The Nuclear Transcription Factor SlNF-YC9 Regulates the Protrusion of Tomato Fruit Tip
Zihan Gao, Ting Long, Pengyu Guo, Junjie Luo, Xiaoqian Nie, Qiaoli Xie, Guoping Chen, Zongli Hu

TL;DR
This study shows that the SlNF-YC9 gene in tomatoes affects fruit shape by regulating genes involved in auxin transport and response.
Contribution
The study identifies SlNF-YC9 as a key regulator of tomato fruit tip protrusion through NF-Y complex formation.
Findings
CRISPR/Cas9 mutants of SlNF-YC9 showed a prominent fruit tip protrusion compared to wild type.
SlNF-YC9 represses FUL2 expression by forming a trimer that binds its promoter.
Auxin-related genes PIN4, PIN5, PIN9, ARF7, and LAX3 were upregulated in SlNF-YC9 mutants.
Abstract
NF-Y transcriptional regulators play crucial roles in diverse biological processes in plants, primarily through the formation of NF-Y complexes that bind to specific DNA motifs. These complexes modulate the expression of downstream genes, which influence plant development and growth. In our research, the function of the NF-Y family C subunit member SlNF-YC9 gene in tomato was investigated with the CRISPR/Cas9 method. In contrast to the WT (wild type), the mutant CR-SlNF-YC9 exhibited a prominent protrusion at the fruit tip. The quantitative PCR analysis displayed that the transcription levels of genes associated with auxin transport (PIN4, PIN5, and PIN9) as well as auxin response genes (ARF7 and LAX3) were enhanced in the CR-SlNF-YC9 fruits than in the WT. Analysis of dual-luciferase reporter and EMSA assays showed that the SlNF-YC9-YB13b-YA7a trimer specifically binds the FUL2…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
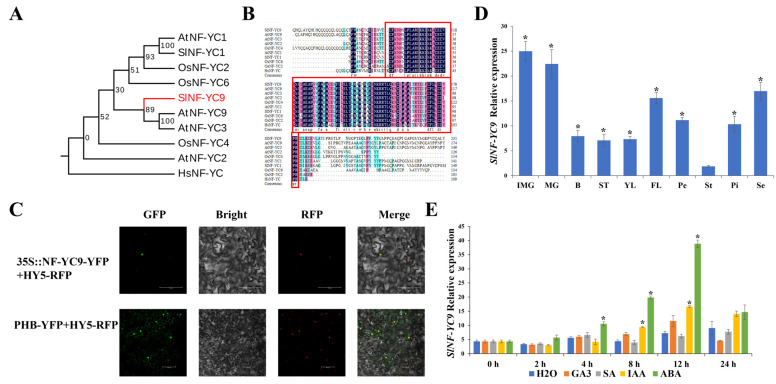 Figure 1
