (12). However, abnormal LOX activity has been associated with a broad spectrum of pathological conditions. For example, deficiencies in LOX or its isoforms are associated with severe cutaneous disorders like cutis laxa, characterized by skin fragility and reduced dermal elasticity due to impaired elastic fiber formation (13, 14). Similarly, LOX dysfunction is linked to pulmonary diseases, including alveolar wall destruction and impaired lung function, highlighting its importance in maintaining respiratory tissue architecture (15, 16). Moreover, altered LOX activity is involved in fibrotic diseases such as liver cirrhosis and pulmonary fibrosis, where excessive extracellular matrix (ECM) remodeling leads to tissue stiffening and organ dysfunction (17, 18). In addition to its role in connective tissues, LOX has attracted attention for its involvement in cardiovascular pathologies (19–21). Elastic fiber abnormalities caused by LOX deficiency can lead to vascular complications, such as aneurysms, emphasizing the enzyme’s critical function in maintaining vascular integrity (22–25).
Beyond these well - established roles, studies have also explored the potential of LOX in cancer biology (26). Emerging evidence suggests that LOX activity may contribute to tumor progression and metastasis by facilitating ECM remodeling, promoting cancer cell invasion, and enhancing angiogenesis (27, 28). Elevated LOX expression has been observed in various cancers, where it promotes ECM stiffening and the formation of a pro - tumorigenic niche (29, 30). Additionally, LOX activity triggers downstream signaling pathways like TGF-β and MAPK, which are strongly linked to epithelial-mesenchymal transition (EMT) and the preservation of cancer stem cells (31–33). LOX has also been shown to interact with key signaling pathways like focal adhesion kinase (FAK) - YAP/TAZ, which are crucial for tumor cell survival and proliferation (32, 34). Further clinical investigation into hepatocellular carcinoma (HCC) tissues revealed that HIF-1α is implicated in LOXL2 protein expression, correlates with poor prognostic factors, and drives processes such as EMT, migration, invasion, and vasculogenic mimicry in HCC cells by regulating LOXL2 (35). Additionally, lysyl oxidase (LOX) released from hypoxic breast tumor cells accumulates in premetastatic niches, cross-links type IV collagen within the basement membrane, and critically contributes to the recruitment of CD11b+ myeloid cells (36).In most cases, LOX is a pivotal factor in the restructuring of the tumor-related extracellular matrix, metastasis, and the premetastatic niche. These findings indicate that the LOX family could be a viable target for preventing and treating metastatic conditions. Nevertheless, LOX can also display antitumor effects in certain cancers, with its metabolic by-products potentially counteracting oncogenesis. For example, the expression levels of LOX are closely related to gastric cancer patient prognosis, with low expression usually associated with poorer clinical outcomes (37). LOX expression is downregulated in epidermal tumor cells of carcinomas but upregulated in the stroma adjacent to invasive tumor cells, and also they proved that LOX enzymatic activity is crucial for preserving the integrity of the dermis as well as maintaining the epidermal homeostasis but not for cancer progression (38). LOX mRNA is translated into an inactive LOX proprotein (pro-LOX) within the cytoplasm. Subsequently, pro-LOX is secreted from the cell, where the N-terminus of the protein is cleaved by BMP-1, resulting in the LOX propeptide (LOX-PP) and the mature, active LOX protein (39). Another study showed that 18-kDa propeptide domain of LOX (LOX-PP) from LOX strongly suppresses the invasive characteristics of lung and pancreatic cancer cells by targeting Bcl2 and NF-kappaB (40). Furtherly, LOX-PP expression was found to diminish the proliferative, migratory, anchorage-independent growth, and tumorigenic abilities of Ewing sarcoma in immunocompromised mice, while the C-terminal of LOX enzymatically active domain of LOX exerted opposing effects, indicating that LOX’s tumor-suppressive role is specifically attributable to its propeptide domain (41). This implies that treatments targeting LOX should be selective, targeting only those cancers where LOX promotes progression, potentially necessitating molecular diagnostic tools, and that LOX or its by-products could potentially serve as anticancer agents (10, 42, 43). Through an extensive literature review, this study aims to systematically analyze the current research status of LOX family and propose innovative and feasible future research directions by integrating it with hot topics in fibrotic diseases and cancer research. This will enhance comprehension of the mechanisms behind LOX’s involvement in tumor growth and advancement, offering fresh perspectives and possible targets for clinical treatment.
