MiR5651, miR170-3p, and miR171a-3p Regulate Cadmium Tolerance by Targeting MSH2 in Arabidopsis thaliana
Xianpeng Wang, Hetong Wang, Xiuru Sun, Zihan Tang, Zhouli Liu, Richard A. Ludlow, Min Zhang, Qijiang Cao, Wan Liu, Qiang Zhao

TL;DR
This study shows how three specific miRNAs improve Arabidopsis' ability to tolerate cadmium stress by targeting a DNA repair gene.
Contribution
The novel finding is that miR5651, miR170-3p, and miR171a-3p regulate cadmium tolerance by targeting AtMSH2 in Arabidopsis.
Findings
miR5651, miR170-3p, and miR171a-3p target AtMSH2 and their expression is negatively correlated with Cd tolerance.
Overexpression of these miRNAs enhances Cd stress tolerance in Arabidopsis.
Exogenous application of these miRNAs improves Cd tolerance in wild-type plants.
Abstract
The DNA mismatch repair (MMR) system plays a crucial role in repairing DNA damage and regulating cell cycle arrest induced by cadmium (Cd) stress. To elucidate the mechanism by which miRNAs target AtMSH2 in regulating Arabidopsis’ response to Cd stress, the wild-type Arabidopsis, Atmsh2 mutant, and three miRNA-overexpressing transgenic lines were grown hydroponically in half-strength MS solution containing cadmium (Cd) at concentrations of 0, 0.5, 1, 2, and 3 mg/L for 5 days. miRNA-seq analysis, bioinformatics prediction, dual-luciferase reporter assays, and qRT-PCR results demonstrated that miR5651, miR170-3p, and miR171a-3p specifically targeted AtMSH2 and their expression levels showed a significant negative correlation. Compared to wild-type (WT) Arabidopsis, Cd stress tolerance was significantly enhanced in miRNA-overexpressing transgenic lines. Moreover, exogenous application of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
