Skeletal Muscle Alterations in Different Phenotypes of Heart Failure with Preserved Ejection Fraction
Beatrice Vahle, Romy Klädtke, Antje Schauer, T. Scott Bowen, Ulrik Wisløff, Axel Linke, Volker Adams

TL;DR
This study compares two rat models of heart failure with preserved ejection fraction and finds differences in skeletal muscle changes linked to their underlying causes.
Contribution
The study reveals distinct skeletal muscle alterations in two HFpEF rat models with different disease etiologies.
Findings
Obese ZSF-1 rats showed muscle atrophy, reduced force, and increased titin phosphorylation compared to controls.
Hypertensive DSS rats did not show similar skeletal muscle changes as the ZSF-1 model.
Fiber type-specific proteins and myostatin were differentially regulated between the two models.
Abstract
Heart failure with preserved ejection fraction (HFpEF) shows diverse disease patterns, with various combinations of comorbidities and symptoms. A common hallmark is exercise intolerance, caused by alterations in the peripheral skeletal muscle (SKM) including a recently indicated titin hyperphosphorylation. Our aim is to compare a metabolic syndrome- (ZSF-1 rats) and a hypertension-driven (Dahl salt-sensitive (DSS) rats) HFpEF rat-model in relation to SKM function and titin phosphorylation. Obese ZSF-1 and high-salt fed DSS rats (HFpEF) were compared to lean ZSF-1 and low-salt fed rats (con). HFpEF was confirmed by echocardiography and invasive haemodynamic measurements. SKM atrophy, in vitro force measurements, titin- and contractile protein expression were evaluated. Obese ZSF-1 HFpEF rats showed muscle atrophy, reduced muscle force and increased titin phosphorylation compared to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
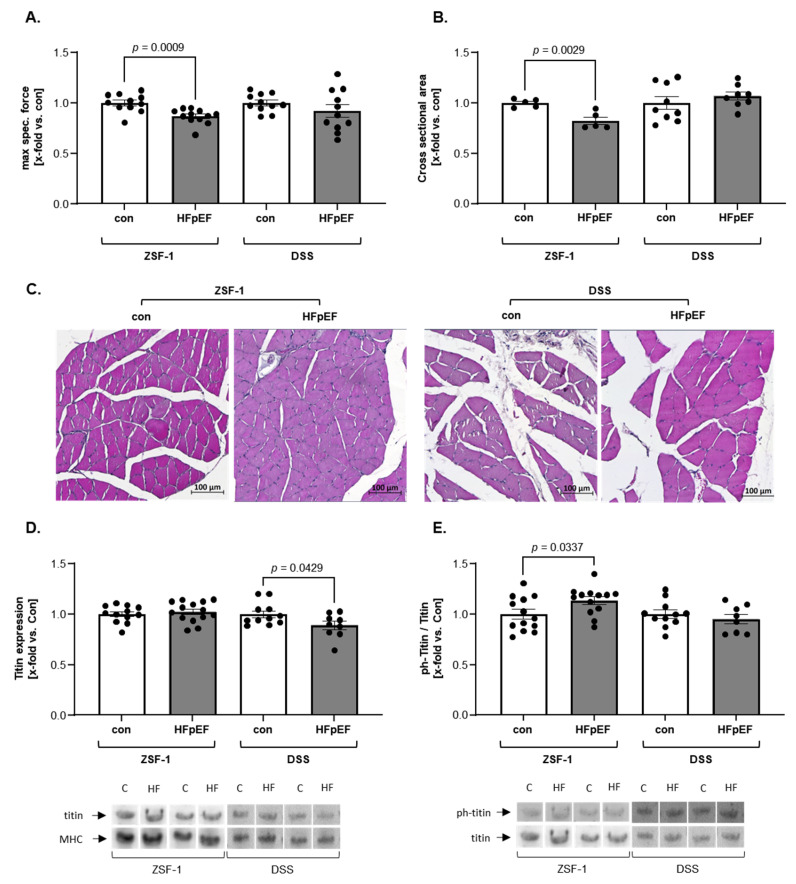 Figure 1
Figure 1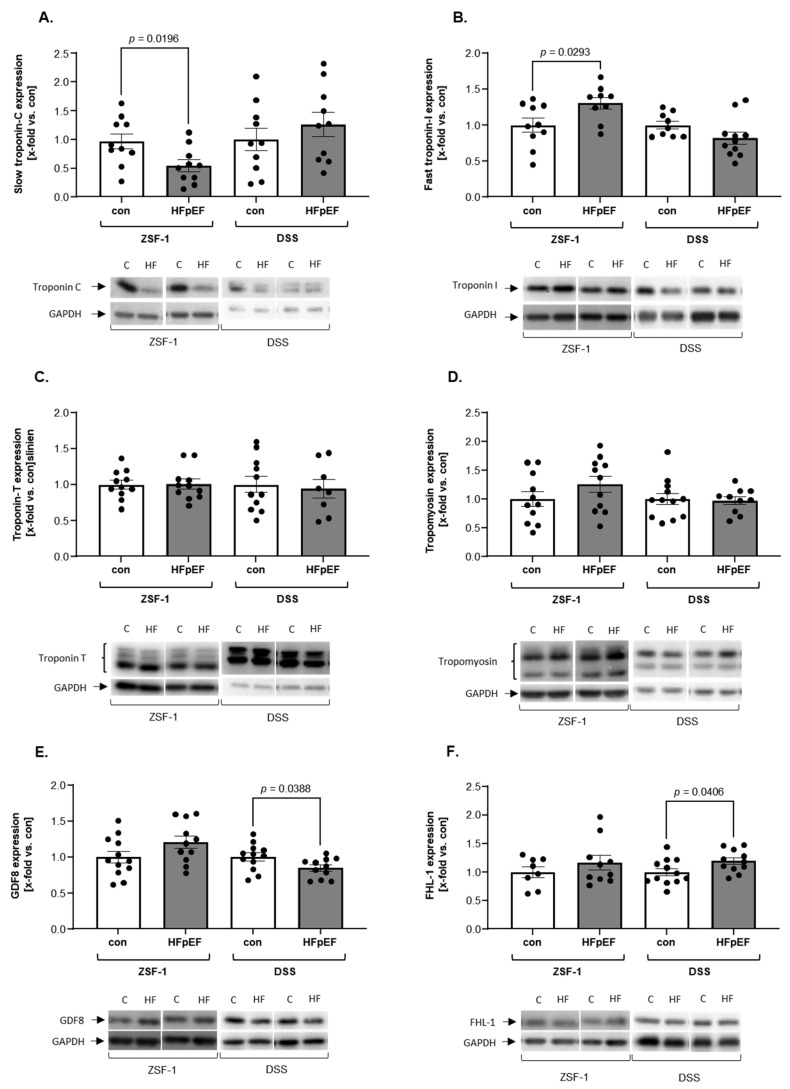 Figure 2
Figure 2- —European Commission, Framework Programme 7
