Virulence-Encoding Genes Conserved in Salmonella Isolated From Humans, Poultry, and Seafood
Yemisi Olukemi Adesiji, Vijaya Kumar Deekshit, Rasheed A. Odunola, Indrani Karunasagar, Oluwafemi B. Daodu, Al-Mustapha Ahmad

TL;DR
This study examines the presence of virulence genes in Salmonella from humans, poultry, and seafood in Nigeria and India.
Contribution
The study identifies conserved and absent virulence genes in Salmonella isolates from different sources and regions.
Findings
The genes sseB, sseD, sseF, sseT, and invH were conserved in most isolates.
Two clam isolates lacked sseD and sseF genes.
The sseC and sseG genes were absent in all tested isolates.
Abstract
Diverse virulence genes encode for the Type III secretion system (T3SS) in bacteria. In Salmonella, these genes are located in the Salmonella pathogenicity Islands 1 and 2 (SPI-1 and SPI-2), and they facilitate bacterial invasion and replication within macrophages, contributing to the burden of nontyphoidal Salmonella infections. In this study, we investigated the prevalence of selected virulence-encoding genes in 30 laboratory stocks of Salmonella enterica serovar Enteritidis from Nigeria (16 isolates) and nonclinical sources comprising poultry and seafood from India (14 isolates). Analysis of PCR amplicons revealed that the genes sseB, sseD, sseF, sseT, and invH were conserved in all the isolates except for two isolates obtained from clams, which did not have the sseD and sseF genes. In addition, the sseC and sseG genes were absent from all the tested isolates. This study provides…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
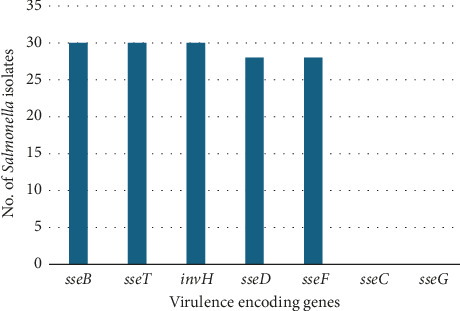 Figure 1
Figure 1- —Department of Science and Technology, Ministry of Science and Technology, India
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · Vibrio bacteria research studies · Escherichia coli research studies
1. Introduction
Salmonella serovars are among the major causes of food-borne infections worldwide, including developing countries [1]. They cause salmonellosis, which affects both humans and animals and is mostly caused by nontyphoidal Salmonella serovars [2]. Salmonellosis is a One Health challenge, as it poses a significant zoonotic threat to humans globally. The pathogenicity of Salmonella is mediated by virulence genes located on Salmonella pathogenicity islands (SPIs) as well as on plasmids and on the chromosome [3]. The 24 currently known SPIs, especially SPI-1 and SPI-2, play a vital role in virulence in Salmonella serovars and encode the Type III secretion system (T3SS). The T3SS is capable of secreting proteins called effectors, thereby facilitating pathogen invasion, survival, and replication within the host cells [2–4].
The SPI-1 system enables the invasion of epithelial cells, while the SPI-2 system facilitates the survival and replication of intracellular bacteria within Salmonella-containing vacuoles [5, 6]. The virulence-encoding genes in SPI-2 encode a T3SS that codes for proteins that form the Type III secretion apparatus, serve as transcriptional regulators, and function as effectors that invade and usually result in host cell destruction [4, 7].
The true burden of food-borne invasive and gastroenteric Salmonella infections in humans is difficult to evaluate in developing countries such as Nigeria and India due to a lack of robust epidemiological (genomic) surveillance systems. The invasive form of nontyphoidal Salmonella (iNTS) has been reported in many countries in sub-Saharan Africa, including Nigeria, using whole-genome sequence-based data [8]. This short communication is the result of our screening for seven virulence genes present in SPI-1 (one gene) and SPI-2 (six genes) from our laboratory stock of previously characterized Salmonella isolates. Evaluation of these virulence-encoding genes aimed to improve the current knowledge of the T3SS in resource-limited settings.
2. Materials and Methods
2.1. Ethical Approval
The ethical clearance for this study was obtained from the ethical review committee of the University Health Services, University of Ilorin, Kwara State, Nigeria, with reference number UIL/UHS/71.
