Beyond Pairwise Interactions: How Other Species Regulate Competition Between Two Plants?
Wang-Xin Cheng, Wei Xue, Jie-Jie Jiao, Hao-Ming Yuan, Lin-Xuan He, Xiao-Mei Zhang, Tao Xu, Fei-Hai Yu

TL;DR
This study shows how the presence of additional plant species can influence competition between two plants, highlighting the importance of considering complex interactions in plant communities.
Contribution
The study introduces a novel approach to assess how third, fourth, and fifth plant species influence pairwise interspecific interactions.
Findings
The presence of neighbor plants generally reduced the growth of target species, indicating competitive interactions.
Other species can alter interactions between two target species, with effects depending on species identities.
The impact of the third species was largely independent of the fourth and fifth species.
Abstract
A plant species in a community often grows with some other plant species. While many studies have assessed interspecific interactions between two target plant species, few have considered the impacts of the other plant species (e.g., the third, fourth, and fifth plant species) on these interactions. To assess the impacts, we grew one seedling of each of the five herbaceous plant species that are common in China (Cynodon dactylon, Plantago asiatica, Taraxacum mongolicum, Nepeta cataria, and Leonurus japonicus) alone (no competition) or with one seedling of one, two, three, or four of the other species. The presence of a neighbor plant generally reduced the growth of the target species, suggesting that the interspecific relationships were mostly competitive. The presence of other neighbor species (the third, fourth, and fifth species) could alter the interspecific interactions between two…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
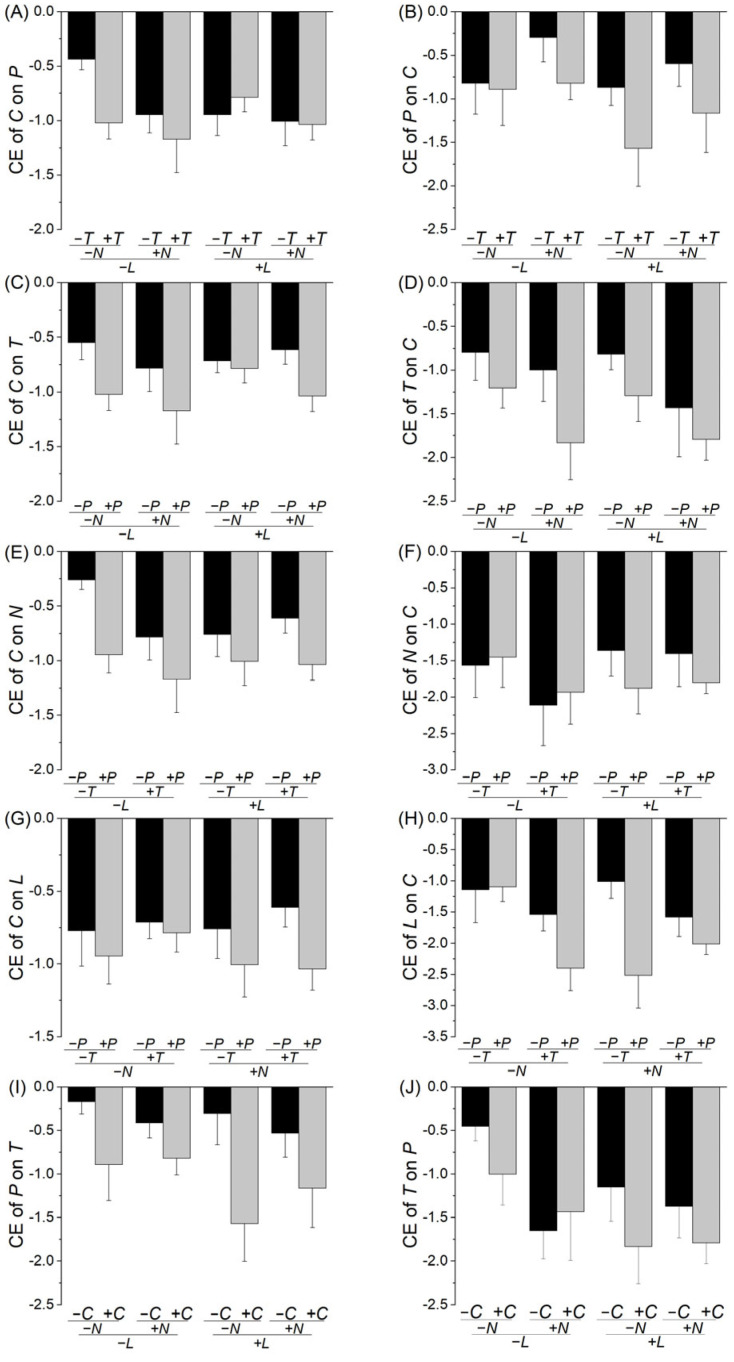 Figure 1
Figure 1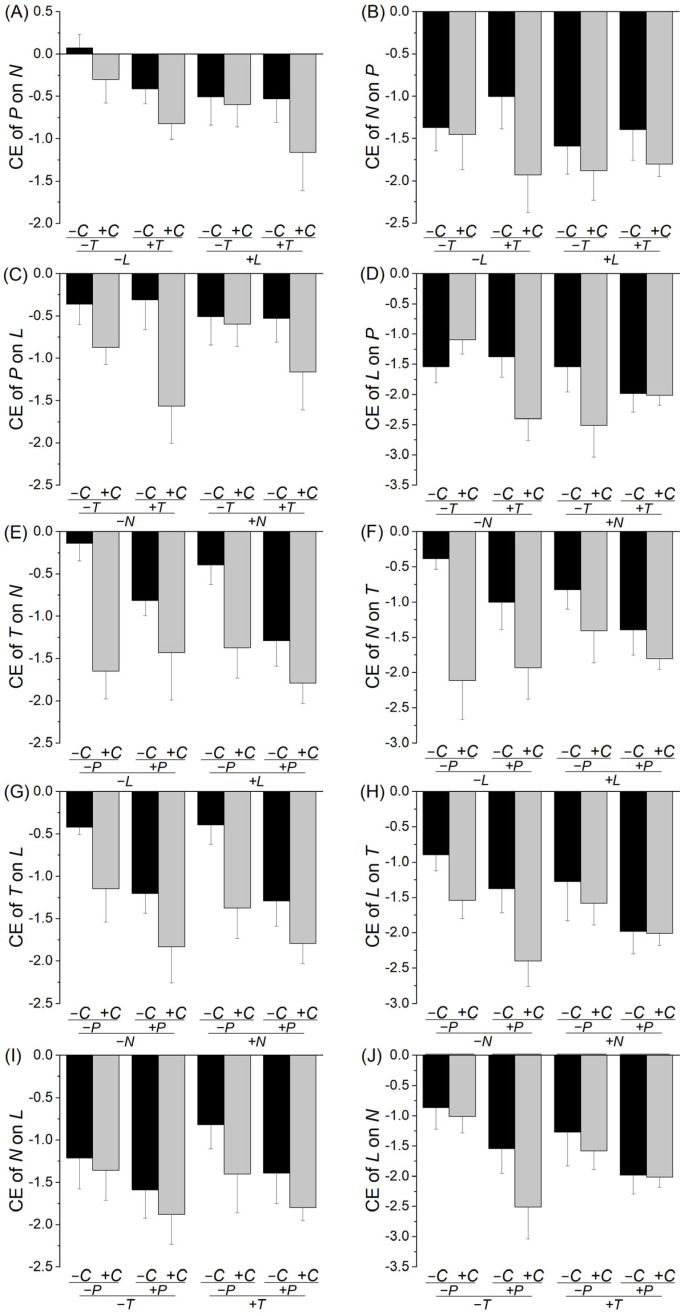 Figure 2
Figure 2- —National Key R&D Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEcology and Vegetation Dynamics Studies · Plant and animal studies · Plant Parasitism and Resistance
1. Introduction
In natural ecosystems, species are rarely found in isolation; they typically grow and interact with a variety of other species [1,2]. Interactions among plant species are important determinants of species coexistence and community structure, and ecosystem function [3,4,5]. Competition is one of the most common plant–plant interactions and plays a key role in natural ecosystems, influencing species survival and growth, resource acquisition, population dynamics, and ecosystem stability [6,7,8]. Understanding factors regulating plant competition is important for developing predictive models of species coexistence and community dynamics [9,10,11].