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCardiomyopathy and Myosin Studies · Cardiovascular Function and Risk Factors · Muscle Physiology and Disorders
1. Introduction
Heart failure (HF) with preserved ejection fraction (HFpEF) is a systemic syndrome resulting from various combinations of comorbidities, such as aging, obesity, diabetes, metabolic syndrome, and hypertension [1]. Clustering into phenogroups, based on the occurring comorbidities and symptoms is pursued, to enable personalized treatment and to optimize the clinical outcome [2].
A common hallmark among all groups is exercise intolerance, partly due to alterations in the peripheral skeletal muscle (SKM) and increased myocardial and skeletal titin stiffness via isoform switch and/or phosphorylation [3,4,5]. Recently, our group reported hyperphosphorylation of titin in limb SKM of an animal model of HFpEF, due to enhanced phosphorylation at the PEVK region known to lead to titin filament stiffening [5]. The relevance of titin hyperphosphorylation in the SKM is further supported by a negative correlation between the phosphorylation state of titin and muscle force. To study HFpEF and to develop possible therapies, different animal models have been employed. The obese ZSF-1 rat and the Dahl salt-sensitive rat (DSS) are two widely used preclinical animal models for translational research in HFpEF [6], that develop the disease due to different comorbidities. Metabolic syndrome is the driving factor in the ZSF-1 rat, whereas the DSS rat model develops HFpEF due to hypertension. The question arises whether the molecular alterations in the SKM and specifically the titin hyperphosphorylation occur in both HFpEF models or if the different comorbidities result in different SKM alterations. Therefore, the aim of the present study is to analyze molecular alterations in the SKM of both HFpEF models with a specific focus on titin phosphorylation and muscle contraction regulating proteins.