Figure 1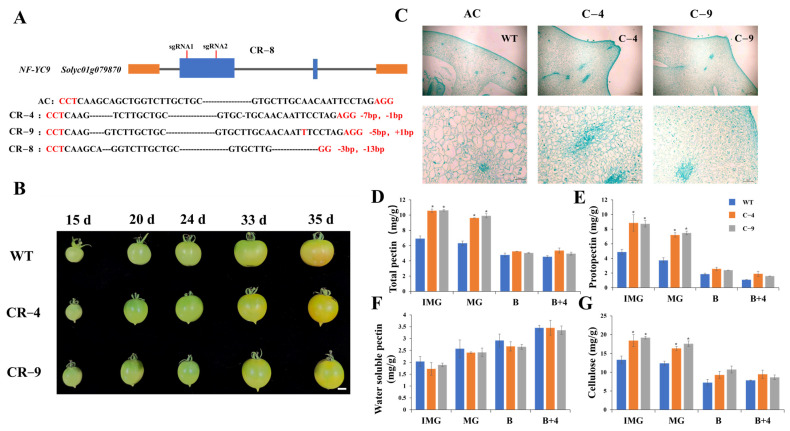 Figure 2
Figure 2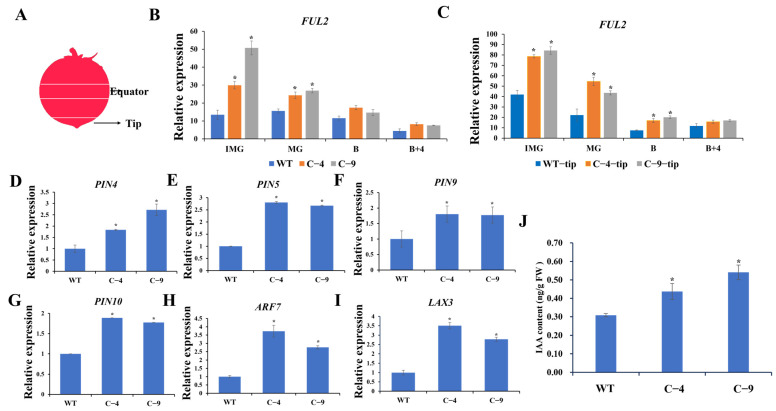 Figure 3
Figure 3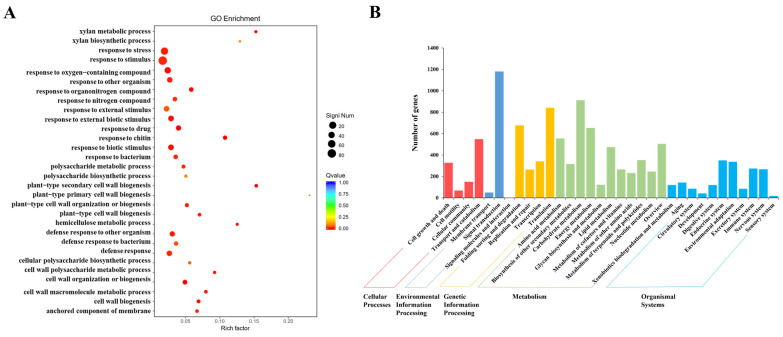 Figure 4
Figure 4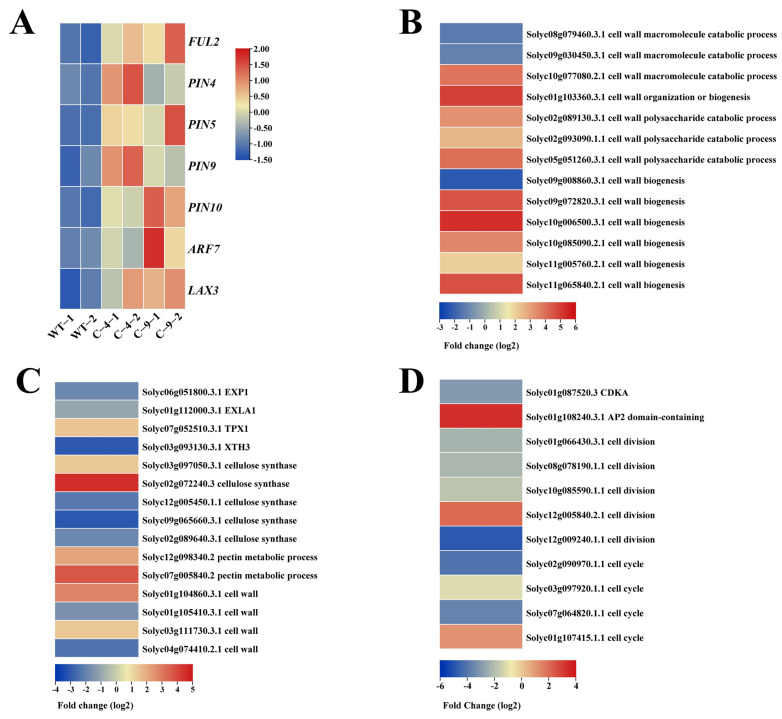 Figure 5
Figure 5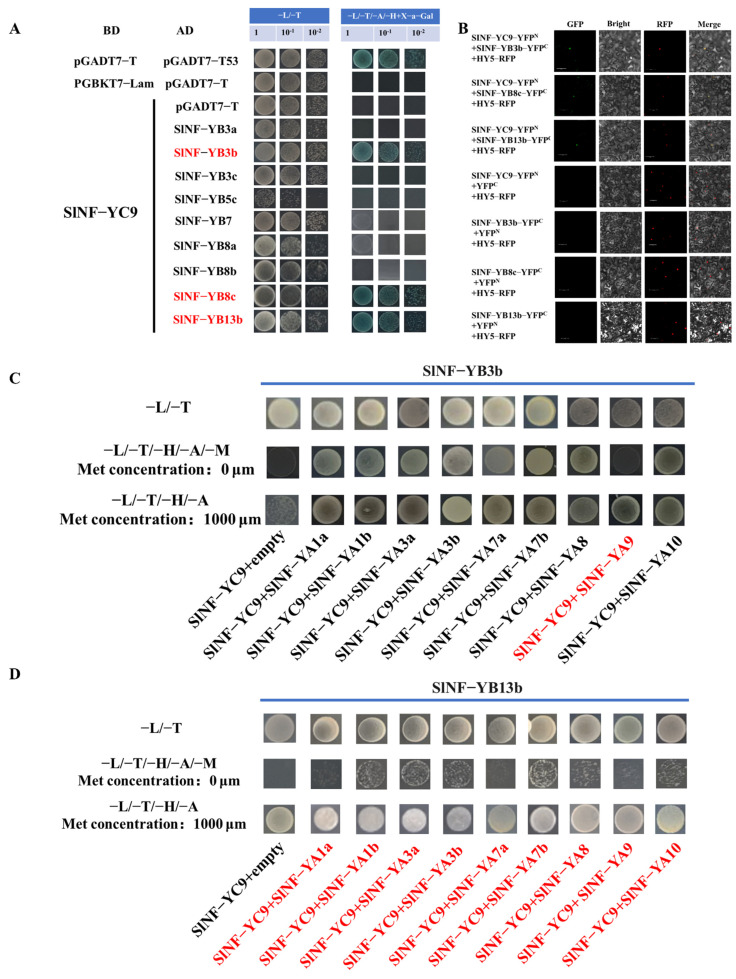 Figure 6
Figure 6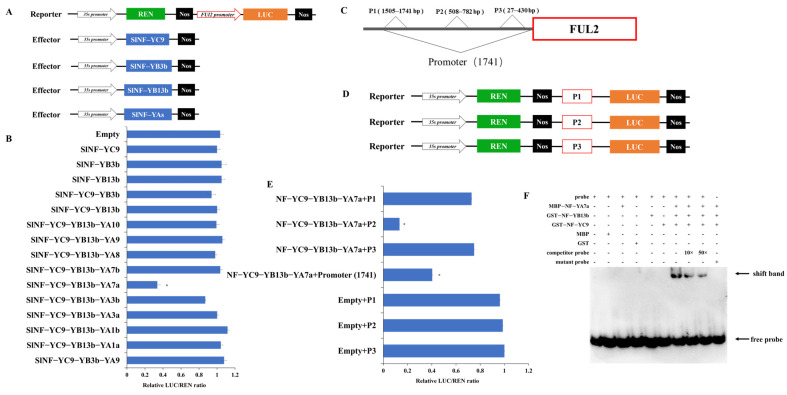 Figure 7
Figure 7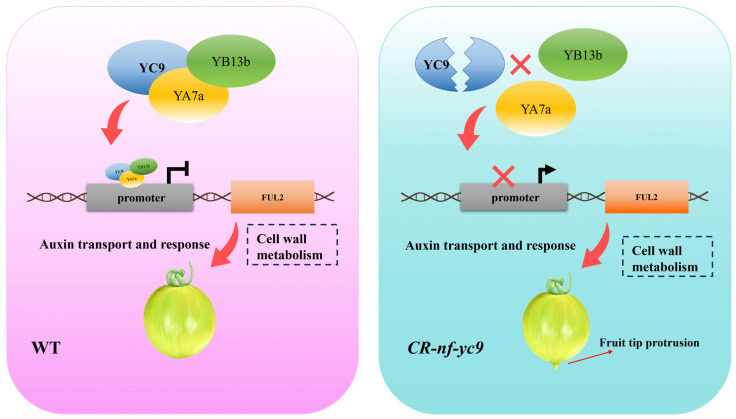 Figure 8
Figure 8- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Reproductive Biology · Plant Gene Expression Analysis
1. Introduction
The tomato (Solanum lycopersicum) is a vital global crop and serves as a model plant for studying fleshy fruits due to its relatively small genome and established genetic transformation system [1,2]. The shape of the tomato fruit is a key factor influencing fruit quality. Tomatoes have developed diverse fruit sizes and shapes through the process of domestication and improvement, including round, heart, long, flat, rectangular, ellipsoid, and obovoid [3]. The locule number (lc) and the fasciated (fas) locus regulate the number of locules in tomato and can directly affect fruit shape and size [4,5]. SUN is a key gene controlling the elongated morphology of tomato fruits, and the enhanced transcript level of the SUN delivered an effect on the length of tomato fruits [6]. A single mutation in a premature termination codon in the OVATE gene results in a transition from round- to pear-shaped fruit [7]. Notably, SUN caused an increase by lengthening the number of cells along the entire proximal-distal axis, whereas OVATE caused fruit elongation in the proximal region [8]. In heart-shaped fruits, the proximal end is longer than the distal end, resulting in a distinct tip protrusion at the distal end [9].