Competitive interactions between two target plant species can be regulated by various environmental factors, including temperature [12,13,14], CO_2_ concentration [15], O_3_ concentration [16], water [17,18], light [19], nutrients [20,21], salinity [22], heavy metal contamination [23], and individual density [24]. For example, in a maize–soybean cropping system, maize typically outcompetes soybean; however, the addition of nitrogen and phosphorus fertilizers can reduce this competitive advantage [25]. Invasive species often outcompete native species, reducing native species’ richness and abundance, but such a competitive effect can be modified by light, soil water, and nutrients [26,27,28].
As a plant species often coexists with more than one plant species, indirect interactions between two plant species can also be essential for species coexistence and biodiversity maintenance [29]. One example of this is that competitive interactions between two target plant species can be modified by organisms from other tropic levels [30]. For instance, Solanum altissima and Solidago carolinense can coexist when insect herbivores are not present, but S. altissima may outcompete S. carolinense when the herbivores are present [31]. Also, compared to the absence of arbuscular mycorrhizal fungi, the presence of the fungi can increase the competitive ability of Vicia faba on Hordeum vulgare by improving the nitrogen uptake of V. faba [32]. Moreover, endophyte-infected plants show a higher competitive ability than those that are not infected [33].
The introduction of new plant species into a community can further complicate competitive interactions and, thus, coexistence between two target species [34,35]. In a recent study, the duckweed Spirodela Polyrhiza was found to suppress another duckweed, Lemna minor, when Pistia stratiotes was not present, but such a suppressing effect diminished when P. stratiotes was included [36]. Studies have also shown that both the species richness of a plant community and genotypic richness of a clonal plant population could alter competitive interactions between two target plant species [37,38]. However, as the number of other plant species increases, interspecific interactions between two target plant species may become more complex. So far, however, it is still unknown as to whether the impact of the other plant species (e.g., the third plant species) on competitive interactions between two target plant species is common and whether the impact varies depending on the presence of more additional plant species (e.g., the fourth and the fifth plant species).
To assess the impacts of other plant species on competitive interactions between two target plant species, we grew one seedling of each of the five herbaceous plant species that are common in China (Cynodon dactylon, Plantago asiatica, Taraxacum mongolicum, Nepeta cataria, and Leonurus japonicus) alone (no competition) or with one seedling of one, two, three, or four of the other species. Specifically, we address (1) how common the impact of the third plant species is on competition between two target plant species, (2) whether the impact of the third plant species varies depending on the identity of the target species and the identity of the third species, and (3) whether the impact of the third plant species varies depending on the presence of the fourth and the fifth species.
2. Results
The presence of neighbor plants generally reduced the growth of the target species, as shown by the negative value of the log response ratio, suggesting that the interspecific relationships were mostly competitive (Figure 1 and Figure 2). The presence of the third species could alter the interspecific interactions between two target species (Table 1, Figure 1 and Figure 2). Specifically, the presence of P. asiatica aggravated (p < 0.05) or tended to aggravate (0.05 < p < 0.1) the competitive effect of C. dactylon on T. mongolicum (Table 1C, Figure 1C); T. mongolicum on C. dactylon (Table 1D, Figure 1D), N. cataria (Table 1O, Figure 2E) and L. japonicus (Table 1Q, Figure 2G); C. dactylon on N. cataria (Table 1E, Figure 1E) and L. japonicus (Table 1G, Figure 1G); L. japonicus on C. dactylon (Table 1H, Figure 1H), T. mongolicum (Table 1R, Figure 2H) and N. cataria (Table 1T, Figure 2J); and N. cataria on L. japonicus (Table 1S, Figure 2I). Similarly, the presence of C. dactylon aggravated (p < 0.05) or tended to aggravate (0.05 < p < 0.1) the competitive effect of P. asiatica on T. mongolicum (Table 1I, Figure 1I), N. cataria (Table 1K, Figure 2A), and L. japonicus (Table 1M, Figure 2C); N. cataria on P. asiatica (Table 1L, Figure 2B) and T. mongolicum (Table 1P, Figure 2F); T. mongolicum on N. cataria (Table 1O, Figure 2E) and L. japonicus (Table 1Q, Figure 2G); and L. japonicus on T. mongolicum (Table 1R, Figure 2H). However, the presence of N. cataria only tended to aggravate (0.05 < p < 0.1) the competitive effect of C. dactylon on P. asiatica (Table 1A, Figure 1A), and T. mongolicum on P. asiatica (Table 1J, Figure 1J); the presence of L. japonicus only aggravated (p < 0.05) the competitive effect of T. mongolicum on C. dactylon (Table 1D, Figure 1D); and the presence of T. mongolicum tended to aggravate (0.05 < p < 0.1) the competitive effect of P. asiatica on C. dactylon (Table 1B, Figure 1B) and N. cataria (Table 1K, Figure 2A), and L. japonicus on C. dactylon (Table 1H, Figure 1H).
