Class III Alcohol Dehydrogenase Modulates Renal Parietal Epithelial Cell Transformation During Chronic Alcohol Consumption in Mice
Midori Katsuyama, Takahisa Okuda, Masamichi Ishizaki, Kentaro Wada, Motoyo Maruyama, Toshio Akimoto, Youkichi Ohno, Takahito Hayashi, Takeshi Haseba

TL;DR
This study shows that Class III alcohol dehydrogenase in the kidney helps regulate testosterone levels and protects against kidney damage during chronic alcohol consumption in mice.
Contribution
The study reveals a novel role of ADH3 in testosterone metabolism and its impact on kidney health during chronic alcohol use.
Findings
ADH3-deficient mice showed increased parietal epithelial cells even without alcohol consumption.
Chronic alcohol consumption elevated testosterone levels and PECs in wild-type mice.
ADH3 appears to protect kidney structure by metabolizing testosterone, which is disrupted by alcohol.
Abstract
Class III alcohol dehydrogenase (ADH3), primarily localized in the liver and kidney, contributes to alcohol metabolism during chronic alcohol consumption (CAC). However, its role in kidney function remains unclear. This study investigated renal morphological changes associated with ADH3-mediated alcohol metabolism. Nine-week-old male wild-type (WT) and ADH3-deficient (Adh3-/-) mice were administered 10% ethanol for 1 month. Histological analyses were performed using periodic acid–Schiff (PAS) staining and electron microscopy. Serum biochemical parameters were also assessed. In WT mice, CAC induced an increase in cuboidal parietal epithelial cells (PECs) in Bowman’s capsule, along with elevated testosterone levels in both serum and urine. Adh3-/- mice showed increased PECs even in the control group, with similarly elevated serum testosterone in both control and ethanol-treated groups.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