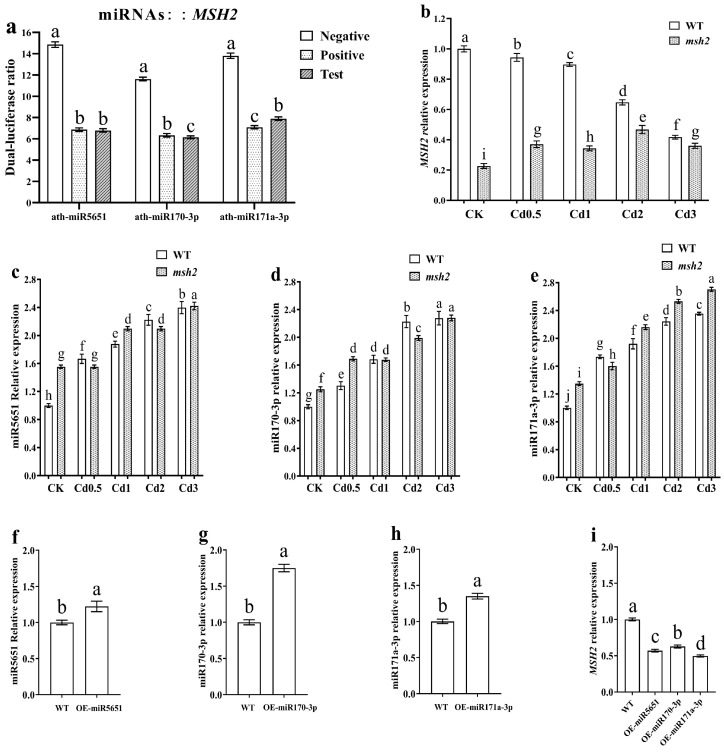 Figure 1
Figure 1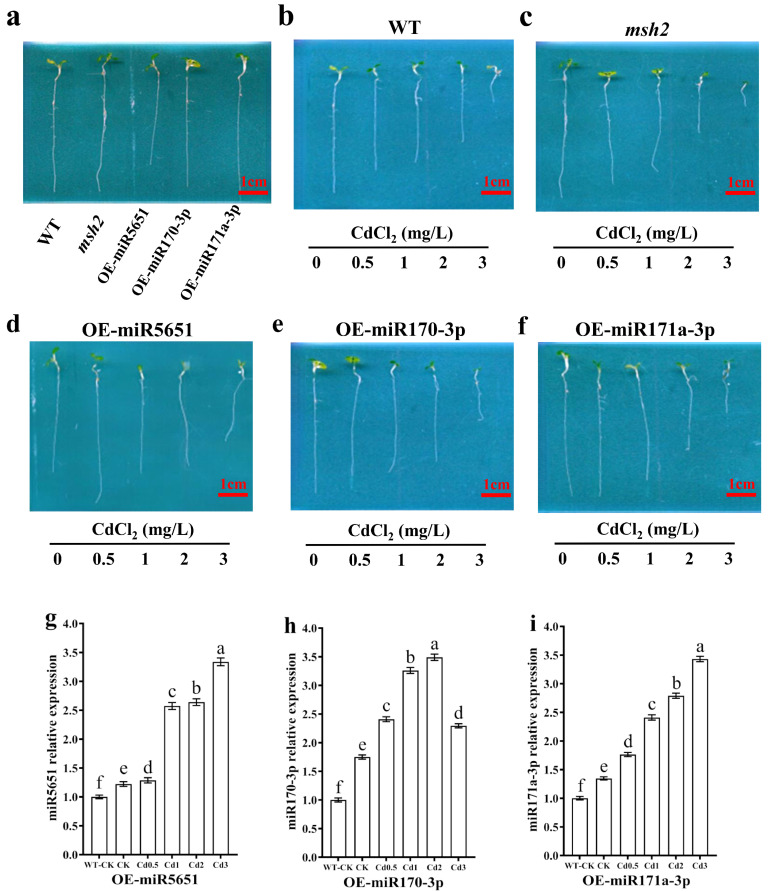 Figure 2
Figure 2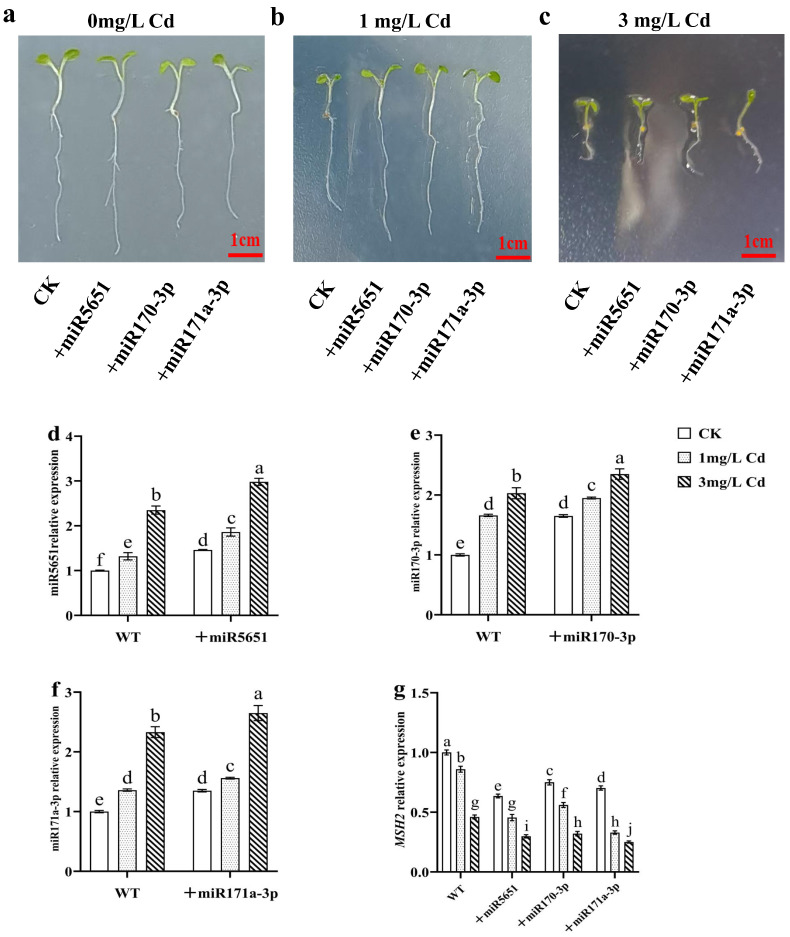 Figure 3
Figure 3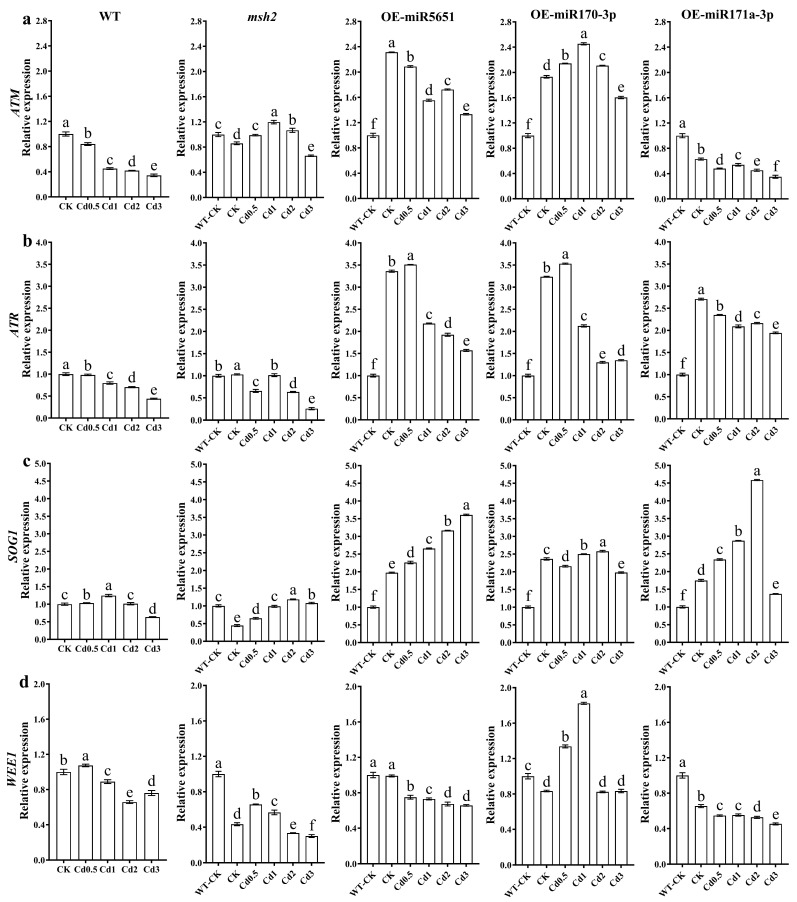 Figure 4
Figure 4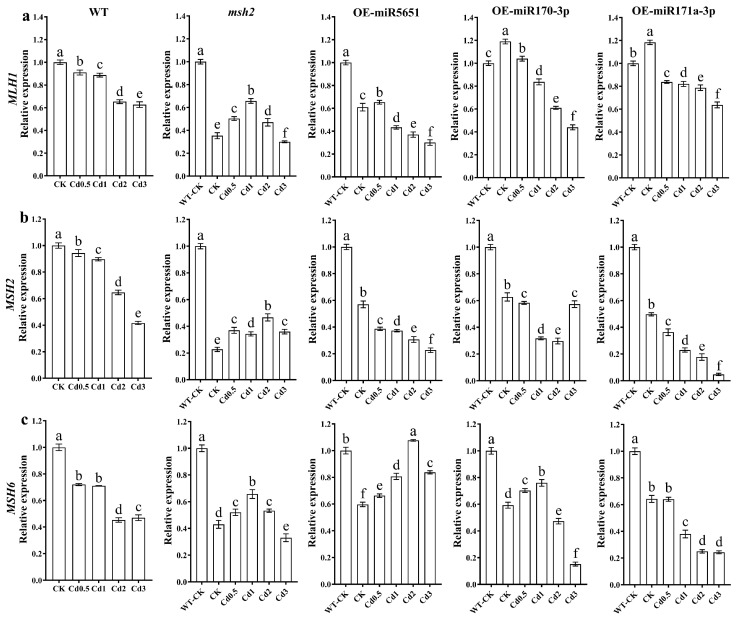 Figure 5
Figure 5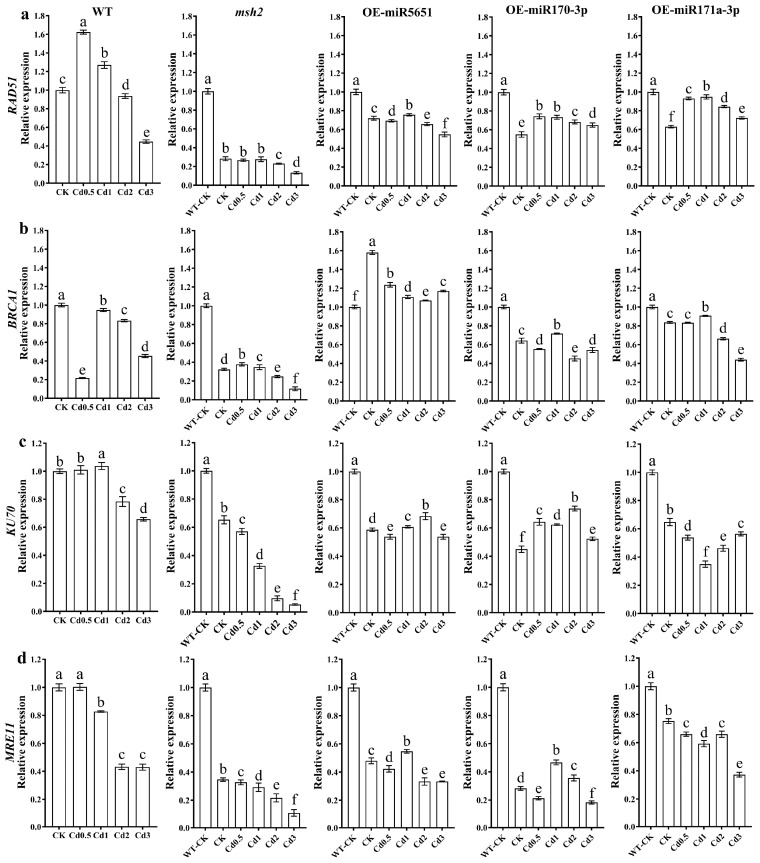 Figure 6
Figure 6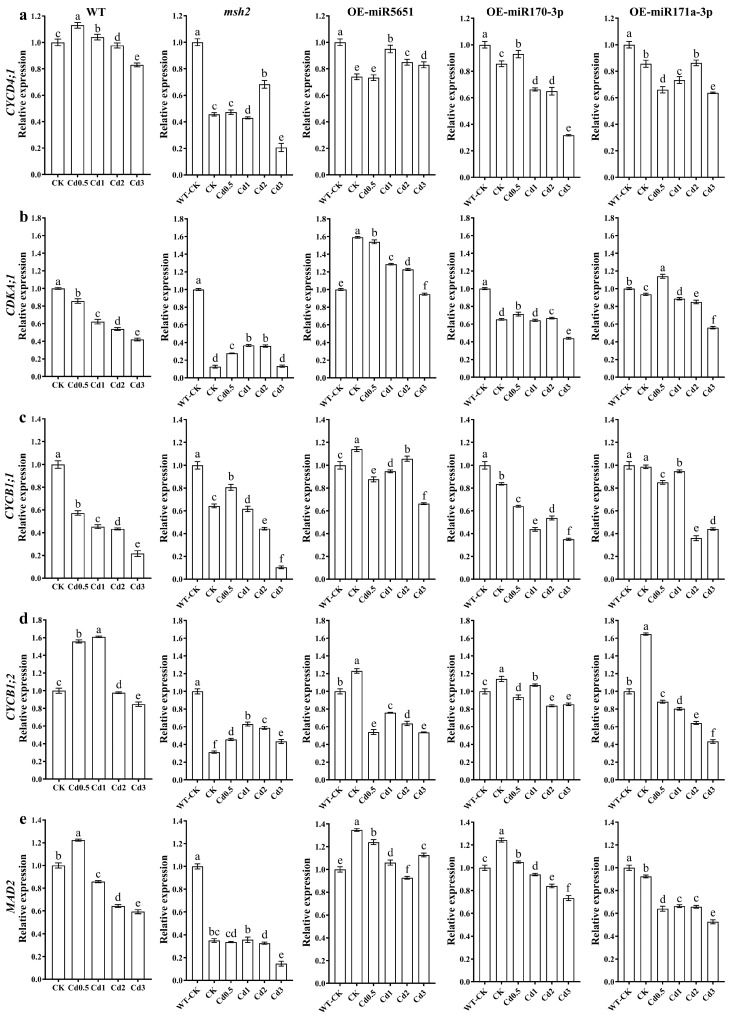 Figure 7
Figure 7- —National Natural Science Foundation of China
- —Liaoning Revitalization Talents Program
- —Science and Technology Plan Joint Project Natural Science Foundation-General Program of Liaoning Province
- —Northeast Geological S&T Innovation Center of China Geological Survey
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Stress Responses and Tolerance · Plant Molecular Biology Research · Plant Micronutrient Interactions and Effects
1. Introduction
Cadmium (Cd) pollution is a severe and urgent global environmental issue. Due to its wide range of sources and strong bioaccumulation characteristics, Cd has emerged as one of the most critical heavy metal pollutants in agricultural soil, which is ranked as the 4th most hazardous inorganic substance by the Agency for Toxic Substances and Disease Registry [1,2,3]. Among the limited number of effective remediation techniques for Cd-contaminated soils, phytoremediation—particularly based on phytoextraction using hyperaccumulators—has been extensively validated and widely recognized