Previously, there have been relatively few studies on the protuberance of pointed tips in tomato fruits. Overexpression of the SlFUL2 (FRUITFULL2), a MADS-box family transcription factor, induces the formation of tomato fruit pointed tips that may be related to the abnormal style abscission [10]. The loss of function of PT (POINTED TIP), encoding a C2H2-type zinc finger protein transcription factor, exhibited the promoted formation of pointed-tip fruits by influencing the mRNA abundance of FUL2 [11]. Recent studies have revealed that an organelle RNA recognition motif (RRM) protein also plays a role in tomato fruit morphology. CR-slorrm2 exhibits fruit pointed tips at the distal end as well as defective style and ovary development [12]. In addition, the fruit tip protrusion phenotype is observed in both tomato silencing mutants of the gene encoding type B heterotrimeric G protein γ-subunit (SlGGB1) [13] and in pyrobactin resistance-like 9 (PYL9) silencing mutants [14]. However, the deeper mechanisms underlying the production of fruit tip protrusion are still unclear and need to be explored in further research.
NF-Y (Nuclear transcription factor Y), also known as HAP (Heme-activating protein) or CBFD (CCAAT binding factor domain), is a class of transcription factors widely distributed in both animals and plants [15,16]. NF-Y specifically recognizes and binds to the CCAAT-box within the promoter region of target genes, thereby regulating their transcription [17,18]. NF-Y consists of three distinct subunits: NF-YA (also known as CBF-B or HAP2), NF-YB (CBF-A or HAP3), and NF-YC (CBF-C or HAP5) [19]. NF-YA, NF-YB, and NF-YC subunits typically form large heterotrimeric complexes that function to bind DNA and regulate gene expression [20,21]. During the formation of the NF-Y complex in yeast and animals, NF-YB and NF-YC first form a heterodimer in the cytoplasm and then translocate to the nucleus, where the NF-YA subunits are subsequently recruited to generate the complete complex [22,23]. In yeast and mammals, each subunit of NF-Y is encoded by one (at most two) gene [18]. In contrast, the NF-Y subunits in plants show a familial expansion and are generally encoded by multiple genes encoding the same class of subunits, which creates additional challenges for the study of NF-Y family genes in plants [17,24,25,26,27,28].
Previous studies have shown that NF-Y transcription factors can be extensively involved in plant growth and development [29,30,31,32,33,34,35,36,37]. NF-Y trimeric complexes play crucial regulatory roles across plant species. In Arabidopsis, NF-YA/B/C subunits can form various nuclear complexes [25], while the legume MtNF-A1/2-B16-C1/2 trimer controls nodule formation [38]. Similarly, cassava possesses the MeNF-YA1/3-B11/16-C11/12 trimeric complex [39]. Moreover, studies have revealed that NF-Y family genes also play essential roles in regulating plant responses to stress [40,41,42] and photomorphogenesis [43,44]. In tomato, the NF-Y complex consisting of the NF-YB8c-YC1e dimer and NF-YA9, positively regulates the expression of the SlCHS1 gene, thereby modulating fruit development [45].
So far, in plants, many studies about NF-Y genes have been carried out by using Arabidopsis thaliana as a model plant, and most research has focused on NF-YA and NF-YB genes; however, the function of NF-YC is not well understood. Hence, it is crucial to investigate the functions of the NF-YC genes of tomatoes. In this study, a SlNF-YC9 knockout vector was constructed and introduced into wild-type tomato using CRISPR/Cas9 technology to produce homozygous SlNF-YC9 knockout plants. Our data indicated that CR-nf-yc9 fruits exhibit the tip protrusion phenotype. In addition, this study revealed that SlNF-YC9 can participate in forming 10 distinct NF-YA-YB-YC9 trimeric complexes. Among them, the SlNF-YC9-YB13b-YA7a trimeric complex was found to suppress the expression of the SlFUL2 gene. The research elucidates the molecular regulatory mechanisms of SlNF-YC9 in tomato and fruit apex development, enriches the understanding of the biological functions and pathways of the tomato NF-Y gene family, and provides a theoretical foundation for tomato variety improvement.
2. Results
2.1. Bioinformatics Analysis, Subcellular Localization, and Expression Pattern of SlNF-YC9
Previous studies have led to the identification of numerous genes in tomato, which have been classified and named based on their amino acid sequence homology [24,45]. We performed a phylogenetic analysis of NF-Y genes in tomato based on available genomic data and the exclusion of several NF-Y genes with highly repetitive sequences. The evolutionary tree results indicated 9 NF-YA genes, 25 NF-YB genes, and 15 NF-YC genes in tomato (Figure S1). Based on the tomato transcript expression data (http://ted.bti.cornell.edu/, accessed on 14 August 2022), NF-YC9, a gene exhibiting relatively high expression levels across various tissues, was selected from the NF-YC gene family as the focus of the study (Figure S2). Phylogenetic tree analysis of SlNF-YC9 homologous proteins using MEGAX showed that SlNF-YC9 has the highest homology with Arabidopsis NF-YC9 and NF-YC3 (Figure 1A). Multiple protein sequence alignments showed that NF-YC9 proteins are highly conserved across species, all having CBF structural domains (Figure 1B).