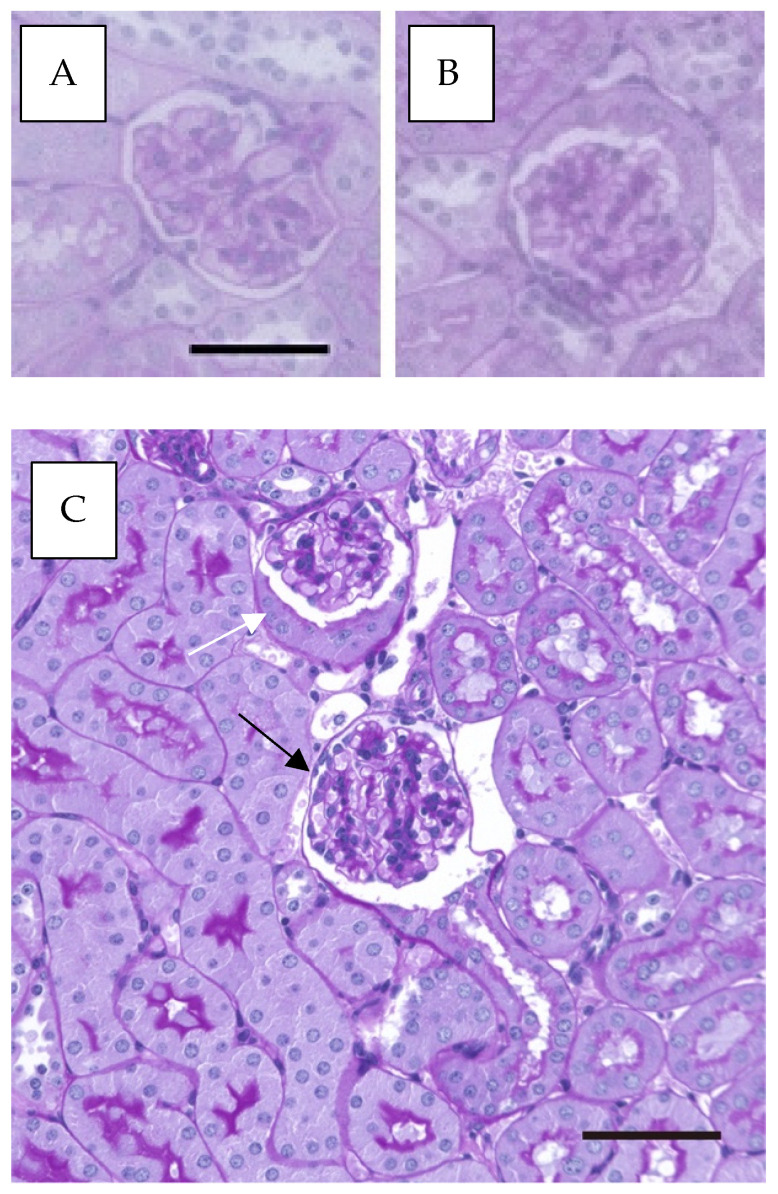 Figure 1
Figure 1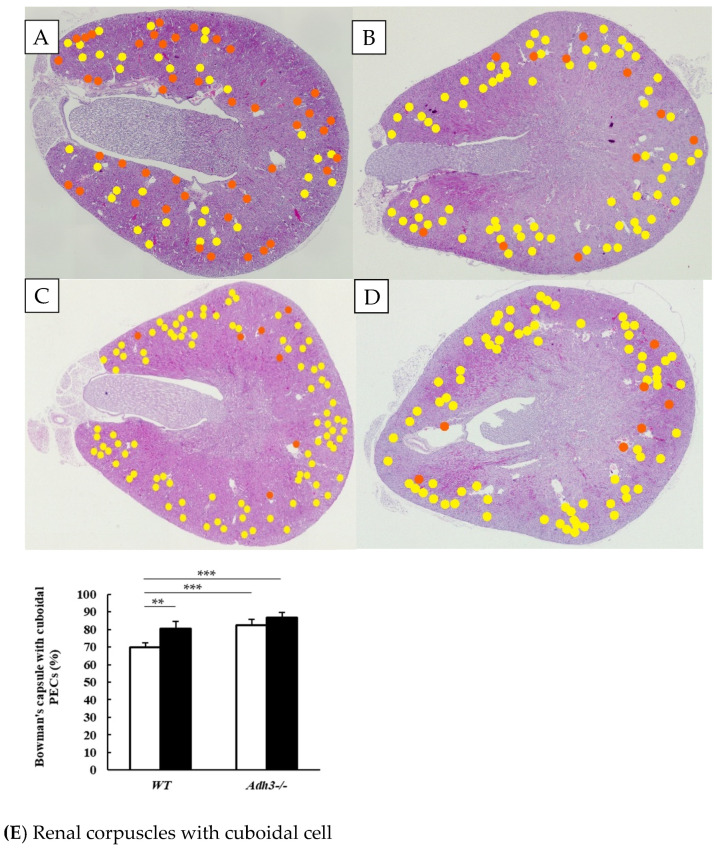 Figure 2
Figure 2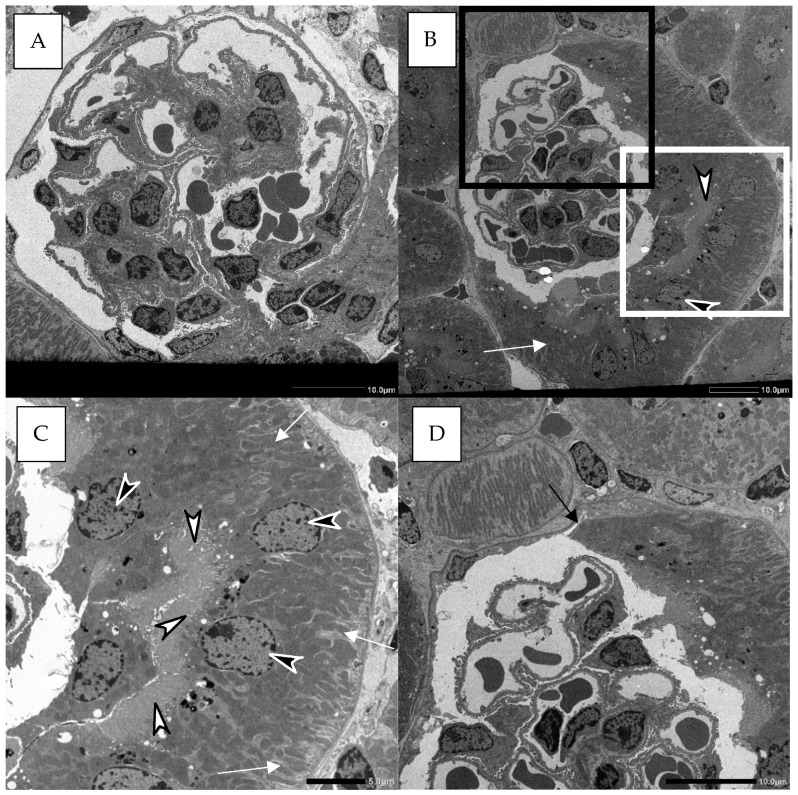 Figure 3
Figure 3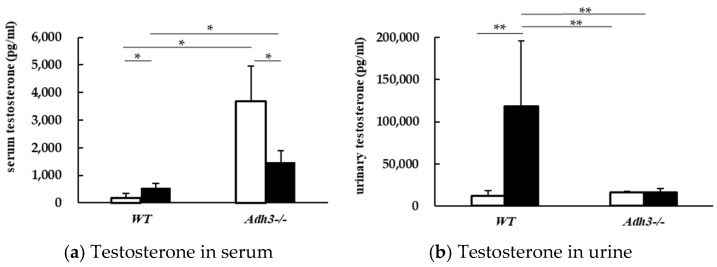 Figure 4
Figure 4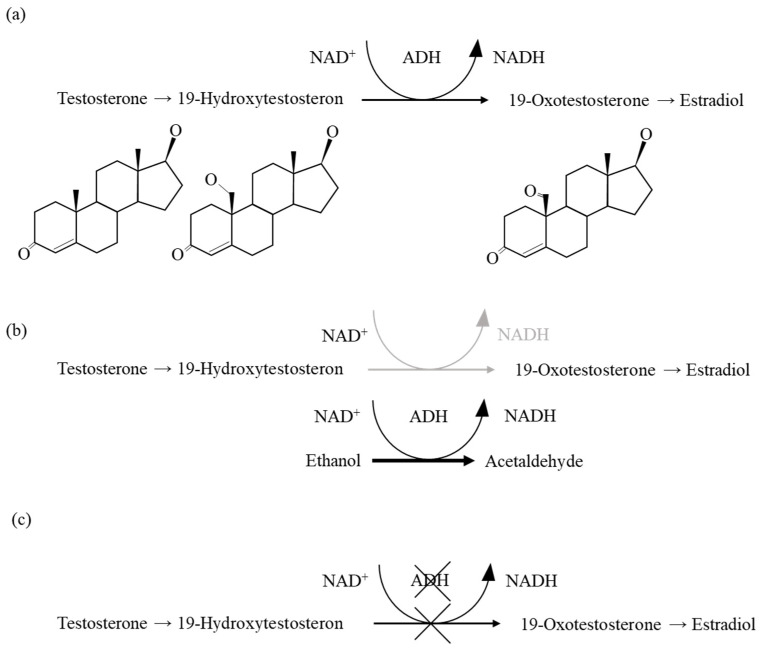 Figure 5
Figure 5- —Ministry of Education, Culture, Sports, Science and Technology (MEXT)
- —Japan Society for the Promotion of Science (JSPS) KAKENHI (Grants-in-Aid for Scientific Research)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAlcohol Consumption and Health Effects · Eicosanoids and Hypertension Pharmacology · Pharmacogenetics and Drug Metabolism
1. Introduction
The nephron is the microscopic structural and functional unit of the kidney, consisting of the renal corpuscle, proximal convoluted tubule, loop of Henle, and distal convoluted tubule. The renal corpuscle, where plasma filtration occurs, is composed of two main structures: the glomerulus and Bowman’s capsule. The parietal epithelial cells (PECs) lining Bowman’s capsule form a layer of flat epithelial cells that are continuous with the proximal tubular epithelium. PECs can undergo morphological changes, swelling and transforming into cuboidal-shaped cells with a crescent formation. This phenomenon has been specifically observed in sexually mature male laboratory mice [1,2,3]. These phenomena are infrequently observed in female mice. This observation suggests that the cuboidal transformation of PECs in male mice is associated with increased testosterone secretion during sexual maturation. Such changes are considered to represent a form of physiological adaptation. Moreover, these cuboidal PECs resemble the epithelium of the proximal convoluted tubule [4]. Although cuboidal PECs resemble proximal tubule cells, the significance of their reabsorptive function within the glomerulus—which primarily serves as a filtration unit—remains unclear. Furthermore, it is still uncertain whether these cells are indeed proximal tubules or a distinct cell population. Conversely, other studies suggest that the appearance of cuboidal PECs may be related to pathological conditions such as glomerulosclerosis [5]. Therefore, whether the presence of cuboidal PECs reflects a physiological adaptation or a pathological change remains unclear and warrants further investigation.
In general, alcohol consumption increases testosterone levels in vivo, whereas Mårdh et al. reported that testosterone allosterically regulates alcohol metabolism via alcohol dehydrogenases (ADHs; EC 1.1.1.1) [6]. Among the six ADH classes, class I (ADH1) is the primary enzyme responsible for alcohol metabolism. However, class III ADH (ADH3), which has the highest K_m_ for ethanol among the ADH family, also contributes to systemic alcohol metabolism in a dose-dependent manner in vivo [7,8], particularly during chronic alcohol consumption (CAC) [8,9]. ADH3 is expressed at the highest levels in the liver, with the second-highest expression observed in the kidney [10]. Recent studies have shown that low blood ethanol concentrations can upregulate ADH protein expression and activity in the kidney [8,11], suggesting that ADH3 may also participate in renal alcohol metabolism. Our previous work demonstrated that ADH3 exacerbates kidney damage during alcohol metabolism [12]. Notably, Swank et al. reported that androgens increase ADH activity in the kidney [13]. On the basis of these observations, we hypothesize that ADH3 increases testosterone levels through alcohol