We found a significant interaction effect of C. dactylon × T. mongolicum × N. cataria on the competitive effect of L. japonicus on P. asiatica (Table 1N): without N. cataria, the presence of C. dactylon weakened the competitive effect of L. japonicus on P. asiatica in the absence of T. mongolicum, but strengthened it in the presence of T. mongolicum; with N. cataria, however, the presence of C. dactylon aggravated the competitive effect of L. japonicus on P. asiatica in the absence of T. mongolicum, but had no impact in the presence of T. mongolicum (Figure 2D). There was also a marginally significant effect of P. asiatica × T. mongolicum × N. cataria on the competitive effect of species L. japonicus on C. dactylon (Table 1H): without N. cataria, the presence of P. asiatica had no impact on the competitive effect of L. japonicus on C. dactylon in the absence of T. mongolicum, but aggravated it in the presence of T. mongolicum; with N. cataria, the presence of P. asiatica aggravated the competitive effect of L. japonicus on C. dactylon, but such an impact was much weaker in the presence than in the absence of T. mongolicum (Figure 1H). However, we found no other significant two- or three-way interaction effects (Table 1).
3. Discussion
While it is well known that interspecific interactions between two plant species can be regulated by many biotic and abiotic factors [15,16,19,22,23], including the presence of species from other trophic levels [31,32] and the presence of other plant species [39,40], relatively few studies have explored how the presence of multiple additional plant species modifies interspecific plant–plant interactions [37]. Understanding these multi-species effects is crucial for predicting species coexistence and community stability in natural ecosystems. Our results suggest that it may be a common phenomenon that the presence of one additional plant species in a community can alter the competition between two target plant species, but it may be uncommon that this impact depends also on the presence of more plant species (i.e., a fourth and/or fifth plant species). These findings highlight the complexity of multi-species interactions within a plant community.
While the impact of the third plant species on competition was found to be relatively common, it varied depending on the identity of the target species. For instance, the presence of Plantago asiatica aggravated the competitive effect of Cynodon dactylon on Taraxacum mongolicum (Table 1C, Figure 1C), T. mongolicum on C. dactylon (Table 1D, Figure 1D), and C. dactylon on Nepeta cataria (Table 1E, Figure 1E), but did not alter the competitive effect of N. cataria on C. dactylon (Table 1F, Figure 1F). This variation may be due to the fact that different target plant species may differ greatly in factors such as their competitive abilities, resource utilization strategies, and potential allelopathic effects [41,42]. For example, C. dactylon uses a strategy of rapid stolon expansion and its root system allows it to rapidly occupy limitedly soil space and acquire soil resources [43], but N. cataria is capable of releasing allelopathic compounds that affect the growth of neighboring plants, altering the competitive balance [44]. Consequently, the impact of the third species on the competitive effect differed among target species, reflecting the complexity of interspecific relationships [45,46].