2. Results
The cardiac function of both animal models exhibited distinct features of HFpEF compared to their respective controls apparent by signs of diastolic dysfunction (increased E/é and LVEDP), cardiac hypertrophy (increased heart weight), and a preserved ejection fraction (LVEF > 60%). Only ZSF-1 HFpEF rats showed signs of a metabolic syndrome evident by increased body weight, blood pressure, and glucose levels, whereas DSS rats presented only with a substantial increase in blood pressure (Table 1).
With respect to muscle atrophy and function we observed a decrease in soleus SKM specific force and cross-sectional area (CSA) in ZSF-1 HFpEF rats, but not in DSS HFpEF rats (Figure 1A,B). In contrast, titin showed a lower total expression in DSS HFpEF rats only (Figure 1C). With respect to titin phosphorylation ZSF-1-, but not DSS HFpEF rats, appeared to have an increased phosphorylation relative to respective controls (Figure 1D).
With respect to contractile muscle- and mass regulating proteins, expression patterns of the troponin–tropomyosin complex (Tn–Tm) (Figure 2A–D) and atrophy-markers (Figure 2E,F) showed contradictory results between models. While slow troponin C (TnC) expression was reduced in ZSF-1 HFpEF rats compared to relative controls, fast troponin I (TnI) expression was higher. DSS rats showed no changes compared to their controls. No animal model showed changes in Troponin T (TnT) or tropomyosin (Tm) expression (Figure 2A–D). With respect to mass regulating proteins, Myostatin (GDF8) was slightly upregulated (p = 0.09) in the ZSF-1 HFpEF rat, while FHL1 showed no change in its expression. In contrast, the DSS rats exhibited a reduced GDF8 expression, whereas FHL-1 expression was enhanced in HFpEF rats compared to controls (Figure 2E,F).
3. Discussion
Defining the most suitable animal for HFpEF is of great relevance for testing potential treatment regimens for HFpEF. In the current literature several different animal models are described, which are all based on different comorbidities driving the development of HFpEF. We previously compared skeletal muscle samples of different HFpEF animal models with human muscle biopsies, primarily investigating molecular SKM parameters and myocardial features and concluded that ZSF-1 rats seemed to offer the most suitable HFpEF model to study therapeutic interventions, especially regarding SKM alterations [6]. In accordance with these previous observations, our current findings revealed significant changes in SKM function, atrophy, and a titin hyperphosphorylation only in ZSF-1- but not in DSS rats with HFpEF.
Titin stiffness can be modulated by posttranslational modification, primarily phosphorylation. In the myocardium of HFpEF a hypophosphorylation was documented [7], whereas in the peripheral SKM our group recently reported a hyperphosphorylation in the tibialis anterior muscle of HFpEF rats [5]. Furthermore, a negative correlation between muscle force and atrophy and the titin hyperphosphorylation was evident. In accordance with these earlier observations, a titin hyperphosphorylation was seen in the current study in the soleus of ZSF-1 HFpEF rats but not in the soleus of the DSS HFpEF rats. This goes along with the observation that in the DSS model no impact on muscle atrophy and muscle function was noted. Further studies evaluating titin phosphorylation in human HFpEF patients are necessary to clarify the importance of titin hyperphosphorylation for modulating muscle atrophy and function.