Materials and methods
Data acquisition and processing
The data employed for this bibliometric study were acquired from the Web of Science Core Collection (WoSCC) database, Scopus, PubMed, and Embase, incorporating the Science Citation Index Extended (SCIE), Social Science Citation Index (SSCI), and Emerging Sources Citation Index (ESCI). The information was obtained from the, Scopus, PubMed, and Embase from 1995 to 2025 on May 30, 2025. The search strategies were as follows:
1#. TS=“lysyl oxidase” OR “LOX” OR “LOXL1” OR “LOXL2” OR “LOXL3” OR “LOXL4” OR “lysyl oxidase like 1” OR “lysyl oxidase like 2” OR “lysyl oxidase like 3” OR “lysyl oxidase like 4”OR “Protein - lysine 6 - oxidase” OR “Copper - dependent amine oxidase”NOT TS=(“lipoxygenase” OR “lactate oxidase” OR “liquid oxygen”)
2#. TS=“lysyl oxidase” OR “LOX” OR “LOXL1” OR “LOXL2” OR “LOXL3” OR “LOXL4” OR “lysyl oxidase like 1” OR “lysyl oxidase like 2” OR “lysyl oxidase like 3” OR “lysyl oxidase like 4”OR “Protein - lysine 6 - oxidase” OR “Copper - dependent amine oxidase” AND TS=(“fibrosis” OR “fibrotic” OR “fibrogenesis”) NOT TS=(“lipoxygenase” OR “lactate oxidase” OR “liquid oxygen”)
3#. TS=“lysyl oxidase” OR “LOX” OR “LOXL1” OR “LOXL2” OR “LOXL3” OR “LOXL4” OR “lysyl oxidase like 1” OR “lysyl oxidase like 2” OR “lysyl oxidase like 3” OR “lysyl oxidase like 4”OR “Protein - lysine 6 - oxidase” OR “Copper - dependent amine oxidase”AND TS=(“cancer” OR “tumor” OR “neoplasm” OR “malignancy” OR “carcinoma”) NOT TS=(“lipoxygenase” OR “lactate oxidase” OR “liquid oxygen”)
The data were limited to English - language articles. After the above - mentioned selection process, a total of 8255 for 1# search strategy, 831 for 2# search strategy and 2041 for 3# search strategy papers according to the above strategies were obtained for bibliometric analysis (Figure 1).
The flowchart of data collection and progressing.
Bibliometric visualized analysis
VOSviewer (version 1.6.20; https://www.vosviewer.com/) was employed to conduct analyses of countries, organizations, journals, authors, co - cited authors, reference co - occurrences, and keyword co-occurrences (44). CiteSpace (version 6.3.R1; https://citespace.podia.com/) was used to analyze citation burst frequencies, keyword bursts (45). Additionally, Graphpad Prism 8 (https://www.graphpad.com/) was applied to analyze thosepublications on LOX from 1995 to 30 May, 2025.
Results
General literature trends on LOX from 1995 - 2025
By applying the search strategy TS=((“lysyl oxidase” OR “LOX” OR “LOXL1” OR “LOXL2” OR “LOXL3” OR “LOXL4” OR “lysyl oxidase like 1” OR “lysyl oxidase like 2” OR “lysyl oxidase like 3” OR “lysyl oxidase like 4”OR “Protein - lysine 6 - oxidase” OR “Copper - dependent amine oxidase”)NOT(lipoxygenase OR “lactate oxidase” OR “liquid oxygen”)) in WoSCC, Scopus, PubMed, and Embase database, a total of 9,261 LOX - related publications were identified over the past 30 years. Among these, 8,255 (89.13%) were articles, and 1,006(10.87%) were reviews (Figure 2A). The total number of articles in the past decade was 4,440, accounting for 56.46% of the total articles in the past 30 years (Figure 2B). The number of article - type publications from 1995 to 2025 showed an upward trend (Figure 2C). Before 2000, the annual publication count was less than 100. After 2013, there was a sharp rise in the number of relevant publications, indicating growing attention and prosperity in this field over the past 10 years.