metabolism, thereby promoting the cuboidal transformation of PECs. Furthermore, evaluating whether this phenotypic change in PECs represents a physiological adaptation or a pathological alteration is of significant relevance in the field of alcohol-related metabolism research and renal pathology. The cuboidal transformation of PECs is occasionally observed not only in mice but also in humans, suggesting that the present study may also provide insights into the mechanisms underlying PECs cuboidalization in humans.
Based on these findings, we examined the relationship between androgens and the transformation of flat PECs into cuboidal PECs in Bowman’s capsule, particularly in the context of growth and alcohol exposure. To date, no studies have investigated the specific effects of alcohol consumption and ADH activity on PEC morphology in the kidney. Therefore, we conducted an experimental study using ethanol-administered mice with different ADH genotypes—wild-type (WT) and ADH3-deficient (Adh3^-/-^) C57BL/6N mice—to investigate whether ADH3-mediated alcohol metabolism influences the transformation of flat PECs into cuboidal PECs within the renal glomeruli.
2. Results
2.1. Alcohol Induces an Increase in Bowman’s Capsules with Cuboidal Parietal Epithelial Cells
Parietal epithelial cells (PECs) in Bowman’s capsule can undergo cellular activation, characterized by enhanced proliferative, migratory, and extracellular matrix-producing capacities [5]. In this study, we observed morphological changes in PECs following alcohol consumption in WT mice. As shown in Figure 1, PECs lining Bowman’s capsule were typically flat (Figure 1A, and black arrow in Figure 1C); however, in alcohol-treated mice, a transformation to cuboidal PECs was observed (Figure 1B and white arrow in Figure 1C).