Besides the well-known muscle mass regulating atrogene muscle RING-finger-protein-1, which have been found to be upregulated in the ZSF-1 and the DSS HFpEF rats [6], GDF8 (also known as myostatin) is another potent modulator of muscle mass [8,9]. In accordance with the induction of muscle atrophy only in the ZSF-1 HFpEF rats, GDF8 expression was slightly elevated in the ZSF-1 HFpEF model but not in the DSS rat model, where even a reduced GDF8 expression was observed. FHL-1 is assumed to activate GDF8 in SKM [10] but is also associated with myocardial hypertrophy [11]. Mutations and decreased expression on the other hand are related with muscular impairment [11]. The increased FHL-1 levels in DSS HFpEF rats may be a sign of counter regulation to the impaired GDF8 expression.
With respect to the initiation of muscle contraction, the Tn–Tm complex is important [12]. Our findings in ZSF-1 HFpEF rats may indicate a shift to fast fibers, since the expression of troponin isoforms depends on the prevalent muscle fiber type. Despite in humans with HFpEF where a fiber-type shift is associated with lower exercise performance [13], no fiber type shift has been reported in DSS rats with HFpEF [14] which may explain some of our findings here.
We confirmed, as previously described, elevated glucose levels in obese rats. Furthermore, obese ZSF-1 rats are characterized by hyperinsulinemia and insulin resistance, as well as glucose intolerance, which is supported by elevated insulin and glucose levels [15,16]. With respect to DSS rats, we detected no alteration of glucose levels in DSS rats with HFpEF. This is in accordance with other studies also describing normal glucose and insulin levels [17]. With respect to muscle force, which was only reduced in ZSF-1 rats, glucose is an important factor for energy delivery, serving as fuel for contraction. Together, this may be an explanation for the reduced force occurring in ZSF-1 HFpEF rats.
Overall, the present study reveals that SKM impairments are not only on the molecular level in obese ZSF1 rats with HFpEF but also in physiological parameters and in sarcomeric titin phosphorylation. In conclusion, our data indicate that skeletal muscle (SKM) deficits in HFpEF are influenced more by the underlying etiology of the disease than by compromised cardiovascular function.
Study Limitations
Despite our important findings regarding skeletal muscle alterations in different HFpEF animal models, with a focus on titin, some limitations have to be mentioned. First, for the characterization of limb muscle function and some molecular alterations, different muscle groups were used (EDL and Sol), due to limited material availability. Nevertheless, different studies have shown that an overall dysregulation in the limb muscle appears, independent of the analyzed muscle group [6,18,19].
Furthermore, only female rats were used in the present study. As a consequence, it is not possible to generalize the results to the total HFpEF population. Nevertheless, registry as well as community-based studies have shown that the plurality of HFpEF patients are elderly women exhibiting a variety of comorbidities, such as hypertension, diabetes, pulmonary disease, chronic kidney disease, and obesity [20,21].
4. Materials and Methods
4.1. Animals
32-week-old female ZSF1 lean (control, n = 13) and ZSF-1 obese (HFpEF, n = 14) rats were purchased from Charles River (Charles River Laboratories, USA) and included into the present study. They were compared with 35-week-old female Dahl salt-sensitive rats (DSS), that were fed with chow, containing 0.3% NaCl (low salt (LS) (control), n = 12) or 8% NaCl (high salt (HS) (HFpEF), n = 11), randomly selected at 7 weeks of age. Both models had access to food and water ad libitum. Before sacrificing, the development of HFpEF was confirmed by echocardiography and invasive haemodynamic measurements
4.2. Echocardiography
Following anesthesia with isoflurane (1.5–2%), cardiac function was assessed by transthoracic echocardiography using a Vevo 3100 imaging system equipped with a 21 MHz transducer (VisualSonics, FujiFilm), as outlined previously [16]. Systolic performance was evaluated by acquiring B-mode and M-mode images of the parasternal long- and short-axis planes at the level of the papillary muscles. For the assessment of diastolic function, pulsed-wave Doppler was utilized in the apical four-chamber view to record mitral inflow velocities (early E wave), while tissue Doppler imaging measured myocardial early relaxation velocity (E′) at the basal septal region of the left ventricle. Quantification of cardiac parameters, such as left ventricular ejection fraction (LVEF) and the E/e′ ratio, was performed using Vevo LAB software version 5.10.0.