as an eco-friendly and sustainable approach [4,5,6]. Nonetheless, Cd is one of the most toxic heavy metals faced by plants. Although it is a non-essential element for plant growth and development, its high mobility in soils allows it to be easily absorbed by plant roots through cation and sulfate transporters and subsequently bioaccumulated in the human food chain, leading to severe health risks such as kidney dysfunction, osteoporosis, and cancers [7,8,9,10,11,12,13]. Exposure to Cd leads to multiple forms of DNA damage including base substitutions, base–base mismatches, insertion/deletion loops, DNA adducts, DNA breaks, DNA methylation, and DNA-strand cross-links [14,15]. In order to maintain genome integrity and prevent DNA damage transmission to daughter cells, the DNA mismatch repair (MMR) system is crucial for base–base mismatches, insertion/deletion loops, interstrand cross-links (ICLs), and double-strand breaks (DSBs), which triggers DNA damage response (DDR) to mediate cell cycle by activating ATR/ATM [16,17,18,19,20]. Thus, the efficacy of the DNA MMR system is considered as a proxy of Cd stress response, through regulating Cd-induced DNA damage sensitivity [21]. Moreover, MSH2, a key component of the MMR system, forms heterodimeric MutS complexes with MSH6, MSH3, and MSH7 in plants, which are responsible for the recognition of base–base mismatches, insertion/deletion loops, interstrand cross-links (ICLs), single-strand breaks (SSBs), and double-strand breaks (DSBs) with the replication protein A (RPA) complex and the MRE11-RAD50-NBS1 (MRN) complex [16,22,23].