2.2. Sample Processing and Confirmation
Laboratory stocks of Salmonella spp. obtained from poultry (meat and fecal samples), seafood (clams), and human clinical sources were resuscitated using the ISO 6579-1 guidelines of 2017 [9] (Table 1). Briefly, the isolates were directly inoculated into Rappaport–Vassiliadis (RV) broth (Merck, Germany) and incubated for 18 ± 2 h at 37°C. All the overnight cultures were later plated on HiChrome-improved Salmonella agar (HiMedia, Laboratories Pvt. Ltd, India). Three biochemical tests were performed to characterize the Salmonella isolates: the urease test (urease medium), Simmons citrate test (Simmons citrate agar), and indole test (Kovac's reagent), following the manufacturer's instructions [10].
2.3. Molecular Characterization
DNA was extracted using the boil and spin method, a protocol described by Ahmed and Dablool in 2017 [11]. Salmonella isolates were genotypically confirmed by PCR using the housekeeping gene invA. This gene was selected because it is the gold-standard PCR target for the confirmation of Salmonella. The cycling conditions and primer sequences for the invA gene were as described by Jayaweera et al. [12].
2.4. Virulence Gene Detection
The extracted DNA of PCR-confirmed Salmonella isolates was then used for duplex PCR amplification to detect the presence of specific genetic elements. The genes targeted were ssaT, sseB, sseF, sseG, sseD, and sseC of SPI-2 and invH of SPI-1, as previously described [13]. The primer sequences, specific annealing temperature, and product size for each PCR reaction are provided in Table 2. The PCR reactions were carried out in a total volume of 30 μL, comprising 3 μL of 10x PCR buffer, 3 μL of 2 mM dNTPs, 1.5 μL of 10 μM primers (each primer), 0.5 μL of 5 U/μL Taq DNA polymerase, and 2 μL of 100 ng/μL DNA template. The reaction mixture was made up to 30 μL with the addition of 15.5 μL of water. The reaction mixture was then subjected to thermal cycling to amplify the target DNA sequence using the following amplification parameters: initial denaturation for 5 min at 95°C; 30 cycles of amplification consisting of denaturation for 15 s at 95°C, annealing for 30 s at 51°C and extension for 30 s at 72°C; and final extension for 7 min at 72°C. The PCR products were then separated on a 1.5% agarose gel (Fisher BioReagents, Geel, Belgium) and visualized using ethidium bromide (MP Biomedicals, Ohio, USA) staining on a gel documentation system (Bio-Rad, USA). Salmonella enterica serovar Enteritidis ATCC13076 was included in all sets of reactions as a positive control to validate the results.
3. Result
3.1. Prevalence of Virulence-Encoding Genes
The 16 human clinical Salmonella isolates and the 14 nonhuman Salmonella isolates were confirmed as Salmonella by PCR amplification of the invA gene, which generated amplicons of 284 bp. The PCR for the virulence-encoding genes revealed that sseT, sseB (belonging to SPI-2), and invH (belonging to SPI-1) were conserved in all 30 Salmonella isolates. In addition, the detection rates for sseD and sseF were 93.3% (n = 28/30) for both genes. Notably, the sseC and the sseG genes were not detected in any of the tested isolates (Figure 1). The characteristics of the isolates included in this study are provided in Table 2.
4. Discussion
This study tested for the presence of selected virulence genes in Salmonella isolates. Our findings revealed that sseB, sseT, sseD, sseF, and invH were present in all the isolates. This could be because genes-encoding putative effector proteins of the T3SS of SPI-2 are required for bacterial invasion, virulence, and multiplication within macrophages [5]. Two clam isolates did not harbor the sseD and sseF genes. Many virulence genes can contribute to Salmonella pathogenicity, and the absence of one particular gene (as seen in the two clam isolates) could be compensated by the activities of other genes because of the redundancy known for the Salmonella genome. The transcriptional operon contains the cluster of most genes encoding components and effectors of the SPI-2 T3SS [6]. Moreover, sseA is the only reported chaperone for the SPI-2-encoded secretion apparatus proteins sseB and sseD [15]. No chaperones have been reported for any of the other SPI-2 effector genes, although sseA and sseB have been proposed to act as chaperones based on the sequence analysis [7]. Additionally, this study revealed that sseD and sseF genes were present in Salmonella isolates from humans and poultry but absent from seafood, suggesting that other virulence factors may contribute to the pathogenicity of this isolate in these animal species [15, 16].