(A) The proportion of articles in the field of LOX. (B) The proportion of articles in the field of LOX over the past decade. (C) The number of publications in the field of LOX by year. (D) The top 10 most productive countries/regions in the field of LOX. (E) Distribution of publications from different organizations in the field of LOX. The thickness of the connecting lines between the nodes reflects the strength of the interaction. The color of the node represents the average year of publication for the country which the node represents.
8,255 documents in total were published by 102 countries/regions. Figure 2D lists the top 10 productive countries/regions for LOX publications. The United States had the most publications and citations (n = 2,944, 35.7%), followed by China (n = 1,689, 20.5%) and Japan (n = 849, 10.3%). These three countries accounted for nearly 66.5% of the total publications (Table 2). The 8,255 analyzed documents were affiliated with 6,489 organizations. Table 3 shows the top 20 most productive institutions, with Harvard University and Boston University leading. Harvard University, the University of California, San Francisco, and the University of Pennsylvania had the highest citation counts (Table 3). Figure 2E depicts the overall connection strength and collaboration network among institutions.
Keyword analysis of hotpots on LOX in human diseases
Research themes and hotspots were identified through keyword analysis. The VOSviewer was applied to visualize the keyword network diagram for 8,255 articles. Specifically, 272 key keywords with a frequency of at least 40 were selected for visualization. Figure 3A shows an overlay map of the top 10 frequently occurring keywords, where the color corresponds to average publication year. Subsequently, disease - related keywords were extracted for further investigation. Clustering of these disease - related keywords resulted in 3 clusters, encompassing a total of 11 items. Among them, “cancer” and “fibrosis” appeared more than 440 times, likely representing prominent research topics. This study also presents the timeline viewer for “cancer” and “fibrosis” (Figure 3B).
(A) The network visualization map of disease-related keywords. (B) The 25 keywords with the Strongest Citation Bursts. The thickness of the connecting lines between the nodes reflects the strength of the interaction. The color of the node represents the average year of publication for the country which the node represents.
Analysis of literatures on LOX in fibrosis research
To conduct an in - depth analysis of LOX - related literature within the realm of fibrosis research, we implemented a comprehensive search strategy. The search terms were configured as follows: TS=“lysyl oxidase” OR “LOX” OR “LOXL1” OR “LOXL2” OR “LOXL3” OR “LOXL4” OR “lysyl oxidase like 1” OR “lysyl oxidase like 2” OR “lysyl oxidase like 3” OR “lysyl oxidase like 4”OR “Protein - lysine 6 - oxidase” OR “Copper - dependent amine oxidase” AND TS=(“fibrosis” OR “fibrotic” OR “fibrogenesis”) NOT TS=(“lipoxygenase” OR “lactate oxidase” OR “liquid oxygen”)with the time span ranging from 1995 to 2025. Over these 30 years, a total of 993 LOX - related publications were identified. Among them, 831 (constituting approximately 83.7%) were original research articles, while 162 (roughly 16.3%) were review articles, as presented in Figure 4A. Notably, the past nine years have witnessed a remarkable upsurge in studies focusing on LOX in the context of fibrosis, as vividly illustrated in Figure 4B.
(A) The proportion of articles in the field of LOX associated with fibrosis research. (B) The number of publications in the field of LOX associated with fibrosis research by year. (C) Distribution of publications from different organizations in the field of LOX associated with fibrosis research. (D) The co-occurrence author map of co-authors in the field of LOX associated with fibrosis research. (E) Timeline view of keywords analysis in the field of LOX associated with fibrosis research. The thickness of the connecting lines between the nodes reflects the strength of the interaction. The color of the node represents the average year of publication for the country which the node represents.
These 831 documents were sourced from 56 different countries and regions. The United States took the lead with 335 literatures, making up 40.3% of the total quantity, closely followed by China with 150 publications (18.1%). Together, these two countries contributed nearly 58.4% of all the relevant works, as detailed in Table 4. The 831 analyzed documents were affiliated with 1288 distinct organizations. Among the top 20 productive institutions, as presented in Table 5, Boston University (with 23 publications) and Harvard Medical School (19 publications) were particularly active. The University of Pennsylvania and the University of California, San Francisco boasted the highest citation counts, and their collaborative network is depicted in Figure 4C.