MicroRNAs (miRNAs), 19–22 nt non-coding RNAs discovered in Caenorhabditis elegans and ubiquitously present in plants and animals, are processed from stem–loop regions of longer RNA transcripts to regulate gene expression at the post-transcriptional level by binding to specific sequences on target mRNAs, leading to mRNA cleavage or translational repression [24]. This mechanism enables miRNAs to critically modulate cellular processes such as differentiation, proliferation, and apoptosis [25,26]. Due to their indispensable regulatory role in living organisms, miRNAs have been considered as a molecular biomarker for various diseases and stress responses in plants [27,28,29,30,31,32]. In plants, upregulated or downregulated miRNAs induced by stress exert their critical physiological regulatory functions by either downregulating negative regulator target genes or upregulating positive regulator target genes during the stress response, thereby mitigating the toxic effects [33,34,35,36]. Due to recent progress in plant molecular biology, miRNAs have emerged as promising crop improvement players, given that miRNAs involve in plant vegetative growth, flowering, senescence, and fruit/grain setting [37,38,39]. Moreover, miRNAs have the potential as strategic tools for breeding stress-resilient crops. For example, miR393 and miR156 can improve salt and drought stress tolerance by targeting TIR1 and SPL genes, respectively, while miR166 and miR395 are capable of controlling plant Cd accumulation by targeting ABC transporters and sulfate transporters to mediate Cd transmembrane transport [40,41,42]. With regard to the epigenetic regulation of the DNA MMR system, miR-21, miR-137, and miR-155 have been found in human colorectal cancer targeting 3′ UTR regions of MSH2 or MSH6 mRNAs, thereby suppressing MMR function [43,44,45]. Although miRNAs mediating DNA MMR function have been reported, only limited information is available for miRNAs targeting MMR genes, especially in plants.
In this study, we employed bioinformatics prediction to preliminarily screen candidate miRNAs targeting the AtMSH2 gene. Subsequently, a tobacco (Nicotiana benthamiana L.) dual-luciferase reporter system was constructed to validate the targeting interaction between MSH2 and candidate miRNAs in vitro. To further confirm this regulatory relationship in vivo, transgenic Arabidopsis lines overexpressing the candidate miRNAs were generated. Finally, under Cd stress conditions, we analyzed the expression dynamics of the relevant miRNAs in wild-type (WT) Arabidopsis and miRNA-overexpressing lines, thereby elucidating miRNAs targeting AtMSH2 mediate Cd stress responses.
2. Materials and Methods
2.1. Materials, Growth, and Treatment Conditions
Arabidopsis thaliana (Columbia ecotype) and Atmsh2 T-DNA insertion mutant lines (SALK_002708, the background of the lines is from Col-0), and tobacco seeds were provided by the Soybean High Yield Cultivation Technology Innovation Team at the College of Agronomy, Heilongjiang Bayi Agricultural University, Daqing, China.
About 500 Arabidopsis seeds were placed in a 2 mL centrifuge tube and surface-sterilized using the 1 mL of hypochlorite (10% v/v) followed by 1 mL of ethanol (75% v/v) for 3 min, then washed with sterile distilled water 5 times. The seeds were immersed in 1 mL sterile water and vernalized at 4 °C for 24 h. The seeds were then sown in a culture bottle containing 150 mL of sterilized half-strength MS medium [46] (Basal Salt Mixture, Caisson, Colorado Springs, CO, USA) with 1.5% (w/v) sucrose (pH 5.8). After years of extensive toxicological screening, our research group determined that the Cd stress concentration range was 0–3 mg/L [18,21]. This range encompasses low-to-high stress levels suitable for investigating miRNA-mediated cadmium responses. Concentrations exceeding 3 mg/L caused elevated Arabidopsis mortality, compromising consistent sampling; whereas concentrations below 0.5 mg/L were insufficient to induce the stress required for toxicity/tolerance studies. For the Cd treatment, 0 (the control), 0.5, 1, 2, and 3 mg/L Cd^2+^ in the form of CdCl_2_·2H_2_O (analytical grade with purity 99.5%, China) were added into the half-strength MS medium [46] solution. Arabidopsis seeds were placed in a climate chamber at 21 ± 1 °C under 12 h light/12 h dark for 5 days.
For the exogenous miRNA treatments, 50 seeds of Arabidopsis were sown in six-well plates containing 2 mL of liquid MS medium with 0, 1, or 3 mg/L Cd treatment and 500 ng synthetic ds-miRNAs. The six-well plates were placed on a continuous shaker at 21 ± 1 °C and 12 h light/12 h dark photoperiod for 5 days. All treatments and analyses were repeated in three independent replicates.
2.2. Bioinformatics Prediction of miRNAs Targeting the AtMSH2
The CDs sequence information of the AtMSH2 gene was obtained from the NCBI database. The candidate miRNAs targeting the AtMSH2 were selected using psRNATarget (https://www.zhaolab.org/psRNATarget/, accessed on 7 August 2017) with the parameters set as Expectation (E) < 2. The Arabidopsis miRNA sequence information was retrieved from the miRBase database (http://www.mirbase.org/, accessed on 7 August 2017).
2.3. Plant Expression Vectors Construction and Dual-Luciferase Analysis
Plant expression vectors containing the precursor sequences of candidate miRNAs (miR5651, miR170-3p, and miR171a-3p) were, respectively, constructed using the pGreen_GUS_competitor plasmid (Addgene ID 55208) following the method described by Liu et al. [47] and designated OE-miR5651, OE-miR170-3p, and OE-miR171a-3p. The plant expression vectors of 3′UTR-AtMSH2 (containing the AtMSH2 fragment targeting the candidate miRNAs), 3′UTR-P (the positive control, completely complementary to the candidate miRNA target sequences), and 3′UTR-N (the negative control, completely non-complementary to the candidate miRNA target sequences) were constructed using the pGreen_3′UTR_sensor (Addgene ID 55206) plasmid, respectively. The primer information is shown in Table S1.
Following the method described by Liu et al. [47], Agrobacterium tumefaciens containing 3′UTR_sensor (3′UTR-AtMSH2, 3′UTR-P, and 3′UTR-N) was simultaneously injected into tobacco plant leaves (with a leaf age of 6 weeks), respectively, with the A. tumefaciens containing GUS_sensor (OE-miR5651, OE-miR170-3p, and OE-miR171a-3p). The tobacco plants were cultivated in a climate chamber at 27 ± 1 °C and a 16 h light/8 h dark photoperiod. After 3 days of growth, the tobacco leaves injected with A. tumefaciens were harvested. The fluorescence intensities of firefly (Lampyridae) luciferase and Renilla reniformis luciferase in tobacco leaves were detected using the Dual-Luciferase Assay Kit reagents (Promega Corporation, Madison, WI, USA. catalog# E4550) on a full-wavelength multifunctional microplate reader (Thermo Fisher Scientific Inc, Waltham, MA, USA). The dual-luciferase analysis had three biological replicates. Each biological replicate had three technical replicates.