In addition, sseF is one of the several genes required to confer bacteria survival by altering the trafficking of the Salmonella-containing vacuoles for SPI-2 [17]. Although not found in any of the isolates, the sseG gene was reported to be responsible for targeting Salmonella to the host Golgi network through poorly understood mechanisms [18]. In addition, studies have reported that sseG is not required for the translocation of effector proteins of SPI-2 [19, 20]. These observations indicated that sseF and sseG do not act as translocators for further substrate proteins in vivo and that their function is distinct from those of sseB, sseC, and sseD. Prior to this, it was reported that the effector proteins sseC and sseD predominantly localized in the bacterial cell membrane with the help of a functional SPI-2 T3SS [21].
The invH gene, which plays a role in SPI-1-mediated effector gene translocation, was present in all Salmonella isolates. It has previously been demonstrated that deletion of the invH gene in S. enterica serovar Typhimurium reduces its invasion efficiency by 70%–80% in vitro [22]. The protein encoded by the invH gene has been described to disrupt cytoskeletal and bacterial cell barriers, resulting in Salmonella invasion of the host gastrointestinal epithelium [23]. However, S. enterica serovar Senftenberg has been reported to elicit enteropathogenic infections in humans, despite the absence of many genes in the SPI-1 locus [24].
The genetic diversity of virulence-encoding genes in Salmonella requires further research to fully elucidate the functions and interactions between effector proteins (sseA–sseG) and other molecular markers [25–27]. In addition, these virulence-encoding genes were also harbored by enteropathogenic Salmonella spp. in poultry. Hence, routine hygiene measures include high-temperature cooking of poultry and clams and reducing the cross-contamination of ready-to-eat food. This study underscores the occurrence of Salmonella harboring virulence genes in humans, poultry, and seafood and highlights the need for One Health integrated surveillance. In addition, qualitative testing of food products is needed to guarantee their wholesomeness. The major limitations of this study are the small sample size and the fact that only a selection of genes in the T3SS was screened.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Majowicz S. E. Musto J. Scallan E. The Global Burden of Nontyphoidal Salmonella Gastroenteritis Clinical Infectious Diseases 201050688288910.1086/6507332-s 2.0-7774928289820158401 · doi ↗ · pubmed ↗
- 2Lerminiaux N. A. Mac Kenzie K. D. Cameron A. D. Salmonella Pathogenicity Island 1 (SPI-1): the Evolution and Stabilization of a Core Genomic Type Three Secretion System Microorganisms 202084 p. 57610.3390/microorganisms 8040576 PMC 723229732316180 · doi ↗ · pubmed ↗
- 3Guo C. Liu Y. Tang X. Liu X. Zhu J. Regulation Mechanisms of Salmonella Pathogenicity Island 1 and 2 Frontiers in Microbiology 202213 p. 84696410.3389/fmicb.2022.846964 · doi ↗
- 4Yin J. Li R. Zhang Y. Chen L. Zhu G. The Roles of the Salmonella Pathogenicity Islands in Bacterial Pathogenesis and Novel Targets for Therapeutic Intervention Frontiers in Cellular and Infection Microbiology 202111 p. 74018610.3389/fcimb.2021.740186 · doi ↗
- 5Jennings E. Thurston T. L. M. Holden D. W. Salmonella SPI-2 Type III Secretion System Effectors: Molecular Mechanisms and Physiological Consequences Cell Host & Microbe 201722221723110.1016/j.chom.2017.07.0092-s 2.0-8503054581428799907 · doi ↗ · pubmed ↗
- 6Lou L. Zhang P. Piao R. Wang Y. Salmonella Pathogenicity Island 1 (SPI-1) and its Complex Regulatory Network Frontiers in Cellular and Infection Microbiology 20199 p. 27010.3389/fcimb.2019.002702-s 2.0-85071582009 PMC 668996331428589 · doi ↗ · pubmed ↗
- 7Heithoff D. M. Shimp W. R. Lau P. W. Human Salmonella Clinical Isolates Distinct From Those of Animal Origin Applied and Environmental Microbiology 20087461757176610.1128/aem.02740-072-s 2.0-4084911981718245251 PMC 2268321 · doi ↗ · pubmed ↗
- 8Park S. E. Pham D. T. Pak G. D. The Genomic Epidemiology of Multi-Drug Resistant Invasive Non-typhoidal Salmonella in Selected Sub-saharan African Countries BMJ Global Health 202168 p. e 00565910.1136/bmjgh-2021-005659 PMC 833056534341020 · doi ↗ · pubmed ↗