To analyze the subcellular localization of the SlNF-YC9 protein, 35S::SlNF-YC9-YFP fusion vectors were constructed and co-injected with a nuclear localization marker for transient expression in tobacco leaves. The results showed that the green fluorescence emitted by SlNF-YC9-YFP precisely colocalized with the red fluorescence of the nuclear localization marker, suggesting that SlNF-YC9 proteins are predominantly localized in the nucleus (Figure 1C). To further investigate the function of SlNF-YC9 in tomato growth and development, we used quantitative real-time PCR (qRT-PCR) to detect the expression levels of SlNF-YC9 in different tissues and organs of tomato. The results revealed that the transcript levels of SlNF-YC9 were highest in IMG and MG fruits, followed by flowers, while the lowest expression was found in the stamens of the four floral organs (Figure 1D). Additionally, we analyzed the transcript accumulation of SlNF-YC9 in seedlings in response to different hormone treatments. The results demonstrated that the expression of SlNF-YC9 was significantly induced within 8–12 h after IAA and ABA treatments, followed by a rapid decrease (Figure 1E).
2.2. The Knockout of SlNF-YC9 Alters Fruit Morphology in Tomato
To assess the biological function of SlNF-YC9 in tomato, the CRISPR/Cas9 system was used to generate transgenic tomato plants with SlNF-YC9 knockout mutants. Two sgRNAs were employed to target the first exon of the SlNF-YC9 gene for knockout, thereby increasing the knockout success rate. Subsequently, we successfully generated three knockout transgenic lines (CR-SlNF-YC9), designated CR-4, CR-8, and CR-9. At the first site of CR-4, there is a 7-bp deletion, and at the second site, a 1-bp deletion; at the first site of CR-9, there is a 5-bp deletion, and at the second site, a 1-bp insertion (Figure 3A). Both of these mutations cause a frameshift, which disrupts the protein function of SlNF-YC9. However, at the first site of CR-8, there is a 3-bp deletion, which does not lead to a frameshift mutation. Therefore, we chose CR-4 and CR-9 for subsequent experiments.
Subsequently, we observed that CR-SlNF-YC9 exhibited a noticeable protrusion at the fruit distal end compared to the wild type (WT) (Figure 2B). Moreover, anatomical analysis of the CR-SlNF-YC9 fruits was performed using paraffin-embedded sections. The results revealed that there was no obvious shedding zone in the pointed tips of the CR-SlNF-YC9 fruits, and notably denser cell arrangement and smaller cell size were observed in CR-SlNF-YC9 compared to the WT (Figure 2C). Furthermore, the pectin and cellulose content in fruits at different developmental stages of CR-SlNF-YC9 were measured. (Figure 2D–G). The results showed that the protopectin, total pectin, and cellulose contents in the knockout lines were significantly higher than those in the WT during the IMG and MG stages (Figure 2D,E,G), while the water-soluble pectin content showed no significant changes (Figure 2F). Furthermore, the qRT-PCR results showed that there were corresponding changes in the CR-SlNF-YC9 mutants in genes involved in cell expansion (EXP1), cell wall structure modification (XTH2 and XTH3), and cell cycle regulatory factors (Cyc B1;1 and CycD3;1) (Figure S3). Taken together, these results provide evidence that knockout of SlNF-YC9 can affect the protrusion formation of tomato fruit tips.
2.3. The Knockout of SlNF-YC9 Affects the Expression Levels of the FUL2 Gene in Tomato Fruits
To investigate the regulatory mechanism of the SlNF-YC9 gene in fruit tip projection, the expression of several fruit morphogenesis genes in WT and CR-SlNF-YC9 fruits was analyzed. It was found that among these genes, only the expression levels of FUL2 were significantly changed (Figure 3B), while the transcript accumulation of the other genes (SUN, OVATE, PT, and SlORRM2) involved in the development of fruit shape had no significant changes compared to the WT (Figure S4). Moreover, the mRNA abundance of FUL2 at different developmental stages and in different fruit parts was examined. According to our results, the transcript levels of FUL2 gradually decreased along with fruit ripening, and the expression levels of FUL2 were significantly up-regulated in CR-SlNF-YC9 at the IMG period compared with the WT (Figure 3B,C). In addition, the mRNA levels of FUL2 were higher in the fruit tip than in the fruit equator at all four fruit periods, suggesting that SlNF-YC9 primarily affects the protrusion of the fruit tip at the distal end of the fruit, rather than the entire fruit.
2.4. The Knockout of SlNF-YC9 Affects the IAA Content and Expression of Auxin-Related Genes in Tomato Fruits
Previous studies have demonstrated that the development of the fruit tips in FUL2-OE was regulated through the auxin pathway [45]. Therefore, the expression levels of auxin-related genes in CR-SlNF-YC9 and WT fruits were examined. The qRT-PCR results showed that the transcript levels of PIN4, PIN5, PIN9, and PIN10 genes, which are crucial for auxin transport, in the fruit tips of CR-SlNF-YC9 were significantly higher than those in the WT (Figure 3D–G), whereas FZY1, FZY2, FZY5, and FZY6 genes associated with IAA synthesis pathways showed no significant differences when compared to the WT (Figure S5). To validate these results, we quantified endogenous IAA levels and observed that the IAA content in fruit tips of CR-SlNF-YC9 was higher than the WT (Figure 3J). Based on the results of the IAA-induced expression profile (Figure 1E), the IAA-responsive genes were also analyzed. The qRT-PCR results showed that ARF7 and LAX3 were significantly upregulated in CR-SlNF-YC9 compared to the WT (Figure 3H,I).
2.5. RNA Sequencing Analysis of CR-SlNF-YC9 Fruit Tips
To enhance the investigation of the molecular mechanism of tomato fruit tips development, a transcriptome analysis of the WT and CR-SlNF-YC9 fruit tips was performed. A total of 959 differentially expressed genes were identified in the knockout mutant compared to the WT, of which 406 were upregulated and 553 were downregulated (Figure S6). To further categorize the differentially expressed genes, we performed Gene Ontology (GO) enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway classification on all differentially expressed genes. Functional annotation analysis showed that knockout of SlNF-YC9 was involved in cell wall biosynthesis, metabolism, auxin polar transport, and regulation of stimulus responses (Figure 4A). In terms of biological processes, the differentially expressed genes were mainly distributed in metabolic and cellular processes, and in terms of molecular functions, the differentially expressed genes were mainly distributed in the areas of catalytic activity and binding activity (Figure 4B). To further validate our results, we focused on growth hormone pathway-related genes and found that the RNA-seq data were consistent with our quantitative results (Figure 5A). Given the more densely arranged and smaller cell morphology of CR-SlNF-YC9 fruit tip cells (Figure 2C), relevant genes involved in cell division, cell cycle, and cell wall organization processes were also focused on. Transcript levels of genes related to cell wall catabolism, such as expansin (EXP), xyloglucan endotransglucosylase hydrolase (XTH), cellulase synthetase, and pectin methyl esterase (PME), were relatively significantly altered in the mutants (Figure 5B,C). Furthermore, the expression of genes related to cell wall biosynthesis, cell division, and cell cycle also changed significantly in the mutants (Figure 5D). These results suggest that SlNF-YC9 may further influence the protrusion of fruit tip formation through the auxin and cell wall pathway.