4.3. Invasive Haemodynamic Measurements
Terminal invasive haemodynamic measurements were conducted in anesthetized rats (ketamine and xylazine) with maintained spontaneous respiration. A rat-specific PV catheter (SPR-838; ADInstruments Limited) was introduced via the right carotid artery and carefully advanced into the mid-cavity of the left ventricle. Left ventricular end-diastolic and end-systolic pressures were subsequently recorded. All pressure data were collected and analyzed using LabChart8 software (ADInstruments).
4.4. Skeletal Muscle Function
Muscle function from soleus was assessed ex vivo as recently described [19]. Briefly the muscle was dissected and mounted vertically in a Krebs–Henseleit buffer-filled organ bath (1205A: Isolated Muscle System—Rat, Aurora Scientific Inc., Aurora ON, Canada). Platinum electrodes stimulated the muscle with a supra-maximal current (700 mA, 500 ms train duration, 0.25 ms pulse width) from a high-power bipolar stimulator (701C; Aurora Scientific Inc., Aurora ON, Canada) to assess muscle function. The muscle was cocked up to an optimal length (Lo) which is defined by the maximal produced twitch force. To determine maximal force, a force-frequency protocol, with rest intervals of 1 min, was performed at 1, 15, 30, 50, 80, 120, and 150 Hz.
4.5. Histological Analyses
To assess the atrophy of the SKM, histological analyses to define the myofiber CSA were performed. Therefore, paraffin-embedded Sol tissue was sectioned (4 µm) and mounted on glass slides. Afterwards a hematoxylin and eosin stain were performed. The measurement of the CSA from the fibers was evaluated through imaging software (Zen imaging software, Zeiss, Jena, Germany). At least 60 fibers per section were measured.
4.6. Titin Analysis
For the assessment of total titin phosphorylation and expression, a vertical agarose gel electrophoresis (VAGE) was performed. Snap frozen soleus tissue was pulverized and dissolved in urea buffer (8 mol/L urea, 2 mol/L thiourea, 0.05 mol/L Tris pH 6.8, 0.075 mol/L DTT, 3% SDS), containing a phosphatase- and protease-inhibitor mixture (Serva, Heidelberg, Germany), at a ratio of 1:50 (weight/volume). The tissue and the buffer were mixed by carefully inverting the tubes several times and subsequently heated at 60 °C for 10 min to enable full solubilization of the tissue. After a short centrifugation step (30 s at 4000× g), a small aliquot of the supernatant was taken for protein quantification according to the manufacturer’s recommendation (660 nm protein assay, Thermo Fischer Scientific, Waltham, MA, USA). Afterward, glycerol, laced with traces of bromophenol blue, was added (final concentration 25%), followed by another centrifugation step (10 min, 13.200× g). The supernatant was collected, aliquoted and stored at −80 °C. When running the gels, 4 to 6 µg of protein was loaded on 1% agarose gels as described by Zhu and Guo [22] at a current of 15 mA for 5 h. To determine the quantity of phosphorylated titin, the gels were phosphor-stained with Pro-Q™ Diamond Phosphoprotein Gel Stain followed by a Sypro Ruby gel stain to detect total protein expression of titin and MHC (both Thermo Fischer Scientific, Waltham, MA, USA) according to the manufacturer’s recommendation. Measurements for total titin were normalized to MHC expression, whereas phosphorylation was normalized to the amount of unmodified protein.
4.7. Protein Analysis
For all other proteins, a standard SDS-PAGE was performed. Frozen EDL tissue was homogenized in RIPA buffer (50 mmol/L Tris pH 7.4, 1% NP-40, 0.25% Na-deoxycholate, 150 mmol/L NaCl, 1 mmol/L EDTA) supplemented with a protease- and phosphatase-inhibitor mixture (Inhibitor mix M, Serva, Heidelberg, Germany). A BCA assay was performed to assess protein concentration (BCA assay, Pierce, Bonn, Germany), and 10–20 μg of protein were loaded onto a gel.