Among the control groups, the number of Bowman’s capsules with cuboidal PECs was significantly higher in Adh3^-/-^ (C) mice compared to WT (C) mice (Control group: WT (C) and Adh3^-/-^ (C)). In WT mice, CAC significantly increased the number of Bowman’s capsules with cuboidal PECs (Figure 2A,B,E). However, in Adh3^-/-^ mice, alcohol consumption did not significantly alter this number (Figure 2C–E; CAC group: WT (E) and Adh3^-/-^ (E)).
2.2. Bowman’s Capsule with Cuboidal Parietal Epithelial Cells Observed by Electron Microscopy
Bowman’s capsules with PECs were examined using an electron microscope (Figure 3). In control WT mice, PECs were predominantly flat (Figure 3A), whereas in ethanol-treated mice, they appeared cuboidal in shape (Figure 3B,D). These cuboidal PECs closely resembled proximal tubular epithelial cells, characterized by abundant mitochondria and the presence of a brush border. Notably, the nuclei were aligned along the brush border (Figure 3B,C). Additionally, the origin of the cuboidal PECs, similar to those in the proximal tubule, extended inward from regions initially occupied by flat PECs (Figure 3D).
2.3. Alcohol Alters Testosterone Parameter
Regarding hormonal changes, serum and urinary testosterone levels were significantly increased in WT mice after alcohol consumption (Figure 4). Conversely, serum testosterone levels were significantly decreased in Adh3^-/-^ mice after alcohol exposure (Figure 4a). Notably, Adh3^-/-^ mice showed significantly higher serum testosterone levels compared to WT mice under control conditions (Figure 4b).
3. Discussion
Previous studies have demonstrated that flat PECs in the murine Bowman’s capsule can transform into proximal tubule-like cuboidal PECs [5]. This transformation has been reported to occur more frequently in male mice than in females [10], likely due to higher testosterone levels [4]. Elevated testosterone has been associated with the cuboidal transformation of PECs. In this study, electron microscopy revealed an increase in cuboidal PECs and brush borders in Bowman’s capsule following alcohol consumption. Additionally, serum and urinary testosterone levels were considerably higher in WT (E) mice compared to WT(C) mice. These findings suggest that the alcohol-induced elevation of testosterone levels may contribute to the increase in cuboidal PECs.
The role of ADH3 in the kidney has been underexplored, with few studies addressing its impact. Katsuyama et al. demonstrated that ADH3 may contribute to renal pathological changes, including increased glomerular permeability induced by CAC, potentially exacerbating alcoholic kidney disorders [12]. Dinu et al. showed that ethanol consumption (2 g/kg) increased ADH activity in the kidney [14]. This study investigated the renal morphological features in ADH3-deficient mice with or without alcohol consumption. We observed a significant increase in the number of cuboidal PECs in the Bowman’s capsules of Adh3^-/-^ (C) mice, and their serum testosterone levels were significantly increased compared to WT (C) mice. These findings suggest that ADH3 regulates testosterone levels, potentially via its role in alcohol metabolism. It is possible that alcohol increases testosterone levels though ADH3-dependent metabolism, leading to the cuboidal transformation of PECs. Previous studies have indicated that testosterone allosterically regulates ethanol oxidation by alcohol dehydrogenase containing the γ subunit [6]. Furthermore, 19-hydroxytesosterone is a competitive substrate for ethanol in ADH [15]. In testosterone metabolism, the NAD-dependent oxidation of 19-hydroxytestosterone to 19-oxotestosterone is mediated by ADH; therefore, it may be competitively inhibited by ethanol, which is also NAD-dependently oxidized by ADH. Thus, CAC is thought to increase concentrations of both 19-hydroxytestosterone and testosterone.
This study indicated that ethanol consumption in WT (E) mice significantly increased serum testosterone levels compared to WT (C) mice. However, in Adh3^-/-^ (C) mice, the lack of ADH3 hindered testosterone metabolism, resulting in a significantly higher serum testosterone level. Based on our findings, we propose a two-step mechanism by which testosterone affects the mouse renal system (Figure 5). Consequently, the high testosterone levels observed in the Adh3^-/-^ (C) mice were associated with an increased number of cuboidal PECs.