Additionally, the impact of the third plant species on plant–plant competition varied with the identity of the third plant species. For example, the competitive effect of C. dactylon on T. mongolicum was not influenced by the presence of N. cataria, Leonurus. japonicus, N. cataria, or L. japonicus, but it was aggravated by the presence of P. asiatica (Table 1C, Figure 1C). The rosette growth form of P. asiatica enables it to quickly occupy aboveground space and intercept light at low canopy levels, which was particularly influential during the early stage of community in this experiment. This trait likely reduced light availability for neighboring seedlings, thereby modifying the growth performance and interspecific interactions of co-occurring species. Again, this context-dependent impact of the third species is likely due to the differences in, for example, competitive abilities and resource acquisition strategies among the third plant species involved [41,42]. These results suggest that different third species may play functionally distinct roles in modifying competition, either amplifying or mitigating the strength of interactions depending on their traits. These findings align with those of previous studies that emphasize the role of plant functional traits (e.g., root [47] or leaf area [48]) in shaping competitive dynamics. For instance, plant size asymmetry has been shown to influence competition strength, with larger individuals often exerting stronger suppressive effects on smaller competitors [36]. Similarly, species with extensive root systems and high resource acquisition rates tend to increase competition intensity, whereas species with complementary resource-use strategies reduce direct competition [49,50]. This could explain why some third species in our study intensified competition, while others had little or no effect. Additionally, indirect interactions mediated by belowground processes, such as root exudates influencing microbial activity, may have further contributed to these species–specific differences in competitive effects [51].
We observed that the impact of P. asiatica on competition between L. japonicus and C. dactylon and the impact of C. dactylon on competition between L. japonicus and P. asiatica varied depending on the presence of both T. mongolicum and N. cataria (Table 1H,N, Figure 1H and Figure 2D). These findings suggest that the presence of a fourth and/or a fifth plant species can influence the impact of the third plant species on plant–plant interactions. However, we found no other significant two- or three-way interaction effects (Table 1), suggesting that these impacts are not so common. One explanation is that increasing species richness may directly reduce the strength of individual pairwise interactions due to resource partitioning or niche differentiation, leading to a more diffuse interaction network [52,53,54,55]. Another explanation lies in the stabilization of interaction networks [56]. Previous studies have shown that competitive rings tend to be more stable when the number of species involved is odd rather than even [57], suggesting that community-level patterns of interaction may be sensitive to species number and composition [58,59]. This complexity highlights the non-additive nature of species interactions, where the presence of more species does not simply accumulate effects but reshapes the entire interaction web.
One caveat is that the current experimental design (additive design) does not allow us to exclude the potential cofounding effect of changing density [60,61], and the lack of morphological, physiological. and allocational measurements precludes the possibility to explore the underlying mechanisms. Therefore, a novel experimental design and morphological, physiological, and allocational measurements are required to better understand the complex multi-species interactions in a plant community. The experiment for lasted more than three months in the greenhouse, which is more than the period of most potting experiments. Also, the pots used were small (10 cm × 10 cm × 10 cm) and plant roots and the above-ground parts had filled the pot. Thus, the experiment duration was long enough to address our questions.
4. Materials and Methods
4.1. Plant Species
To address the questions, we chose five plant species, i.e., Cynodon dactylon (L.) Persoon (Poaceae), Plantago asiatica L. (Plantaginaceae), Taraxacum mongolicum Hand.-Mazz. (Asteraceae), Nepeta cataria L. (Lamiaceae), and Leonurus japonicus Houtt. (Lamiaceae) (Table 2; Flora of China, www.iplant.cn (accessed on 30 May 2023)). They were chosen because they are all herbaceous plant species that are common in China and can co-occur in, for example, hillside grasslands, abandoned fields, and roadsides in eastern China where the experiment was conducted. C. dactylon is cable of clonal growth by rooting at the nodes of its creeping stems, while all of the other four species are non-clonal plants. C. dactylon, T. mongolicum, and N. cataria are perennial, P. asiatica is biennial or perennial, while L. japonicus is annual or biennial.
Seeds of these species were purchased online from the Ferryman’s Seed Superstore (Zhoukou, Henan Province, China). In June 2023, we sowed the seeds of the five species in trays (54 cm long × 28 cm wide × 5 cm deep) filled with peat and placed the trays in a greenhouse at Taizhou University in Taizhou, Zhejiang Province, China. After one month of germination and growth, for each species, more than 96 seedlings of a similar size were selected and used for the experiment described below.
4.2. Experimental Design
The competition experiment used an additive design. One seedling of each of the five species was grown alone (no competition) in a pot or with one seedling of one, two, three or four of the other species in a pot, resulting in a total of 31 treatments (species combinations; Table 3). Each treatment was replicated six times, resulting in a total of 186 pots.