2.6. SlNF-YC9 Can Form Multiple Trimeric Complexes with SlNF-YBs and SlNF-YAs
The NF-YB and NF-YC subunits typically form dimers, which then form heterotrimeric complexes upon recruitment of NF-YA, which then recognize the CCAAT element that binds to the promoter of the target gene, thereby regulating the transcription of the target gene [16,25]. Therefore, we speculate that it is also possible that the formation of the fruit tip projection is further regulated in CR-SlNF-YC9 by the formation of the NF-Y complex. Nine SlNF-YBs proteins with relatively high expression in tomato fruits were selected by tomato transcript expression data, followed by protein-protein interaction experiments to screen for SlNF-YBs proteins capable of forming SlNF-YB-YC9 complexes. These SlNF-YBs-AD were co-expressed with SlNF-YC9-BD in yeast cells, revealing that only SlNF-YB3b-AD, SlNF-YB8c-AD, and SlNF-YB13b-AD could be grown on a quadruple dropout medium (SD/-Leu-Trp-His-Ade+X-a-gal), similar to the yeast cells harboring pGADT7-T and pGBKT7-53 (positive control) (Figure 6A). The BiFC assay results showed that stronger fluorescent signals were detected in the epidermal cell nucleus of N. benthamiana leaves only when SlNF-YC9-YFPN was co-expressed with SlNF-YB3b-YFPC, SlNF-YB8c-YFPC, and SlNF-YB13b-YFPC (Figure 6B). These results demonstrate that SlNF-YC9 was able to interact with SlNF-YB3b, SlNF-YB8c, and SlNF-YB13b to form dimers.
After screening for three different SlNF-YB proteins that form dimers with SlNF-YC9, further screening was performed to identify SlNF-YA subunits recruited by the NF-YB-YC dimer. These SlNF-YA proteins, obtained from our evolutionary tree analysis of NF-YA (Figure S1) in tomato, were subjected to yeast three-hybrid experiments with SlNF-YB3b, SlNF-YB8c, SlNF-YB13b, and SlNF-YC9, respectively (a total of 27 different combinations, Table S1). SlNF-YAs and SlNF-YC9 were inserted into the pBridge vector to construct the SlNF-YC9-YA expression vector, which was co-transformed with the SlNF-YBs-pGADT7 expression vector into yeast for Y2H progression. The yeast was then grown on double (SD/-Leu-Trp), quadruple (SD/-Leu-Trp-His-Ade), and quintuple (SD/-Leu-Trp-His-Ade-Met) dropout media. The yeast co-transformed with the SlNF-YC9-YB13b dimer and all SlNF-YAs grew normally on media with high concentrations of methionine but failed to grow in methionine-free media. However, the yeast co-transformed with SlNF-YC9-YB3b and SlNF-YA9 failed to grow in methionine-free media (Figure 6C,D). In addition, the yeast co-expressing the SlNF-YC9-YB8c dimer with all SlNF-YAs grew normally at different concentrations of methionine (Figure S7A, B). The results indicated that SlNF-YC9-YB3b forms a heterotrimer with SlNF-YA9, and NF-YC9-YB13b forms heterotrimers with SlNF-YA1a, SlNF-YA1b, SlNF-YA3a, SlNF-YA3b, SlNF-YA7a, SlNF-YA7b, SlNF-YA8, SlNF-YA9, and SlNF-YA10, respectively. While the SlNF-YC9-YB8c dimer cannot form any intact trimeric complexes with SlNF-YAs.
2.7. SlNF-YC9-YB13b-YA7a Trimeric Complex Represses the Promoter Activity of FUL2
The CR-SlNF-YC9 fruits exhibited similar phenotypes of fruit tip protrusion as the FUL2-OE fruits [10], and FUL2 was significantly upregulated in the CR-SlNF-YC9 fruits (Figure 2B). Previous studies have shown that the NF-Y complex can regulate the transcriptional activity of downstream target genes by binding to the CCAAT element in the promoter region [45]. Moreover, five CCAAT elements were found in the FUL2 promoter sequence (Table S2). Therefore, we hypothesized that the SlNF-YC9 alone, or SlNF-YC9-YB dimers, or SlNF-YC9-YB-YA heterotrimers may directly regulate the expression of the FUL2 gene. Dual-luciferase reporter assays were performed. The effector vectors containing the coding sequences of SlNF-YC9, SlNF-YB3b, SlNF-YB13b, and SlNF-Yas, respectively, and the reporter vector containing FUL2 promoter sequences (1741 bp upstream of the transcriptional start site) (Figure 7A), were transiently expressed in tobacco leaves via Agrobacterium-mediated infiltration, according to the different combinations shown in Figure 7B. Dual-luciferase assays revealed that only the complex formed by SlNF-YC9-YB13b-YA7a significantly inhibited FUL2 promoter activity in vivo (Figure 7A,B), whereas the other eight SlNF-YC9-YB13b-YAs trimeric complexes, the individual NF-Y proteins (SlNF-YC9, SlNF-YB3b, SlNF-YB13b), the SlNF-YC9-YB3b and SlNF-YC9-Y13b dimers did not affect the promoter activity of FUL2.