2.4. miRNA Overexpression Transgenic Arabidopsis Plants Construction
miRNA overexpression transgenic Arabidopsis plants (OE-miR5651, OE-miR170-3p, and OE-miR171a-3p) were transformed using the inflorescence infection method following the method of Cheng et al. [48]. The background of the transgenic Arabidopsis plants is Col-0. A. tumefaciens containing the GUS_sensor (OE-miR5651, OE-miR170-3p, and OE-miR171a-3p) was used. The transgenic Arabidopsis plants were screened with 20 mg/L glufosinate-ammonium. DNA was extracted from the plants for PCR detection of the Bar gene. Homozygous transgenic Arabidopsis plants were cultivated to the T3 generation for subsequent experiments.
2.5. RNA Extraction, First-Strand cDNA Synthesis, and qRT-PCR Analysis
The total RNA was extracted from 0.1 g fresh samples preserved at −80 °C using TransZol Plant (TRANS, Beijing, China) following the manufacturer’s instructions. The RNA concentration was detected using the ultramicrospectrophotometer NanoDrop 2000C (Thermo Fisher Scientific, Waltham, MA, USA). The first strand of cDNA was synthesized from 1 μg total RNA using the HiScript^®^ III RT SuperMix for qPCR (Vazyme Biotech, Nanjing, China) following the manufacturer’s instructions. The qRT-PCR analysis was performed using the ChamQ Universal SYBR qPCR Master Mix (Vazyme Biotech, Nanjing, China) on the ABI Step One™ + real-time PCR system (ABI, Waltham, MA, USA). The UBQ 10 gene was used as the reference gene for signal normalization. The primers used for qRT-PCR are listed in Table S1. Relative gene expression levels between different treatments were calculated using the calculation method 2^−∆∆CT^ [49]. The qRT-PCR experiments had three biological replicates. Each biological replicate had three technical replicates.
2.6. miRNAs Expression Analysis
The miRNAs were extracted from 0.1 g fresh samples preserved at −80 °C using the TaKaRa MiniBEST Plant RNA Extraction Kit (Takara Bio Inc, Kusatsu-shi, Japan). The miRNA first strand was synthesized using the miRNA 1st Strand cDNA Synthesis Kit (by stem–loop) (Vazyme Biotech, Nanjing, China). The miRNA expression level was detected using the miRNA Unimodal SYBR qPCR Master Mix fluorescence quantitative detection kit (Vazyme Biotech, Nanjing, China) on the ABI Step One™ + real-time PCR system (ABI, Waltham, MA, USA). The U6 gene was used as the reference gene for signal normalization. The primers used for miRNA expression analysis were listed in Table S1. Relative expression levels of miRNAs between different treatments were calculated using the calculation method 2^−∆∆CT^ [49]. The qRT-PCR experiments had three biological replicates. Each biological replicate had three technical replicates.
2.7. Statistical Analysis
All the experimental data were analyzed using SPSS (version 29.0) and reported as the mean ± SD (standard deviation) values. Different letters indicate statistically significant differences between treatments at p < 0.05 by one-way ANOVA with Tukey’s test. The figures were produced using GraphPad Prism10.
3. Results
3.1. Bioinformatics Prediction Results of miRNAs Targeting the AtMSH2
To obtain AtMSH2 targeting miRNAs, the CDs sequence of the AtMSH2 gene was searched using the NCBI database and analyzed using the psRNA Target website. Expectation (E) < 2 was set to achieve a higher credibility. cDNA library selection: “Arabidopsis thalian, transcript, removed miRNA gene, TAIR, version 10” “released on 2010_12_14” miRNAs targeting the AtMSH2 gene were screened, and the matching results with the AtMSH2 gene are shown in Table 1. Three candidate miRNAs were obtained through bioinformatics analysis techniques, namely miR5651, miR170-3p, and miR171a-3p.
3.2. Determination of the Targeting Relationship Between miRNAs and AtMSH2
Dual-luciferin analysis was used to verify the targeting relationship between miR5651, miR170-3p, miR171a-3p, and AtMSH2. Compared with the negative control group, the relative luciferase activity in the miR5651, miR170-3p, and miR171a-3p test groups was significantly decreased, and showed no significant difference with the positive control group (Figure 1a). The qRT-PCR results demonstrated that Cd stress significantly downregulated the expression level of the AtMSH2 gene in WT, while upregulated the expression levels of miR5651, miR170-3p, and miR171a-3p when compared with the control (Figure 1b–e). Furthermore, these effects exhibited a dose-dependent relationship with the Cd concentration. In Atmsh2 mutants, Cd stress dose-dependently upregulated the expression of AtMSH2, miR5651, miR170-3p, and miR171a-3p.
In the miR5651, miR170-3p, and miR171a-3p overexpression transgenic Arabidopsis plants, the expression levels of miR5651, miR170-3p, and miR171a-3p were significantly upregulated, respectively. At the same time, the expression levels of AtMSH2 in those miRNA-overexpressing transgenic plants were significantly decreased (Figure 1f–i). Taken together, miR5651, miR170-3p, and miR171a-3p can target and negatively regulate the expression of AtMSH2.
3.3. The Effect of miRNAs Targeting and Regulating AtMSH2 on Plant Growth Under Cd Stress
To explore the effect of miRNAs targeting and regulating AtMSH2 on the Arabidopsis plants growth under Cd stress, the WT, Atmsh2, OE-miR5651, OE-miR170-3p and OE-miR171a-3p plants were subjected to different Cd concentration stress for 5 days. As showed in Figure 2a, there was no significant difference on the plant growth between the WT, Atmsh2, OE-miR170-3p, and OE-miR171a-3p under the normal condition, with an exception that OE-miR5651 plant roots are shorter than those lines. Compared with their respective control (0 Cd treatment), 0.5 mg/L Cd treatment could significantly inhibit WT and Atmsh2 plant roots growth, and the inhibition gradually increased with the increase in Cd concentration (Figure 2b–f). However, 0.5 mg/L Cd treatment had no significant effect on the root growth of OE-miR170-3p and OE-miR171a-3p plants when compared with their respective control. In contrast, 0.5 mg/L Cd treatment could promote root growth in OE-miR5651 plants compared to the control. In addition, when the Cd concentration reached 1 mg/L, the degree of root growth inhibition increased with the increase in Cd concentration.
qRT-PCR results showed that the expression levels of miR5651, miR170-3p, and miR171a-3p in the OE-miR5651, OE-miR170-3p, and OE-miR171a-3p plants were upregulated by the Cd stresses with a dose-dependent relationship to the Cd concentration with an exception that the expression level of miR170-3p in the OE-miR170-3p plant roots under the Cd3 treatment was lower than that in the Cd1 and Cd2 treatment (Figure 2g–i).