Recent reports have proposed two mechanisms for the formation of cuboidal PECs: physiological differentiation [4,10] and renal dysfunction [5,16,17]. Hanker et al. suggested that an increase in PECs, associated with foot process effacement and glomerulosclerosis, could result in renal dysfunction [5,16,17]. However, despite the increased cuboidal PECs in the Adh3^-/-^ (C) mice, they did not exhibit signs of renal dysfunction [12]. Additionally, while the Adh3^-/-^ (E) mice showed some mitochondrial collapse in proximal tubular cells under electron microscopy, the damage was less severe compared to the WT (E) mice. This suggests that alcohol consumption has a less significant impact in the absence of kidney ADH3.
Previous studies have reported that gross structural abnormalities or microscopic lesions in the kidney were not evident during alcohol consumption [18], suggesting that renal dysfunction may be less apparent. However, alcohol consumption could lead to albumin and proteins leakage into the urine, as well as increased blood urea nitrogen (BUN) and serum creatinine levels [19]. In our previous study, albumin and protein were significantly leaked into the urine after alcohol consumption in WT mice [12], consistent with these findings. In contrast, ADH3 deficiency did not significantly lead to urinary albumin and protein leakage. Since electrolyte balance and osmotic pressure were generally stable, it is likely that the reabsorptive function of the proximal tubule remained normal in ADH3-deficient mice. Therefore, alcohol consumption causes urinary abnormalities rather than overt renal disorders in this experimental condition. Accordingly, WT mice with alcohol consumption exhibited more abnormal renal function parameters compared to Adh3^-/-^ (E) mice. Therefore, our findings suggest that ADH3 metabolizes alcohol during CAC and may exacerbate renal dysfunction.
Furthermore, alcohol consumption may induce an increased number of cuboidal PECs in the presence of ADH3. Notably, in the absence of ADH3 and without alcohol ingestion, high serum testosterone levels were also associated with an increase in cuboidal PECs. During alcohol metabolism, ADH3 in the kidney may be involved in the metabolism of ethanol, and its primary physiological role in testosterone metabolism could be inhibited. This inhibition, in turn, may contribute to pathological abnormalities in the kidney. Therefore, ADH3 may be a key factor in alcohol-induced renal dysfunction. Although it has been previously reported that alcohol consumption increases testosterone levels, our study is the first to identify a role for ADH3 in this process. Furthermore, the finding that its mechanism may induce morphological changes in PECs provides important new insights from the perspective of renal pathology.
ADH3 induces pathological changes in the renal proximal tubules by metabolizing alcohol at this site, leading to increased glomerular permeability during CAC. However, in Adh3^-/-^ mice, no significant changes were observed in podocyte dilation, creatinine levels, or BUN, indicating the absence of overt renal dysfunction [12].
For example, it takes time for diabetic nephropathy to become clinically recognizable as renal damage. To better compare our findings, we plan to administer alcohol for one year instead of one month in future studies. Additionally, since short-term and long-term alcohol consumption may have different effects on testosterone levels, a one-year alcohol administration study may provide further insight into the long-term impact of alcohol and ADH3 on testosterone metabolism and renal morphology.
4. Materials and Methods
4.1. Animals
An ADH3-deficient mouse (Adh3^-/-^) contained homozygous null mutants for the Adh3 genes. This mouse was originally obtained from the Burnham Institute, USA, in 1999 [20], and subsequently backcrossed with the C57BL/6N background strain (WT mice, Sankyo Lab., Co., Ltd., Tokyo, Japan) for up to 13 generations. The congenic Adh3^-/-^ strain has been maintained at Nippon Medical School (Tokyo, Japan) since 2013 and has been bred at Kagoshima University (Kagoshima, Japan) since 2020.