To verify the specific binding site of the SlNF-YC9-YB13b-YA7a complex to the FUL2 promoter, the reporter vectors were categorized into three types (Figure 7C,D): P1 includes sequences from 1505–1741 bp upstream of the transcription start site (containing two CCAAT binding sites); P2 includes sequences from 508–782 bp upstream of the transcription start site (containing one CCAAT binding site); P3 includes sequences from 27–430 bp upstream of the transcription start site (containing two CCAAT binding sites). The relative LUC/REN ratios in co-transformed tobacco leaves carrying the effectors SlNF-YC9, YB13b, YA7a, and the P2 reporter plasmid were significantly downregulated compared to the control (Figure 7E). To further confirm the direct binding of the SlNF-YC9-YB13b-YA7 trimer to the SlFUL2 promoter, this study performed EMSAs using biotin-labeled oligonucleotides containing the CCAAT motif. The results demonstrate that when a mixture of the three tomato transcription factor proteins (SlNF-YC9, SlNF-YB13b, and SlNF-YA7) was incubated with the biotin-labeled double-stranded DNA probe harboring the CCAAT cis-element, a clear mobility shift was observed (Figure 7F). The results suggest that the SlNF-YC9-YB13b-YA7a complex may inhibit the expression of the FUL2 gene by binding to the P2 region of the FUL2 promoter.
3. Discussion
The NF-Y family is an important class of multi-subunit nuclear transcription factors that are widely involved in the regulation of gene expression in plants. Members of the NF-Y family play a crucial role in various physiological processes during plant growth and development, including seed germination, root development, and flower formation [46,47]. In tomato, virus-induced gene silencing (VIGS) is used to suppress the NF-Y genes, and five NF-Y genes were found to affect fruit ripening [24]. In our study, the CR-nf-yc9 mutant showed no changes in fruit ripening but did show a phenotype in fruit tip development (Figure 2B). Some genes affecting fruit shape development, such as SUN, OVATE, and PT, showed no significant differences in expression levels in CR-SlNF-YC9 fruit tips compared to WT. (Figure S4). Notably, we found that the mRNA abundance of FUL2 in CR-nf-yc9 was significantly upregulated (Figure 3B,C), which corresponds to the past studies that overexpression of FUL2 can cause protrusion of tomato fruit tips [10,11]. Based on these results, we hypothesized that the regulatory mechanism of SlNF-YC9 in tomato fruit tip protrusion is likely to be connected to the FUL2 gene.
NF-Y family genes are widely found in plants and animals and play important roles as transcription factors in the regulation of gene expression. They usually consist of three subunits: NF-YA, NF-YB, and NF-YC [48]. The three subunits interact with each other, collectively recognizing and binding to specific DNA sequences, thereby regulating the transcriptional activity of target genes [18,49]. Previous studies have shown that the assembly process of NF-Y heterotrimer is strictly ordered. NF-YB and NF-YC assemble into dimers outside the nucleus via histone folding structural domains. Subsequently, inside the nucleus, the A1 (α-helix) of the NF-YA subunit tightly binds to the surface grooves to form the NF-Y heterotrimer [49]. The assembly of the NF-Y complex in plants follows the same mechanism. In tomato, the complete NF-Y complex consisting of NF-YB8c-YC1e dimer and NF-YA9 positively regulates the expression of the SlCHS1 gene and specifically binds to the CCAAT element upstream of the SlCHS1 gene promoter [45]. Moreover, NF-YA7 can interact with NF-YB2-YC4 dimer to form a heterotrimer, which in turn is involved in regulating the response of Brassica napus under drought stress [50]. In our study, we screened SlNF-YC9-YB3b-YA9, SlNF-YC9-YB13b-YA1a/1b/3a/3b/7a/7b/8/9/10, a total of 10 different trimeric complexes, by yeast two-hybrid, BiFC (Figure 6A,B), and three-hybrid experiments (Figure 6C,D). The CCAAT motif is a typical promoter element widely found in the promoter regions of many genes in eukaryotes [51,52,53]. The NF-Y complex recognizes and specifically binds to the sequence through its unique DNA-binding ability, thereby regulating the transcriptional expression of downstream genes [21,54,55]. In this research, dual luciferase experiments showed that neither SlNF-YC9 alone nor the complexes formed by SlNF-YC9 and SlNF-YBs could affect FUL2 transcription (Figure 7B). However, only the SlNF-YC9-YB13b-YA7a complex was able to inhibit FUL2 expression (Figure 7B,E). Our study provided insights into the many different forms of NF-Y family complexes in tomato, revealing the diversity and specificity of these complexes in the regulation of gene expression.
Plant hormones are pivotal in plant growth and development, and the NF-Y family also plays a critical role in the regulation of plant hormones. In Arabidopsis, the NF-YC3/4/9 genes can inhibit GA-regulated seed germination; at the protein level, they were able to interact with the DELLA protein RGL2 to inhibit ABI5 expression [56]. Overexpression of the CdtNF-YC1 gene in rice significantly enhances the expression of ABA response, synthesis, and signal genes and improves plant drought tolerance [41]. In our study, SlNF-YC9 affected the expression levels of several IAA transporter genes in fruit tips (Figure 3E–H). Additionally, the hormone expression pattern of SlNF-YC9 indicated its responsiveness to IAA (Figure 1E), and IAA-responsive genes were also significantly upregulated in CR-SlNF-YC9 (Figure 3I,J). Subsequently, significantly higher IAA content was detected in fruits of SlNF-YC9 knocked out compared to the WT (Figure 3D). The above results suggest that SlNF-YC9 plays an important function in auxin transport and response.
Cell wall metabolism is crucial in plant development, involving the synthesis, degradation, and remodeling of cell wall components, which directly affect plant structure and morphology. During tomato fruit development, pectin and cellulose play key roles. Their interaction regulates the mechanical properties of the cell wall, thereby influencing the fruit’s size, shape, firmness, and market quality [57,58,59,60,61,62]. In our analysis, CR-SlNF-YC9 fruits contained significantly higher levels of protopectin, total pectin, and cellulose than WT (Figure 2D,E,G). Additionally, a large number of changes in cell wall metabolism, pectin, and cellulase-related gene expression were detected in CR-nf-yc9 by RNA-seq (Figure 5B–D). Notably, the key gene EXP1 involved in cell expansion showed a significant decrease in CR-SlNF-YC9, corresponding to which paraffin-embedded experiments revealed that knockout of SlNF-YC9 reduced cell size in tomato fruit tips. (Figure 2C). The above results suggest that SlNF-YC9 may affect the shape of tomato fruits by influencing cell expansion and pectin and cellulose metabolism, but further experiments are needed to verify this.