As shown in Figure 3, exogenous miRNAs treatments had no significant effect on the plant growth of WT under normal growth conditions. As expected, exogenous miR5651, miR170-3p, and miR171a-3p treatments improved the Cd tolerance of WT plants. In addition, exogenous miRNAs treatments upregulated the expression levels of miR5651, miR170-3p, and miR171a-3p, and downregulated the expression levels of AtMSH2 genes in the WT plant roots under the Cd stress, respectively.
3.4. The Effect of miRNAs Targeting and Regulating AtMSH2 on DNA Damage Response Signal Transduction Under Cd Stress
To explore the effect of miR5651, miR170-3p, and miR171a-3p targeting and regulating AtMSH2 on the DNA damage signal transduction in Arabidopsis plants under Cd stress, the relative expression levels of AtATM, AtATR, AtSOG1, and AtWEE1 genes in WT, Atmsh2, OE-miR5651, OE-miR170-3p, and OE-miR171a-3p plant roots under Cd stress were determined by qRT-PCR. As shown in Figure 4, compared with the control (WT-CK), Cd stress dose-dependently downregulated the expression levels of AtATM and AtATR in the WT, Atmsh2, OE-miR5651, OE-miR170-3p, and OE-miR171a-3p plant roots, with the exception that 0.5–2 mg/L Cd treatment upregulated the expression levels of AtATM in the OE-miR170-3p. It is worth noting that the expression levels of AtATM and AtATR in OE-miR5651, OE-miR170-3p, and OE-miR171a-3p plant roots were significantly higher than those of WT and Atmsh2 plant roots under the same Cd treatments. Furthermore, Cd treatments upregulated the expression levels of AtSOG1 in these miRNA overexpression plant roots when comparted with their corresponding control, and had higher expression levels than those of WT and Atmsh2. In contrast, the expression levels of AtWEE1 in OE-miR5651 and OE-miR171a-3p plant roots under Cd treatments were lower than those of WT, but higher than those of Atmsh2.
3.5. The Effect of miRNAs Targeting and Regulating AtMSH2 on DNA Mismatch Damage Repair Under Cd Stress
To explore the effect of miR5651, miR170-3p, and miR171a-3p targeting and regulating AtMSH2 on the DNA damage mismatch damage repair in Arabidopsis plants under Cd stress, the relative expression levels of AtMLH1, AtMSH2, and AtMSH6 genes in WT, Atmsh2, OE-miR5651, OE-miR170-3p, and OE-miR171a-3p plant roots under Cd stress were determined by qRT-PCR. As shown in Figure 5, compared with the control group (WT-CK), Cd stress dose-dependently downregulated the relative expression levels of AtMLH1, AtMSH2, and AtMSH6 in WT plant roots. Compared with CK, 0.5–2 mg/L Cd treatment significantly upregulated the relative expression levels of AtMLH1, AtMSH2, and AtMSH6 in Atmsh2 plant roots, but 3 mg/L Cd treatment significantly downregulated the expression levels of these genes. Compared with CK, 0.5–3 mg/L Cd treatment significantly downregulated the relative expression levels of AtMLH1 and AtMSH2 in the root systems of OE-miR5651, OE-miR170-3p, and OE-miR171a-3p plants, with an exception that 0.5 mg/L Cd treatment significantly upregulated the expression level of AtMLH1 in OE-miR5651. Compared with CK, Cd stress upregulated the expression level of the AtMSH6 gene in OE-miR5651. In OE-miR170-3p, the expression level of the AtMSH6 gene was upregulated by 0.5–1 mg/L Cd treatment and downregulated by 1–2 mg/L Cd treatment. In OE-miR171a-3p, the expression level of the AtMSH6 gene was significantly downregulated by 0.5–3 mg/L Cd treatment.
3.6. The Effect of miRNAs Targeting and Regulating AtMSH2 on DNA HR and NHEJ Under Cd Stress
To explore the effect of miR5651, miR170-3p, and miR171a-3p targeting and regulating AtMSH2 on the homologous recombination (HR) and non-homologous end joining (NHEJ) of DNA damage repair in Arabidopsis plants under Cd stress, the relative expression levels of AtRAD51, AtBRCA1, AtKU70, and AtMRE11 genes in WT, Atmsh2, OE-miR5651, OE-miR170-3p, and OE-miR171a-3p plant roots under Cd stress were determined by qRT-PCR. As shown in Figure 6, compared with the WT-CK, 2–3 mg/L Cd treatment significantly downregulated the expression levels of AtRAD51, AtBRCA1, AtKU70, and AtMRE11 genes in WT plant roots. However, 0.5–1 mg/L Cd treatment upregulated the expression level of AtRAD51 gene in WT, but downregulated the expression level of AtBRCA1 gene, and showed no significant effect on the expression level of AtKU70. The expression levels of AtRAD51, AtKU70, and AtMRE11 in OE-miR5651, OE-miR170-3p, and OE-miRNA171a-3p plant roots under 0.5–2 mg/L Cd treatments were significantly lower than those of WT, but higher than those of Atmsh2. Under Cd treatments, the expression level of AtBRCA1 in the roots of OE-miR5651 plants was the highest among all genotypes, while that in Atmsh2 roots was the lowest.
3.7. The Effect of miRNAs Targeting and Regulating AtMSH2 on Cell Cycle Regulation Under Cd Stress
To explore the effect of miR5651, miR170-3p, and miR171a-3p targeting and regulating AtMSH2 on the cell cycle regulation of Arabidopsis plants under Cd stress, the relative expression levels of AtCYCD4;1, AtCDKA;1, AtCYCB1;1, AtCYCB1;2, and AtMAD2 genes in WT, Atmsh2, OE-miR5651, OE-miR170-3p, and OE-miR171a-3p plant roots under Cd stress were determined by qRT-PCR. As shown in Figure 7, 1–3 mg/L Cd treatments downregulated the expression levels of AtCYCD4;1, AtCDKA;1, AtCYCB1;1, AtCYCB1;2, and AtMAD2 genes in the WT plant roots, with an exception that 1 mg/L Cd treatments upregulated the expression of AtCYCB1;2. Under Cd treatments, the expression level of AtCYCD4;1, AtCDKA;1, AtCYCB1;1, AtCYCB1;2, and AtMAD2 in the roots of OE-miR5651, OE-miR170-3p, and OE-miR171a-3p plants was higher than that in Atmsh2 roots. In addition, the expression level of AtCDKA;1, AtCYCB1;1, and AtMAD2 in the miRNA overexpression plant roots under Cd treatments was higher than that in the WT.