In the past decade, 5,438 authors made contributions to LOX and fibrosis - related articles. The most productive author had 10 publications. Core contributors were defined as those with at least two publications. Employing VOSviewer, a network visualization was constructed. Authors with three or more publications were selected for further in - depth analysis, including visualization, clustering, and mapping of co - author distribution. In the fibrosis - related LOX study, 37 authors publishing five or more papers. Javier Diez ranked first with 10 publications, trailed by Wolfgang Jarolimek (8) and caravan, peter (7), as shown in Figure 4D. Table 6 presents the top 20 co-cited references, with the most - cited one being from Nature Medicine in 2010 by Barry - Hamilton V. This reference explored LOX’s role in the tumor microenvironment, especially in metastasis and angiogenesis. Figure 4E showcases a keyword network for LOX research. “Lysyl oxidase” is interconnected with terms such as “dilated cardiomyopathy”, “tumor microenvironment”, and those related to fibrosis, vividly demonstrating LOX’s diverse functions across various medical conditions. Keywords like “cancer” and “extracellular matrix” stand out due to their high frequency, and the color - coded lines further reveal the complexity of the research landscape.
Analysis of literatures on LOX in cancer research
Our search strategy for LOX in cancer research was formulated as: TS=“lysyl oxidase” OR “LOX” OR “LOXL1” OR “LOXL2” OR “LOXL3” OR “LOXL4” OR “lysyl oxidase like 1” OR “lysyl oxidase like 2” OR “lysyl oxidase like 3” OR “lysyl oxidase like 4”OR “Protein - lysine 6 - oxidase” OR “Copper - dependent amine oxidase”AND TS=(“cancer” OR “tumor” OR “neoplasm” OR “malignancy” OR “carcinoma”) NOT TS=(“lipoxygenase” OR “lactate oxidase” OR “liquid oxygen”)from 1995 to 2025. In the past 30 years, 2490 LOX - related publications in the cancer domain were unearthed. Among these, 2,041 (8%) were articles, and 449 (18%) were review articles, as shown in Figure 5A. The number of such publications has been steadily increasing. In the recent nine years, the annual output was approximately ten - fold that of 1996, as depicted in Figure 5B. These 2,041 documents originated from 78 countries and regions. The United States had the greatest number of publications (n = 704), followed by China (n = 519) and Japan (n = 175), together accounting for 68.5% of the total, as shown in Table 7. The 2027 analyzed documents were associated with 2,486 organizations. Among the top 20 productive academic institutions in Table 8, Shanghai Jiao Tong University (44), and Fudan University (42) were at the forefront. The University of California, San Francisco had the highest citation count, as manifested in the collaboration network presented in Figure 5C.
(A) The proportion of articles in the field of LOX associated with cancer research. (B) The number of publications in the field of LOX associated with cancer research by year. (C) Distribution of publications from different organizations in the field of LOX associated with cancer research. (D) Timeline view of keywords analysis in the field of LOX associated with cancer research. (E) The co-occurrence author map of co-authors in the field of LOX associated with cancer research. (F) The recurrence rate of cancer types. The thickness of the connecting lines between the nodes reflects the strength of the interaction. The color of the node represents the average year of publication for the country which the node represents.
Over the past decade, 13,219 authors published on LOX and cancer. Using VOSviewer, a network visualization was generated. Authors with four or more publications were selected for further analysis. In the cancer - related LOX study, 137 authors were involved. Philip C. Trackman had the most publications (25), followed by Janine T. Erler (22) and Thomas R. Cox (15). Their co - authorship map is presented in Figure 5D. Table 9 enumerates the top 20 co - cited references. The reference with the highest citation count was from a 2006 publication in Nature, authored by Janine T. Erler. This study delved into LOX’s role in tumor metastasis, especially its ECM - regulating mechanism for tumor invasion. Figure 5E’s keyword visualization displays nodes such as “activation”, “lysyl oxidase”, “lysyl oxidase - 2”, “gene”, “breast cancer”, “cancer”, and “extracellular matrix” connected by lines, highlighting the intricate relationships within LOX research. It clearly indicates that LOX research is closely intertwined with cancer, particularly breast cancer, the extracellular matrix, biological activation, and genetics.