Afterwards, the proteins were transferred to a PVDF membrane with a semi-dry blotter (VWR). The following primary antibodies were used: myostatin (GDF8) (19142-1-AP, 1:1000), four and a half LIM domain 1 (FHL-1) (10991-1-AP, 1:1000), tropomyosin (11038-1-AP, 1:2000), troponin C (13504-1-AP, 1:1000), troponin T (15513-1-AP, 1:1000) (all Proteintech, Germany), and troponin I (ab184554, 1:2000, Abcam, UK). According to the primary antibody, the membranes were afterwards incubated with a horseradish peroxidase-conjugated secondary antibody. Finally, the protein signal was visualized via enzymatic chemiluminescence (Super Signal West Pico, Thermo Fisher Scientific Inc., Bonn, Germany), and densitometry was used for quantification (1D scan software package version 15.08b; Scanalytics Inc., Rockville, MD, USA). GAPDH (1:10,000; HyTest Ltd., Turku, Finland) served as loading control to normalize the measurements. The data are presented as x-fold change relative to control.
4.8. Statistical Analyses
Statistical analysis was performed between groups in each animal model via unpaired, two-tailed t-tests, using GraphPad Prism 10.2.3. The values are reported as mean ± standard error of the mean (SEM) and p < 0.05 was considered significant.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Pieske B. Tschöpe C. de Boer R.A. Fraser A.G. Anker S.D. Donal E. Edelmann F. Fu M. Guazzi M. Lam C.S.P. How to diagnose heart failure with preserved ejection fraction: The HFA-PEFF diagnostic algorithm: A consensus recommendation from the Heart Failure Association (HFA) of the European Society of Cardiology (ESC)Eur. Heart J.2019403297331710.1093/eurheartj/ehz 64131504452 · doi ↗ · pubmed ↗
- 2Segar M.W. Patel K.V. Ayers C. Basit M. Tang W.W. Willett D. Berry J. Grodin J.L. Pandey A. Phenomapping of patients with heart failure with preserved ejection fraction using machine learning-based unsupervised cluster analysis Eur. J. Heart Fail.20202214815810.1002/ejhf.162131637815 · doi ↗ · pubmed ↗
- 3Loescher C.M. Hobbach A.J. Linke W.A. Titin (TTN): From molecule to modifications, mechanics, and medical significance Cardiovasc. Res.20221182903291810.1093/cvr/cvab 32834662387 PMC 9648829 · doi ↗ · pubmed ↗
- 4Del Buono M.G. Arena R. Borlaug B.A. Carbone S. Canada J.M. Kirkman D.L. Garten R. Rodriguez-Miguelez P. Guazzi M. Lavie C.J. Exercise Intolerance in Patients With Heart Failure: JACC State-of-the-Art Review J. Am. Coll. Cardiol.2019732209222510.1016/j.jacc.2019.01.07231047010 · doi ↗ · pubmed ↗
- 5Vahle B. Heilmann L. Schauer A. Augstein A. Jarabo M.-E.P. Barthel P. Mangner N. Labeit S. Bowen T.S. Linke A. Modulation of Titin and Contraction-Regulating Proteins in a Rat Model of Heart Failure with Preserved Ejection Fraction: Limb vs. Diaphragmatic Muscle Int. J. Mol. Sci.202425661810.3390/ijms 2512661838928324 PMC 11203682 · doi ↗ · pubmed ↗
- 6Goto K. Schauer A. Augstein A. Methawasin M. Granzier H. Halle M. Van Craenenbroeck E.M. Rolim N. Gielen S. Pieske B. Muscular changes in animal models of heart failure with preserved ejection fraction: What comes closest to the patient?ESC Heart Fail.2021813915010.1002/ehf 2.1314233350094 PMC 7835579 · doi ↗ · pubmed ↗
- 7Koser F. Hobbach A.J. Abdellatif M. Herbst V. Türk C. Reinecke H. Krüger M. Sedej S. Linke W.A. Acetylation and phosphorylation changes to cardiac proteins in experimental H Fp EF due to metabolic risk reveal targets for treatment Life Sci.202230912099810.1016/j.lfs.2022.12099836179815 · doi ↗ · pubmed ↗
- 8Mc Pherron A.C. Lawler A.M. Lee S.J. Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member Nature 1997387839010.1038/387083 a 09139826 · doi ↗ · pubmed ↗