4. Discussion
MiR5651, miR170-3p, and miR171a-3p were found as underlying regulators to regulate of AtMSH2 based on bioinformatics prediction and our preliminary studies using miRNA sequencing. Building on our previous finding that MSH2 responds to Cd stress, we further observed Cd-responsive expression of these miRNAs. Moreover, overexpression of the above miRNAs promoted Cd tolerance of Arabidopsis. Both elucidating how these three miRNAs target the AtMSH2 gene to modulate DDR pathways and clarifying the mechanisms underlying the Cd resistance phenotype, constitute the primary objectives of this study.
4.1. miR5651, miR170-3p, and miR171a-3p Downregulate AtMSH2 Expression but Do Not Impair MMR-Mediated DDR
In this study, miR5651, miR170-3p, and miR171a-3p were first validated to target AtMSH2 by tobacco dual-luciferase reporter systems in vitro and Arabidopsis transgenic lines in vivo. In miRNA-overexpression transgenic lines, AtMSH2 expression was significantly suppressed by the above three miRNAs. However, the suppression level is lower than that observed in Atmsh2, with comparatively minor effects on MLH1 and MSH6 expression. Although the knockdown effect of miRNAs at the post-transcriptional level is inferior to T-DNA insertion mutants that knock out target genes at the DNA level, it is an effective approach to regulate the expression of target genes [50]. Nonetheless, multiple regulatory pathways, including DNA level, transcriptional level, post-transcriptional level, translational level, and post-translational level, exist within cells to control gene expression. As an epigenetic regulatory mechanism, miRNAs can only partially regulate gene expression and cannot fully govern its downstream functions and signaling pathways [51,52]. Therefore, overexpression of these miRNAs cannot substantially suppress AtMSH2 expression or significantly impair the associated MLH1 and MSH6 genes, thereby weakening the MMR-mediated DDR.
MMR-mediated DDR is crucial for Cd-induced DNA damage, which involves MSH2, MSH6, MSH3, and MSH7 that form MutS homolog complexes that recognize base–base mismatches, insertion/deletion loops and interstrand cross-links. When MMR-mediated DDR is activated by Cd-induced DNA damage, HR repair is recruited, and cell cycle arrest occurs until lesions are repaired [16]. ATM and ATR are the key protein kinases, activate thousands of transcriptional factors that respond to mismatches, SSBs, and DSBs induced by DDR. DNA lesions like mismatches and SSBs predominantly trigger ATR-dependent DDR, and activation of ATM is usually responsible for DSBs. In this study, compared with WT and Atmsh2, AtATR and AtATM expression was significantly promoted, and HR and NHEJ repair were not significantly suppressed. Furthermore, cell cycle arrest primarily occurred at the G_2_ phase, driven by upregulated mitotic checkpoint AtMAD2 and both stably expressed AtCYCD4;1 and AtCDKA;1 responsible for G_1_-S transition, which suggested that MMR-mediated DDR remained functional despite miRNA-mediated AtMSH2 downregulation. Phenotypic and gene expression analyses revealed that transgenic plants overexpressing candidate miRNAs (miR5651, miR170-3p, and miR171a-3p) exhibited sustained robust expression of AtRAD51 and AtBRCA1, whereas these DNA repair genes were significantly downregulated in Atmsh2 mutants. Collectively, our findings demonstrate that miR5651, miR170-3p, and miR171a-3p enhance Arabidopsis tolerance to Cd stress through fine-tuned modulation of stress-responsive pathways, rather than complete suppression of AtMSH2 expression.
4.2. miR5651, miR170-3p, and miR171a-3p Promote Cd Tolerance Due to Multiple DDR Engagement
According to the phenotype of overexpressed miRNA transgenic lines exposed to Cd and Cu stress, the above three miRNAs could promote Arabidopsis Cd tolerance compared with WT and Atmsh2. With the increasing gradient of Cd concentration, the root growth reduction in overexpressed miRNA seedlings was mitigated compared with WT seedlings. Heavy metal stress inevitably leads to reactive oxygen species (ROS) and damage to nucleic acids, proteins, and lipids. Cd stress primarily induces DNA damage, a critical cellular injury, which is one of the primary culprits responsible for growth inhibition. When DNA damage happens, cell cycle arrest will be triggered to maintaining genome stability and replication accuracy. G_1_/S and G_2_/M arrest are common responses to DNA lesions induced by heavy metals, whereas G_2_/M arrest supports plant growth potential. There are two convincing reasons to explain why G_2_/M arrest is better than G_1_/S for plants exposed to heavy metals stress. On the one hand, G_1_/S arrest will cause the comprehensively stationary state of cell reproduction. Based on this state, cell morphology remains unchanged, which suggests that plant cells will not enlarge. Furthermore, plant growth retardation is observed at the whole plant level, which is usually assumed to be stress sensitive [21]. On the other hand, the mitosis will activate until DNA damage is repaired. Although multiple DNA repair pathways exist from G_1_ to G_2_ phase, G_2_ phase is preferred by more error-free repair including nucleotide excision repair (NER), base excision repair (BER), MMR, and HR repair. Therefore, when cell cycle is arrest at G_2_/M phase due to DNA damage, DNA repair is efficiently driven through DDR, leading to the following mitosis after lesions repaired. Also, since the finished DNA replication and promoted synthesis of mRNAs and proteins, cell volume will increase for preparations of M phase. Thus, compared with at G_1_/S arrest, plants at G_2_/M phase are assumed to be more tolerant for stress.
In Atmsh2, AtMSH2 expression was significantly downregulated due to T-DNA insertion in the promoter region of the AtMSH2 gene, leading to the severe impairment of MMR function. MMR disorder further resulted in DDR switching, resulting in a transition from G_2_/M to G_1_/S arrest [18]. Therefore, stress intolerance was observed in Atmsh2 when exposed to Cd. In miRNA-overexpressed transgenic lines, the miRNAs partially suppressed AtMSH2 expression while retaining mismatch recognition, whereas MMR-mediated DDR maintained functional due to elevated AtATR expression and stably expressed AtRAD51 and AtBRCA1, which suggests functional MMR-mediated DDR [17]. However, both significantly expressed AtATR and AtATM indicated the activation of other multiple DDR, recruiting multiple DNA repair pathways. The underlying process can accelerate DNA damage repairing and finish DDR, leading to the mitosis. Thus, the engagement of multiple DDR promotes plant Cd tolerance in miRNAs-overexpressed transgenic lines because of limited knockdown effect on AtMSH2.