Mice of two genotypes (WT and Adh3^-/-^) were weaned at 4 weeks of age and housed in groups of 4–6 per cage under specific pathogen-free conditions. The environment was maintained at 24 °C ± 2 °C with a 14 h light/10 h dark cycle starting at 7:00 AM. Mice were provided a standard rodent diet (MF pellets, Oriental Yeast Co., Ltd., Tokyo, Japan) containing 12.5% fat calories and water ad libitum. For each experiment, five mice were used per group.
All animal experiments were conducted in compliance with the ARRIVE guidelines and were approved by the Institutional Committee of Laboratory Animals at Nippon Medical School (Approval No. 28-001) and the Animal Ethics Committee of the Division of Laboratory Animal Resources and Research at Kagoshima University (Approval No. MD23061) following the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publication No. 8023, revised 1978).
4.2. Chronic Alcohol Consumption Experiment
CAC experiments were performed by providing 9-week-old male mice with a 10% (w/v) ethanol solution in water ad libitum as their only drinking source for one month, following the method previously described [21]. To acclimate the mice, the ethanol concentration was gradually increased by 2% per day over the course of 1 week until reaching 10%. Mice receiving the ethanol solution were assigned to the CAC group (E group): WT (E) and Adh3^-/-^ (E). Mice provided only water served as controls (C group): WT (C) and Adh3^-/-^ (C).
4.3. Tissue Preparation
After 1 month of ethanol consumption, WT and Adh3^-/-^ mice were anesthetized, had blood collected from the retro-orbital venousplexus, and were euthanized by cervical dislocation. For light microscopic examination, kidneys were fixed in 10% neutral-buffered formalin (Cat. No. 068-01663, Wako, Osaka, Japan). Within two weeks, the fixed tissues were dehydrated using a vacuum rotary processor (Sakura Finetek Japan Co., Ltd., Tokyo, Japan) and embedded in paraffin. Paraffin blocks were sectioned at a thickness of 2 μm and stained with periodic acid–Schiff (PAS) stain or hematoxylin and eosin (HE) stain for morphological evaluation following the manufacturer’s protocol (PAS stain: periodic acid, Cat. No. 86171; Schiff regent, Cat. No. 40921; sulfurous acid solution, Cat. No. 40941; Mayer’s hematoxylin, Cat. No. 30002. HE stain: Carrazzi’s hematoxylin, Cat. No. 30022; Eosin Y, Cat. No. 31901. Muto Pure Chemicals Co., Ltd., Tokyo, Japan). Digital images were captured using a BZ-9000 All-in-One Microscope (Keyence Corporation, Osaka, Japan).
For electron microscopic examination, 1 mm^3^ kidney samples were fixed in 2.5% glutaraldehyde (Cat. No. 303-2, Nisshin-EM Co., Ltd., Tokyo, Japan; product by TAAB Laboratories Equipment Ltd., Berks, UK) prepared in 0.1 M phosphate buffer (Cat. No. 80362, Muto Pure Chemicals Co., Ltd., Tokyo, Japan). Tissues were then post-fixed in 1% osmium tetroxide (Cat. No. 300, Nisshin-EM Co. Ltd., Tokyo, Japan; product by Heraeus South Africa (Pty) Ltd., Port Elizabeth, South Africa), dehydrated in a graded ethanol series, and embedded in Oken Epok 812 resin (Cat. No. 99-02-1001, Okenshoji Co., Ltd., Tokyo, Japan). Ultrathin sections (70 nm) were prepared using a Leica Ultracut R ultramicrotome (Leica, Wetzlar, Germany), stained with uranyl acetate and lead citrate, and examined using a JEM-1400 Plus electron microscope (JEOL Ltd., Tokyo, Japan).
4.4. Measurement of the Ratio of Bowman’s Capsules with Cuboidal PECs and Flat PECs
The number of Bowman’s capsules lined with cuboidal or flat PECs was counted in each group using a mechanical tally counter. All classifications were conducted through visual inspection using a microscope. PECs exhibiting even slight deviations from a flat morphology were classified as cuboidal. Image analysis was performed on hematoxylin and eosin (HE)-stained sections using Adobe Photoshop (version 21.0.1; Adobe Systems, Inc., San Jose, CA, USA).