Figure 5F is a bar chart illustrating the recurrence occurrences of various cancer types. The x - axis lists different cancer types, ranging from breast cancer to rhabdomyosarcoma. The y - axis represents the number of recurrences. Blue bars are used to depict the recurrence counts for each cancer. Breast cancer has the highest number of recurrences, reaching 69, while rhabdomyosarcoma has the lowest, with only 2 occurrences. A red dashed line is overlaid on the chart, which shows a downward trend, indicating that the recurrence numbers generally decrease from one cancer type to another. This chart effectively presents the differences and trends in recurrence frequencies among these cancers. This research on LOX encompasses the top ten most common tumors globally, with studies on lung cancer (15 documents), breast cancer (69 documents), colorectal cancer (32 documents), prostate cancer (19 documents), stomach cancer (33 documents), liver cancer (43 documents), cervical cancer (6 documents), thyroid cancer (2 documents), and bladder cancer (3 documents), but excludes research on esophageal cancer.
Discussion
In one of their other studies, it was revealed that LOX, secreted by hypoxic breast tumor cells, accumulates at pre - metastatic sites. It crosslinks collagen IV in the basement membrane, playing a pivotal role in the recruitment of CD11b+ myeloid cells (36). These CD11b+ cells bind to the crosslinked collagen IV and produce matrix metalloproteinase - 2, which cleaves collagen, thereby facilitating the invasion and recruitment of bone marrow - derived cells (BMDCs) and metastasizing tumor cells (46, 47).Regarding human colorectal carcinoma cell lines, the induction of LOX augmented HIF - 1 expression, whereas LOX silencing led to a reduction. Mechanistic investigations disclosed that LOX activated the PI3K (phosphoinositide 3 - kinase) - Akt signaling pathway, which, in turn, upregulated the synthesis of HIF - 1α protein in a manner contingent on LOX - mediated hydrogen peroxide production (48–50).
LOX proteins, especially LOXL2, are enzymes involved in the cross - linking of collagen fibers, thus playing a critical role in the stabilization of the extracellular matrix (ECM) (51–53). In cancer, dysregulation of these enzymes has been implicated in tumor progression and metastasis (42). The ECM serves as both a structural scaffold and a signaling hub, and its abnormal remodeling by LOX proteins contributes to the mechanical properties of tumors, facilitating invasion and metastasis. Emerging evidence suggests that LOX activity is regulated by various signaling pathways, including TGF - β and Wnt/β - catenin, which are frequently dysregulated in cancer (30, 54–56). Understanding the molecular mechanisms underlying LOX function could offer novel insights into tumor biology and therapeutic strategies (35, 57, 58).
The expression of LOX proteins has been shown to correlate with poor clinical outcomes in several cancers, such as breast, prostate, and ovarian cancers (59–61). This is largely attributable to their role in enhancing tumor stiffness, which promotes cancer cell survival, proliferation, and migration. Additionally, LOX enzymes may contribute to the maintenance of cancer stem cell properties by altering the ECM niche, thereby supporting tumor recurrence and resistance to therapy (28, 62, 63). Furthermore, LOX proteins are known to interact with other components of the tumor microenvironment, including immune cells, further complicating their role in cancer progression. These findings underscore the necessity for a comprehensive understanding of LOX - mediated ECM remodeling in cancer pathogenesis (64–67).
The outcomes of the mapping in Figure 4E suggest promising future research directions. These include delving into the interactions between LOX and the extracellular matrix during cancer metastasis and invasion, as well as exploring related gene regulatory pathways (68–72). Such research could potentially unlock novel approaches for diagnosing and treating diseases by targeting LOX (73–75). Inhibiting Lox enzymes has emerged as a promising therapeutic approach to combat cancer metastasis (76–81). Preclinical studies have demonstrated that targeting LOXL2 can reduce tumor stiffness, impair cancer cell invasion, and sensitize tumors to chemotherapy. Currently, several inhibitors of LOX enzymes are undergoing preclinical development and early - stage clinical trials (82, 83). These inhibitors aim to disrupt the cross - linking of collagen fibers, thereby reducing tumor stiffness and limiting cancer cell dissemination (84–90). Additionally, understanding the regulatory mechanisms of LOX expression could lead to novel therapeutic approaches, such as targeting upstream signaling pathways or combining LOX inhibition with immunotherapy to enhance anti - tumor immune responses (91–93). However, further research is needed to optimize these strategies and address potential off - target effects (94–96).