Taken together, our study shows that a rather pronounced bulging of the fruit tip occurs in CR-nf-yc9. The mechanistic analysis suggests that the knockout of SlNF-YC9 prevents the formation of the complete SlNF-YC9-YB13b-YA7 complex in tomato plants, thereby relieving the repressive effect of the SlNF-Y complex on the expression of the FUL2 gene. This enhances the expression of the FUL2 gene and subsequently promotes the formation of the fruit tip protrusion by the auxin signaling pathway. In addition, SlNF-YC9 may also affect fruit development through the cell expansion, cell wall metabolism pathway (Figure 8). In conclusion, our study enriches the regulatory network of tomato fruit shape and also further uncovers the biological functions of SlNF-Y transcription factors in plants.
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
The study involved wild-type tomato (Solanum lycopersicum Mill cv. Ailsa Craig, AC++), CR-SlNF-YC9 transgenic plants, along with Nicotiana benthamiana. The plants were grown under standard conditions: daytime temperature (26 °C), nighttime temperature (18 °C), 16 h of light and 8 h of darkness, 70% relative humidity, and a light intensity of 250 μmol m^−2^ s^−1^.
4.2. Bioinformatics Analysis
The sequence information of SlNF-YC9 was acquired from the SGN and NCBI websites, accessed on 2 October 2020. The evolutionary tree depicted in this article was generated using MEGA 6.06 and EVOLVIEW software V2. The DNAMAN 5.2.2 tool was employed to perform the alignment of several sequences between SlNF-YC9 and its family proteins. We analyzed the FUL2 promoter using plantCARE (Plant promoter Database, accessed on 2 June 2022).
4.3. Subcellular Localization Assay
The entire coding sequence of SlNF-YC9, with the termination codon removed, was amplified and fused with GFP to create the PHB-eGFP expression vector. The engineered vector was introduced into Agrobacterium GV3101, then transferred to tobacco via Agrobacterium-mediated genetic transformation. Following a 2-day period of darkness treatment, the fluorescence signal was detected using a confocal microscope. HY5-RFP was used as a nuclear localization marker [63]. The primers utilized for constructing the vector are displayed in Table S3.
4.4. SlNF-YC9 Knockout Vector Construction and Plant Transformation
The CRRISPR-GE (http://skl.scau.edu.cn/home/, accessed on 1 October 2020) website was used to select a knockout target on the first exon sequence of SlNF-YC9, and then construct the CRISPR/Cas9-SlNF-YC9 vector. The vector was introduced into the Agrobacterium strain LBA4404 and then transferred to the wild-type tomato by the Agrobacterium-mediated genetic transformation method. Transgenic plants were screened on a resistant medium containing kanamycin at a concentration of 50 mg L^−1^. Subsequently, the gene sequence of SlNF-YC9 was analyzed through sequencing to identify the specific type of knockout. The homozygous knockout transgenic lines were used for further investigation. The primers used for vector construction are shown in Table S3.
4.5. Measurement of Pectin and Cellulase Content
Determination of pectin content: The samples (Fruit tips of 20 days post-anthesis) were ground in liquid nitrogen. A 0.2 g portion of material was weighed, mixed with 1 mL of 95% ethanol, and placed in a drying oven at 95 °C for 30 min. After cooling to room temperature, the mixture was centrifuged at 8000 rpm for 5 min, and then the supernatant was discarded. Then, 1 mL of water was added to the precipitate, and the mixture was placed in a 50 °C water bath for 30 min. After cooling, the mixture was centrifuged at 8000 rpm for 5 min, and the supernatant was discarded (the supernatant can be used for the determination of water-soluble pectin). Next, 1 mL of extract was added to the precipitate, mixed well, and placed in a 95 °C water bath for 1 h. After cooling, the mixture was centrifuged at 8000 rpm for 10 min, and the supernatant was collected for the determination of original pectin. The formula is as follows: pectin content (mg/g) = (C × V) × ΔA2_530_/ΔA1_530_/(W × V1/V2). The formula ΔA1_530_ represents the difference between the absorbance values of the standard tube and the blank tube at 530 nm; ΔA2_530_ represents the difference between the absorbance values of the measurement tube and the control tube at 530 nm; C is the standard sample concentration (mg/mL); V is the volume of the standard added to the reaction system (mL); V1 is the volume of the sample added to the reaction system (mL); V2 is the volume of extract (mL); W is the sample’s fresh weight (g).
Determination of cellulase content using the Cellulose Extraction kit (Michy Biology, Suzhou, China). The samples were ground in liquid nitrogen, 0.1 g of material was weighed, mixed with 1 mL of 80% ethanol, placed in an 80 °C water bath for 30 min, cooled down, and then centrifuged at 8000 rpm for 5 min, with the supernatant being discarded. Then 1 mL of acetone was added, vortexed for 2 min, centrifuged at 8000 rpm for 5 min, and the supernatant was discarded. Then 1 mL of reagent I was added, vortexed and shaken for 2 min, soaked for 15 h, and then centrifuged at 8000 rpm for 5 min, and the supernatant was discarded. After washing twice with distilled water, the precipitate was dried. Finally, 1 mL of reagent III was added to the precipitate and mixed well, then centrifuged at 10,000 rpm for 10 min, and the supernatant was taken to measure the absorbance at 620 nm. The free cellulase content was calculated using the following formula: Cellulase content (mg/g) = [(ΔA_620_ − 0.0194)/(2.8737 × V1)]/(W × V1/V2) × Dilution Factor, where ΔA_620_ is the difference between the absorbance values of the measurement tube and the blank tube at 530 nm; V1 is the volume of samples (mL); V2 is the volume of extract (mL); W is sample fresh weight (g).
4.6. Fruit Tip Tissue Sections
Fruit tips of WT and CR-SlNF-YC9 were collected 15 days post-anthesis, treated with 1% hydrochloric acid for 10 min, and then placed in 70% FAA fixative for 24 h and embedded in Paraplast Plus (Sigma, Saint Louis, Missouri, USA). Longitudinal sections of the fruit tips were stained briefly with 0.04% (w/v) toluidine blue solution. After that, the samples were examined with a dissecting microscope (Nikon E100).