4.5. Measurement of Biochemical Parameters in Serum and Urine
Blood samples were collected and centrifuged at 3000× g for 15 min to obtain serum. A mouse metabolic gauge (Shinano Manufacturing Co., Ltd., Tokyo, Japan) was used to collect 24-h urine samples one month after the initiation of ethanol administration. Urine collection was conducted in two intervals: from 10:00 am to 18:00 pm and from 18:00 pm to 12 noon the following day. The concentrations of serum and urinary testosterone were measured using a Testosterone Enzyme Immunoassay Kit (Cat. No. K032-H1, Funakoshi Co., Ltd., Tokyo, Japan; product by Arbor Assays, Ann Arbor, MI, USA). Absorbance was measured with a SpectraMax^TM^ i3x microplate reader (Molecular Devices Japan, Tokyo, Japan) according to the manufacturer’s instructions.
4.6. Statistical Analysis
All continuous variables are presented as the mean ± standard deviation (SD). Differences between the control and ethanol-treated groups were analyzed using one-way analysis of variance, followed by Tukey’s Honest Significant Difference test for post hoc comparisons. Statistically significant differences are indicated in each figure. All statistical analyses were performed using JMP software, version 14 (SAS Institute Inc., Cary, NC, USA).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahmadizadeh M. Echt R. Kuo C.H. Hook J.B. Sex and strain differences in mouse kidney: Bowman’s capsule morphology and susceptibility to chloroform Toxicol. Lett.19842016117110.1016/0378-4274(84)90142-56695407 · doi ↗ · pubmed ↗
- 2Hamada Y. Sex difference and fine structure on epithelium cells of Bowman’s capsule in mice (author’s transl)Jikken Dobutsu 19792848549052767210.1538/expanim 1978.28.4_485 · doi ↗ · pubmed ↗
- 3Ichii O. Yabuki A. Ojima T. Matsumoto M. Suzuki S. Rodent renal structure differs among species J. Vet. Med. Sci.20066843944510.1292/jvms.68.43916757886 · doi ↗ · pubmed ↗
- 4Dietert S.C. The columnar cells occurring in the parietal layer of Bowman’s capsule. Cellular fine structure and protein transport J. Cell Biol.19673543544410.1083/jcb.35.2.4354861776 PMC 2107126 · doi ↗ · pubmed ↗
- 5Kuppe C. Leuchtle K. Wagner A. Kabgani N. Saritas T. Puelles V.G. Smeets B. Hakroush S. van der Vlag J. Boor P. Novel parietal epithelial cell subpopulations contribute to focal segmental glomerulosclerosis and glomerular tip lesions Kidney Int.201996809310.1016/j.kint.2019.01.03731029503 PMC 7292612 · doi ↗ · pubmed ↗
- 6Mårdh G. Falchuk K.H. Auld D.S. Vallee B.L. Testosterone allosterically regulates ethanol oxidation by homo- and heterodimeric gamma-subunit-containing isozymes of human alcohol dehydrogenase Proc. Natl. Acad. Sci. USA 1986832836284010.1073/pnas.83.9.28362939458 PMC 323401 · doi ↗ · pubmed ↗
- 7Haseba T. Kameyama K. Mashimo K. Ohno Y. Dose-Dependent Change in Elimination Kinetics of Ethanol due to Shift of Dominant Metabolizing Enzyme from ADH 1 (Class I) to ADH 3 (Class III) in Mouse Int. J. Hepatol.2012201240819010.1155/2012/40819022164338 PMC 3227458 · doi ↗ · pubmed ↗
- 8Okuda T. Haseba T. Katsuyama M. Maruyama M. Akimoto T. Igarashi T. Ohno Y. Metabolic pharmacokinetics of early chronic alcohol consumption mediated by liver alcohol dehydrogenases 1 and 3 in mice J. Gastroenterol. Hepatol.2018331912191910.1111/jgh.1426029663519 · doi ↗ · pubmed ↗