Lysyl oxidase (LOX) fulfills a critical function in vascular fibrosis and extracellular matrix (ECM) remodeling by catalyzing the oxidation of lysine residues in collagen and elastin, thereby facilitating ECM cross-linking and contributing to vascular wall stiffening (22). It orchestrates the phenotypic transition of vascular smooth muscle cells (VSMCs) from a contractile to a synthetic phenotype via the transforming growth factor-β (TGF-β)/Smad signaling cascade, which accelerates fibrotic processes and diminishes vascular compliance (97). LOX exacerbates vascular inflammation and endothelial dysfunction by activating the nuclear factor-κB (NF-κB) pathway, culminating in the secretion of pro-inflammatory cytokines, including interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α) (98). Additionally, LOX contributes to oxidative stress-induced damage to vascular endothelial cells, compromising endothelial barrier integrity and initiating atherosclerotic plaque formation (99). It enhances matrix metalloproteinases (MMPs) activity, leading to collagen degradation in the fibrous cap, which impacts plaque stability and the risk of acute cardiovascular events (100). LOX may also polarize macrophages toward a pro-inflammatory M1 phenotype, augmenting lipid phagocytosis and foam cell formation, while suppressing polarization to the anti-inflammatory M2 phenotype, thereby exacerbating intra-plaque inflammation and lipid accumulation (101). LOX interacts with lipid metabolism by promoting LDL oxidation to form oxidized LDL (ox-LDL) and can inhibit ATP-binding cassette transporters, obstructing cholesterol reverse transport from foam cells to high-density lipoprotein (HDL) (102). Therapeutic interventions targeting LOX, such as β-aminopropionitrile (BAPN) and LOXL2-specific inhibitors like GSK2878163, have demonstrated efficacy in mitigating ECM cross-linking and decelerating atherosclerosis progression in animal models (20). Early studies in chickens and pigs exhibited variable effects of dietary interventions on LOX activity, whereas in rabbits with advanced atherosclerosis and ApoE-deficient mice on a high-fat diet, LOX activity was elevated, and BAPN treatment ameliorated atherosclerosis (103). LOX expression is also implicated in human plaque stability. Its involvement in collagen cross-linking and its regulation by factors such as hypoxia and cytokines highlight its pivotal role in plaque stability, necessitating further investigation into its contribution to plaque instability (7, 20, 104)
Fibrosis and cancer intersect through the lysyl oxidase (LOX) enzyme, both sharing a disrupted extracellular matrix (ECM) microenvironment. LOX acts as a pivotal mediator in this context. Current fibrosis research has emphasized a detailed analysis of LOX isoform expression, uncovering its overexpression in lung, liver, heart, skin fibrosis, and hypertension. Barry-Hamilton discovered that LOXL2 drives the pathologic microenvironment of cancer and fibrotic diseases in lung and liver, with elevated levels in diseased stroma. AB0023, an inhibitory antibody targeting LOXL2, effectively reduced disease markers in both cancer and fibrosis models (95). Inflammation-driven conditions such as hepatitis not only foster fibrosis but also contribute to carcinogenic mutations, with LOX sustaining a pro-inflammatory and ECM rigidifying niche, thereby perpetuating an “inflammation-fibrosis-cancer” continuum (105, 106). Conversely, in fibrosis linked to cancer, LOX and transforming growth factor-β (TGF-β)-activated myofibroblasts secrete growth factors like HGF and FGF, which promote tumor cell viability and angiogenesis LOX’s enzymatic role extends to myelofibrosis, influencing megakaryocyte differentiation and platelet formation, and is critical for PDGF signaling and cell proliferation (107, 108). In a GATA-1-deficient myelofibrosis model, LOX overexpression drives ECM buildup and marrow fibrosis, which BAPN can mitigate. Elevated LOX activity in the marrow significantly contributes to myelofibrosis, with human myeloproliferative neoplasms showing heightened expression of several LOX isoforms (109). In mesothelioma, the build-up of asbestos in the pleural cavity causes prolonged inflammation, which prompts the malignant transformation of mesothelial cells and leads to fibrosis (110, 111). Therapeutically, LOX inhibition could concurrently benefit fibrosis and cancer treatment, such as LOXL2 inhibitors reducing ECM stiffness in fibrotic tumors to improve drug efficacy. Antifibrotic approaches include non-selective LOX inhibitors like BAPN and selective LOXL2 inhibitors, as well as MSC therapy, which can decrease LOX expression in liver injury models (3). The relationship between LOX family and extracellular matrix remodeling among various diseases is graphically represented in Figure 6.