On 24 July 2023, we transplanted the seedlings into square pots (10 cm × 10 cm × 10 cm) filled with a 2:1:1 (v:v:v) mixture of peat, river sand, and a local soil, and covered the pots with shading nets to facilitate initial seedling establishment. The local soil was collected from the mountainous area of Taizhou and contained 0.62 ± 0.17 (mean ± SE) g kg^−1^ total nitrogen and 0.13 ± 0.03 g kg^−1^ total phosphorus. During the first two weeks, the survival status of each seedling was monitored daily and dead seedlings were replaced immediately. The shading net was then removed. During the experiment, we watered the plants once the soil surface of most pots dried out and supplied 50 mL of a 10% Hoagland nutrient solution to each pot once a month [62].
4.3. Harvest and Measurement
We harvested the aboveground parts of each species in each pot on 2 November 2023, after more than three months of growth. All plant materials were oven-dried at 70 °C for at least 48 h and weighed to obtain the biomass.
4.4. Data Analysis
To directly quantify the competitive effect, we calculated the log response ratio as follows: LogRR_ij_ = ln (Bij/Bi), where LogRR_ij_ is the log response ratio of species j on species i, Bij represents the aboveground biomass of species i in the presence of species j for each replicate, and Bi represents the mean aboveground biomass of target species i when grown alone (no competition) across the replicates. A three-way ANOVA was performed to analyze the effect of the presence each of the other three species (the 3rd species) and their two-way and three-way interactions on the competitive effect (LogRR) of the two target species. A significant two-way interaction effect indicates that the effect of the 3rd species on the competition between two target species depends on the presence of the 4th species, and a significant three-way interaction effect means that the effect of the 3rd species on the competition between two target species depends also on the presence of the 5th species. All analyses were carried out using SPSS Statistics 26.0 software (IBM Corp., Armonk, NY, USA).
5. Conclusions
We conclude that interspecific interactions between two plant species in a multi-species community may be commonly regulated by the presence of other species. However, it seems uncommon that the impact of the third plant species on competition is regulated by the presence of the fourth and/or the fifth plant species. This study highlights the complex interactions among multiple plant species within a community and also the importance of including these high-order interactions when modelling community dynamics and species coexistence.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ninkovic V. Rensing M. Dahlin I. Markovic D. Who is my neighbor? Volatile cues in plant interactions Plant Signal. Behav.201914610.1080/15592324.2019.163499331267830 PMC 6768235 · doi ↗ · pubmed ↗
- 2Bascompte J. Disentangling the web of life Science 200932541641910.1126/science.117074919628856 · doi ↗ · pubmed ↗
- 3Brooker R.W. Maestre F.T. Callaway R.M. Lortie C.L. Cavieres L.A. Kunstler G. Liancourt P. Tielbörger K. Travis J.M.J. Anthelme F. Facilitation in plant communities: The past, the present, and the future J. Ecol.200896183410.1111/j.1365-2745.2007.01295.x · doi ↗
- 4Adler P.B. Smull D. Beard K.H. Choi R.T. Furniss T. Kulmatiski A. Meiners J.M. Tredennick A.T. Veblen K.E. Competition and coexistence in plant communities: Intraspecific competition is stronger than interspecific competition Ecol. Lett.2018211319132910.1111/ele.1309829938882 · doi ↗ · pubmed ↗
- 5Fukano Y. Tachiki Y. Kasada M. Uchida K. Evolution of competitive traits changes species diversity in a natural field Proc. R. Soc. B-Biol. Sci.2022289810.1098/rspb.2022.1376 PMC 951562236168760 · doi ↗ · pubmed ↗
- 6Chesson P. Mechanisms of maintenance of species diversity. Annu Rev. Ecol. Syst.20003134336610.1146/annurev.ecolsys.31.1.343 · doi ↗
- 7Novoplansky A. Picking battles wisely: Plant behaviour under competition Plant Cell Environ.20093272674110.1111/j.1365-3040.2009.01979.x 19389051 · doi ↗ · pubmed ↗
- 8Golivets M. Wallin K.F. Neighbour tolerance, not suppression, provides competitive advantage to non-native plants Ecol. Lett.20182174575910.1111/ele.1293429516604 · doi ↗ · pubmed ↗