4.7. qRT-PCR Analysis and RNA-Sequencing
Total RNA was extracted from samples (Fruit tips of 20 days post-anthesis) using the Takara RNAiso Plus reagent. The M-MLV Reverse Transcriptase kit (Promega, Beijing, China) was used to convert RNA samples into cDNA. According to the manufacturer’s instructions, the transcription levels of specific genes were quantitatively analyzed using the SYBR Premix Ex Taq II kit (Takara, Ōtsu, Shiga, Japan) and gene-specific primers, using SlCAC (Solyc08g006960) as an internal reference gene. Quantitative reverse-transcription-PCR (qRT-PCR) was carried out using the CFX Connect Real-Time System (Bio-Rad, Hercules, California, USA). The analysis of relative expression levels was conducted using the 2^−ΔΔCT^ method. Primers used in reverse transcription and qPCR are listed in Table S3.
4.8. RNA Sequencing
Fruits of the CR-SlNF-YC9 and WT were harvested 15 days after anthesis. The apices were collected from the distal end of the fruit and immediately frozen in liquid nitrogen. Samples were collected in three biological replicates and sent to Sangon Biotech (Shanghai, China) for RNA-seq analysis. Differentially expressed genes (DEG) with more than twofold change in expression level with p value < 0.05 were screened out in Table S4.
4.9. Yeast Two-Hybrid and BiFC Assays
Yeast two-hybrid assays: The bait plasmid pGBKT7-SlNF-YC9 and the prey plasmid pGADT7-SlNF-YC3a, 3b, 3c, 5c, 7, 8a, 8b, 8c, and 13b were co-transformed into yeast strain Y2HGold. The transformed yeast was then plated on the SD medium lacking tryptophan and leucine, followed by incubation for 3 days. Thereafter, a single colony was selected and inoculated on the SD medium devoid of tryptophan, histidine, adenine, and leucine. This plate was incubated upside down for 1–2 days [64].
BiFC assays: The recombinant SlNF-YC9-YFPN, SlNF-YB3b-YFPC, SlNF-YB8c-YFPC, and SlNF-YB13b-YFPC vectors were transformed into the Agrobacterium tumefaciens GV3101. Then, SlNF-YC9-YFPN, SlNF-YB3b-YFPC, SlNF-YB8c-YFPC, SlNF-YB13b-YFPC, and control HY5-RFP were infiltrated into 4-week-old tobacco leaves by Agrobacterium-mediated genetic transformation. The subsequent experimental steps refer to the methods in the subcellular localization assay [65]. The primers used to construct the vector are shown in Table S3.
4.10. Yeast Three-Hybrid
Based on the principles of yeast two-hybrid experiments, SlNF-YC9 was inserted into the MCS I site (Bait site) of the pBridge vector, and SlNF-YAs were inserted into the MCS II site (Bridge site, Met-sensitive site) of the pBridge vector. SlNF-YBs were inserted into the pGADT7 vector. The constructed pGADT7 and pBridge plasmids were then co-transfected into the Y2HGold yeast. The transformed yeast cells were cultured on double (SD/-Leu-Trp), quadruple (SD/-Leu-Trp-His-Ade), and quintuple (SD/-Leu-Trp-His-Ade-Met) dropout media. The plates were incubated upside down for 1–2 days. The primers used to construct the vectors are shown in Table S3.
4.11. Transient Expression Assays
The coding sequences of SlNF-YC9 and SlNF-YB3b and SlNF-YB13b and SlNF-YA 1a, 1b, 3a, 3b, 7a, 7b, 8, 9, 10, were introduced into pGreenII 62-SK for effector, and promoter fragments of FUL2 were cloned into the pGreenII 0800-LUC vector as reporters, respectively. After the introduction of the recombinant plasmid into Agrobacterium GV3101, the recombinant plasmid was infiltrated into tobacco leaves. Post-infiltration with Agrobacterium, the leaves were stored in the dark for 1 day, followed by 2 days under light. The activities of LUC and REN were determined by using an enzyme marker according to the instructions of the Promega Dual-luciferase assay kit, and the ratio of LUC/REN was calculated [66].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Schultink A. Qi T. Bally J. Staskawicz B. Using forward genetics in Nicotiana benthamiana to uncover the immune signaling pathway mediating recognition of the Xanthomonas perforans effector Xop J 4New Phytol.20192211001100910.1111/nph.1541130156705 · doi ↗ · pubmed ↗
- 2Klee H.J. Giovannoni J.J. Genetics and control of tomato fruit ripening and quality attributes Annu. Rev. Genet.201145415910.1146/annurev-genet-110410-13250722060040 · doi ↗ · pubmed ↗
- 3Rodriguez G.R. Munos S. Anderson C. Sim S.C. Michel A. Causse M. Gardener B.B. Francis D. van der Knaap E. Distribution of SUN, OVATE, LC, and FAS in the tomato germplasm and the relationship to fruit shape diversity Plant Physiol.201115627528510.1104/pp.110.16757721441384 PMC 3091046 · doi ↗ · pubmed ↗
- 4Munos S. Ranc N. Botton E. Berard A. Rolland S. Duffe P. Carretero Y. Le Paslier M.C. Delalande C. Bouzayen M. Increase in tomato locule number is controlled by two single-nucleotide polymorphisms located near WUSCHEL Plant Physiol.20111562244225410.1104/pp.111.17399721673133 PMC 3149950 · doi ↗ · pubmed ↗
- 5Cong B. Barrero L.S. Tanksley S.D. Regulatory change in YABBY-like transcription factor led to evolution of extreme fruit size during tomato domestication Nat. Genet.20084080080410.1038/ng.14418469814 · doi ↗ · pubmed ↗
- 6Xiao H. Jiang N. Schaffner E. Stockinger E.J. van der Knaap E. A retrotransposon-mediated gene duplication underlies morphological variation of tomato fruit Science 20083191527153010.1126/science.115304018339939 · doi ↗ · pubmed ↗
- 7Liu J. Van Eck J. Cong B. Tanksley S.D. A new class of regulatory genes underlying the cause of pear-shaped tomato fruit Proc. Natl. Acad. Sci. USA 200299133021330610.1073/pnas.16248599912242331 PMC 130628 · doi ↗ · pubmed ↗
- 8Wang Y. Clevenger J.P. Illa-Berenguer E. Meulia T. van der Knaap E. Sun L. A Comparison of sun, ovate, fs 8.1 and Auxin Application on Tomato Fruit Shape and Gene Expression Plant Cell Physiol.2019601067108110.1093/pcp/pcz 02430753610 · doi ↗ · pubmed ↗