The role of the LOX family in extracellular matrix (ECM) remodeling. The LOX family proteins, dependent on vitamin C and copper ions for their activity, contain both a catalytic domain and a propeptide domain. They play a crucial role in normal ECM remodeling involving elastin and collagen under physiological conditions. Overexpression or increased activity of LOX proteins, however, results in abnormal ECM remodeling linked to pathologies such as atherosclerosis, organ fibrosis, and various cancers. The LOX family acts as a key regulator in the transition between fibrosis and cancer. When LOX is secreted outside the cell, the N - terminus of the protein is cleaved by BMP – 1, forming the LOX propeptide (LOX - PP) and the catalytic domain of LOX protein. The catalytic domain of LOX proteins can promote cancer progression, whereas the LOX-PP has been shown to suppress cancer growth in certain types of cancer. WANG, Z. (2025) https://BioRender.com/2pa62d9.
Conclusion
This bibliometric analysis of LOX research from 1995 to 2025 identified 9,261 related publications. Keyword analysis showed “cancer” and “fibrosis” as key hotspots. LOX’s diverse functions and its links to cancer and fibrosis. This study provides an overview of LOX research trends, hotspots, and collaborations, guiding future research on LOX in diseases and therapies.
Future perspectives
Through our analysis of original research on LOX, we found that breast cancer accounts for the largest number of LOX-related studies. Therefore, we conducted a further in-depth analysis of LOX research in breast cancer. The results showed that: in breast cancer, the expression of LOXL1 is significantly upregulated, while that of LOX, LOXL2, and LOXL3 remains unchanged. Conversely, LOXL4 shows significant downregulation. Among these, only elevated LOXL2 expression demonstrates a strong correlation with a shortened disease - free survival duration in breast cancer patients, highlighting its potential as a critical biomarker for prognosis (59). The promotion of breast cancer progression by LOX family molecules is primarily mediated through several key mechanisms. Firstly, collagen cross - linking occurs, which contributes to the alteration of the extracellular matrix microenvironment (112). Secondly, the induction of epithelial - mesenchymal transition (EMT) is achieved by downregulating the expression of the E - cadherin epithelial marker and simultaneously upregulating the transcription of TWIST, thus facilitating the acquisition of a mesenchymal phenotype by cancer cells (113). Thirdly, the activation of the FAK/Src signalling pathway plays a pivotal role, triggering a cascade of intracellular events that enhance cell motility and invasiveness (28). Finally, the establishment of a premetastatic niche involving bone marrow-derived cells (BMDCs) is accomplished via collagen restructuring, providing a supportive microenvironment for the dissemination and colonization of cancer cells (114). As we look to the future of breast cancer research and treatment, the exploration of LOX family proteins holds immense promise. Despite the significant strides made in breast cancer diagnosis and prognosis with medical advancements and biomarker discoveries, distant metastasis remains a formidable challenge to patient survival.
The emerging role of LOX family proteins in promoting breast cancer, from regulating initiation and spread to mediating intracellular EMT remodelling, positions them as pivotal players in the disease process. Their potential as biomarkers for early detection, disease staging, and evaluating chemotherapy effectiveness, along with their candidacy as targets for metastasis prevention, has already been recognized. The development of various inhibitors, though showing promise, also underscores the need for a more refined therapeutic approach considering their differential selectivity for LOX family enzymes.
Moving forward, novel drug therapies targeting LOX family proteins are likely to be at the forefront of personalized and precise treatment strategies for breast cancer patients. An especially exciting frontier lies in investigating the role of LOX family proteins in the tumour immune microenvironment. Unravelling the regulatory mechanisms of these proteins on the breast cancer immune microenvironment and exploring the synergistic effects with immunotherapy could open new vistas in treatment.
To fully harness the potential of LOX family proteins in breast cancer management, several key research areas demand attention. There is an urgent need to bridge the knowledge gap regarding the oncogenic effects of LOXL1 and LOXL3 in breast cancer through in - depth fundamental and clinical studies. Additionally, while downstream targets of LOX family proteins have been the focus of existing research, understanding their upstream gene regulation and degradation pathways is essential. The use of gene - edited cells and mouse models can facilitate such investigations.