4.3. miR5651, miR170-3p, and miR171a-3p Are Capable to Induce Plant-to-Plant Cd Tolerance
Wild-type Arabidopsis seedlings acquired Cd tolerance after exogenous application of miR5651, miR170-3p, and miR171a-3p. This observation supports the role of these miRNAs in enhancing Cd stress tolerance and suggests that plants tolerant to Cd overexpressing these miRNAs may improve the resistance to Cd in neighboring plants via miRNA transfer between plants. In oncology, miRNAs serve as metastasis biomarkers and mediate distant cellular communication [52], whereas in plants, they act as signaling molecules that enable gene silencing across species and stress adaptation [24,53]. Although detailed mechanisms of miRNA entry into plant cells remain incompletely resolved, endocytosis and pinocytosis facilitate transmembrane transport, a process documented in animal studies where miRNAs originating from plants, including representative examples such as miR168a, traverse mammalian intestinal barriers via sequential transepithelial transport, ultimately regulating liver gene expression [54,55]. Similarly, miRNAs from plants, specifically rice (Oryza sativa L.) miR159a.1-1 and miR167a, enter insect epithelia, modulating PLCβ and RdRp expression [56]. The exogenous miRNA application experiment was conducted as a supplementary investigation to our prior mechanistic research, assessing the phytoremediation potential of this approach. Given inherent efficiency limitations in exogenous miRNA transmission and greater complexity of regulatory processes in natural systems, we selected 0, 1, and 3 mg/L Cd^2+^ concentrations rather than a full gradient for validation. Intercellular transport is further evidenced by miRNA trafficking mediated by phloem or xylem through plasmodesmata [57,58,59,60,61], with grafting experiments confirming transfer of miR166a and miR395b from rootstock to scion to regulate sulfur metabolism in tomato (Solanum lycopersicum L.) [41,62,63,64].
Plant miRNAs exhibit remarkable stability extracellularly. For instance, miR2911 from honeysuckle (Lonicera japonica Thunb.) maintains antiviral activity even after boiling [65]. In this study, chemically synthesized exogenous miRNAs delivered without RNA-binding proteins effectively enhanced Cd tolerance in Arabidopsis. This demonstrates that plant miRNAs possess intrinsic signaling capacity independent of protective complexes. While this study used exogenous application, endogenous miRNA secretion via extracellular vesicles provides a natural pathway for communication between plants [66]. These insights suggest viable strategies for phytoremediation and crop breeding: engineering plants that secrete miRNAs could confer tolerance across entire fields to Cd, enhancing decontamination efficiency while reducing risks of transgene dispersal.
5. Conclusions
In this study, miR5651, miR170-3p, and miR171a-3p targeting AtMSH2 were validated using the dual-luciferase reporter systems in vitro, followed by transformation with miRNAs in vivo. The qRT-PCR revealed that these miRNAs exhibited dose-dependent upregulation under Cd stress. However, based on the plant growth under Cd stress, miRNA-overexpressed mutants displayed enhanced Cd tolerance. Furthermore, this observation was further supported by exogenous application of these miRNAs to wild-type Arabidopsis, suggesting the miRNAs transferring and mediating the nearby plants in a plant-to-plant manner. Mechanistically, overexpression of these miRNAs activated ATR- and ATM-dependent DDR, inducing G_2_/M arrest to allow error-free repair. Notably, AtRAD51 and AtBRCA1 expression remained stable, ensuring HR efficiency despite partial suppression of AtMSH2 function. The partial suppression of AtMSH2 preserved MMR function, while co-activation of multi-pathway DDR engagement, enhancing Cd tolerance. This study provides a novel principle for elucidating Cd tolerance and offers insights into Cd tolerance breeding. However, the current research remains at the laboratory-based theoretical exploration stage, and its practical efficacy in authentic Cd-contaminated soil and aquatic environments requires validation. Further development of this study will accelerate the practical application of Cd phytoremediation technologies, while enhancing the efficiency of crop breeding under Cd stress and providing novel genetic resources and regulatory targets for Cd-tolerant germplasm innovation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agency for Toxic Substances and Disease Registry (ATSDR)Available online: www.atsdr.cdc.gov/(accessed on 13 April 2025)
- 2Xie Q. Deng W. Su Y. Ma L. Yang H. Yao F. Lin W. Transcriptome analysis reveals novel insights into the hyperaccumulator phytolacca acinosa Roxb. responses to cadmium stress Plants 20241329710.3390/plants 1302029738256850 PMC 10819451 · doi ↗ · pubmed ↗
- 3Liu H. Rong X. Zhao H. Xia R. Li M. Wang H. Cui H. Wang X. Zhou J. Bioaccumulation of atmospherically deposited Cadmium in soybean: Three-year field experiment combined with Cadmium isotopes Environ. Sci. Technol.202458177031771610.1021/acs.est.4c 0796139317642 · doi ↗ · pubmed ↗
- 4Niu L. Li C. Wang W. Zhang J. Scali M. Li W. Liu H. Tai F. Hu X. Wu X. Cadmium tolerance and hyperaccumulation in plants—A proteomic perspective of phytoremediation Ecotoxicol. Environ. Saf.202325611488210.1016/j.ecoenv.2023.11488237037105 · doi ↗ · pubmed ↗
- 5Raza A. Habib M. Kakavand S. Zahid Z. Zahra N. Sharif R. Hasanuzzaman M. Phytoremediation of Cadmium: Physiological, biochemical, and molecular mechanisms Biology 2020917710.3390/biology 907017732708065 PMC 7407403 · doi ↗ · pubmed ↗
- 6Hussain M. Kaousar R. Ali S. Shan C. Wang G. Wang S. Lan Y. Tryptophan seed treatment improves morphological, biochemical, and photosynthetic attributes of the sunflower under Cadmium stress Plants 20241323710.3390/plants 1302023738256789 PMC 10819145 · doi ↗ · pubmed ↗
- 7Zhang Z. Wang S. Wang J. Zhang C. Liu D. Wang C. Xu F. The overexpression of LOW PHOSPHATE ROOT 1 (LPR 1) negatively regulates Arabidopsis growth in response to Cadmium (Cd) stress Plant Physiol. Biochem.202319655656610.1016/j.plaphy.2023.02.00336774911 · doi ↗ · pubmed ↗
- 8Babar H. Muhammad Nadeem A. Shafeeq-ur-Rahman Aqleem A. Jumei L. Muhammad F. Cadmium stress in paddy fields: Effects of soil conditions and remediation strategies Sci. Total Environ.20217541421883325494210.1016/j.scitotenv.2020.142188 · doi ↗ · pubmed ↗