Identifying effective biomarkers, clarifying substrate - binding mechanisms, and understanding the molecular intricacies of LOX family proteins in breast cancer are crucial steps. Determining the optimal time for intervention to maximize treatment efficacy is another promising avenue. Given the pre - clinical success of LOX - targeting drugs in breast cancer and their efficacy in other cancer types’ clinical trials, further research into miR - mediated mechanisms of LOX family member activity could illuminate the tumour microenvironment’s molecular landscape and lead to innovative clinical therapies.
Limitations
Despite the significant contributions of existing research on LOX family, several limitations should be noted. First, potential omissions may arise due to inappropriate synonyms or abbreviations, as the keyword-based search strategy might have missed or mis-included relevant studies using alternative terminology introducing risks of incomplete retrieval due to semantic variability, particularly in fields with evolving nomenclature. Second, database-specific coverage limitations exist: although representative databases (e.g., PubMed, Web of Science) were systematically searched, other databases (e.g., CINAHL, BIOSIS Previews) and non-English/regional databases (e.g., CNKI) may have led to unintentional gaps, potentially missing valuable data on LOX in fibrosis or cancer. Third, temporal gaps in database updates constrain timeliness: even with a standardized search date, newly published studies—especially in rapidly advancing fields like precision oncology—may not yet be indexed, risking omission of breakthroughs in LOX-targeted therapies published shortly before the search date and affecting the review’s currency. Finally, the inability to capture post-submission retractions poses a challenge: studies retracted after manuscript submission could not be excluded, as retraction notices often lag behind original publications. While study integrity was verified at the time of review, unforeseen post-submission retractions introduce a minor risk of outdated or invalid data persisting, a common limitation in cross-sectional reviews that underscores the importance of ongoing critical evaluation in scientific discourse.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chen W Yang A Jia J Popov YV Schuppan D You H . Lysyl oxidase (LOX) family members: rationale and their potential as therapeutic targets for liver fibrosis. Hepatology. (2020) 72:729–41. doi: 10.1002/hep.31236 32176358 · doi ↗ · pubmed ↗
- 2Csiszar K . Lysyl oxidases: A novel multifunctional amine oxidase family. Prog Nucleic Acid Res Mol Biol. (2001) 70:1–32. doi: 10.1016/s 0079-6603(01)70012-8 11642359 · doi ↗ · pubmed ↗
- 3Trackman PC . Lysyl oxidase isoforms and potential therapeutic opportunities for fibrosis and cancer. Expert Opin Ther Targets. (2016) 20:935–45. doi: 10.1517/14728222.2016.1151003 PMC 498879726848785 · doi ↗ · pubmed ↗
- 4Stoyell-Conti FF Suresh Kumar M Zigmond ZM Rojas MG Santos Falcon N Martinez L . Gene inactivation of lysyl oxidase in smooth muscle cells reduces atherosclerosis burden and plaque calcification in hyperlipidemic mice. Atherosclerosis. (2024) 397:118582. doi: 10.1016/j.atherosclerosis.2024.118582 39260002 PMC 11465391 · doi ↗ · pubmed ↗
- 5Langton AK Tsoureli-Nikita E Griffiths CEM Katsambas A Antoniou C Stratigos A . Lysyl oxidase activity in human skin is increased by chronic ultraviolet radiation exposure and smoking. Br J Dermatol. (2017) 176:1376–8. doi: 10.1111/bjd.14959 27514484 · doi ↗ · pubmed ↗
- 6Kumarasamy A Schmitt I Nave AH Reiss I van der Horst I Dony E . Lysyl oxidase activity is dysregulated during impaired alveolarization of mouse and human lungs. Am J Respir Crit Care Med. (2009) 180:1239–52. doi: 10.1164/rccm.200902-0215 OC PMC 545114419797161 · doi ↗ · pubmed ↗
- 7Rodríguez C Martínez-González J Raposo B Alcudia JF Guadall A Badimon L . Regulation of lysyl oxidase in vascular cells: lysyl oxidase as a new player in cardiovascular diseases. Cardiovasc Res. (2008) 79:7–13. doi: 10.1093/cvr/cvn 102 18469024 · doi ↗ · pubmed ↗
- 8Kagan HM Li W . Lysyl oxidase: properties, specificity, and biological roles inside and outside of the cell. J Cell Biochem. (2003) 88:660–72. doi: 10.1002/jcb.10413 12577300 · doi ↗ · pubmed ↗