Gut Fungal Community Modulates Fat Deposition in Ningxiang Pigs: Species-Specific Regulation via the Glucose–SCFAs Metabolic Axis
Pengfei Huang, Hanmin Wang, Juan Wang, Zhenrong Qiu, Chunfeng Wang, Han Liu, Qiye Wang, Yali Li, Huansheng Yang

TL;DR
This study shows that gut fungi in Chinese pigs influence fat levels by affecting glucose and short-chain fatty acids, offering new strategies for pig breeding.
Contribution
The paper identifies a fungal-driven 'Glucose–SCFAs axis' as a novel mechanism for fat regulation in pigs.
Findings
NX pigs had higher fat and glucose but lower SCFAs compared to DLY pigs.
Aspergillus and Penicillium in NX pigs correlated with glucose and reduced SCFAs.
Rhodotorula in DLY pigs was linked to higher SCFAs and lipolysis.
Abstract
This study reveals a gut fungal-mediated mechanism regulating fat deposition in indigenous Chinese pigs. Compared to lean-type Duroc × Landrace × Yorkshire (DLY) pigs, high-fat Ningxiang (NX) pigs showed significantly higher fat deposition and serum glucose, but markedly lower colonic short-chain fatty acids (SCFAs). Distinct fungal communities were observed: NX pigs enriched with Aspergillus and Penicillium positively correlated with glucose but negatively with SCFAs, while DLY pigs’ dominant Rhodotorula strongly correlated with SCFAs. Metabolic analysis linked NX-enriched fungi to enhanced polysaccharide degradation and glucose bioavailability. We propose a fungal-driven “Glucose–SCFAs axis”: NX-associated fungi elevate glucose while suppressing SCFAs. Conversely, Rhodotorula in DLY enhances SCFA-induced lipolysis. Despite limited understanding of gut fungal roles in fat deposition…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
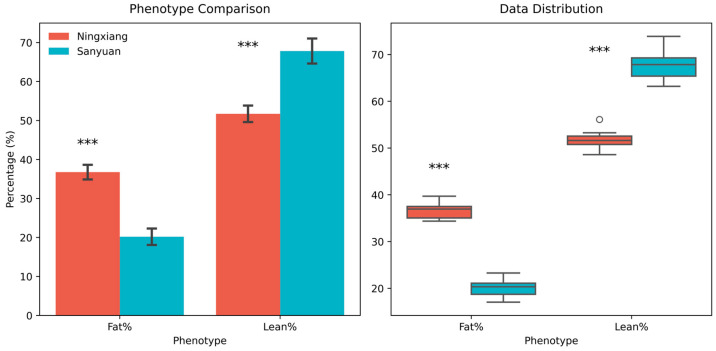 Figure 1
Figure 1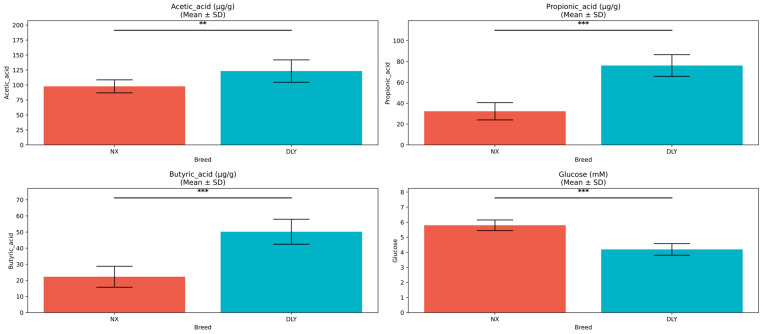 Figure 2
Figure 2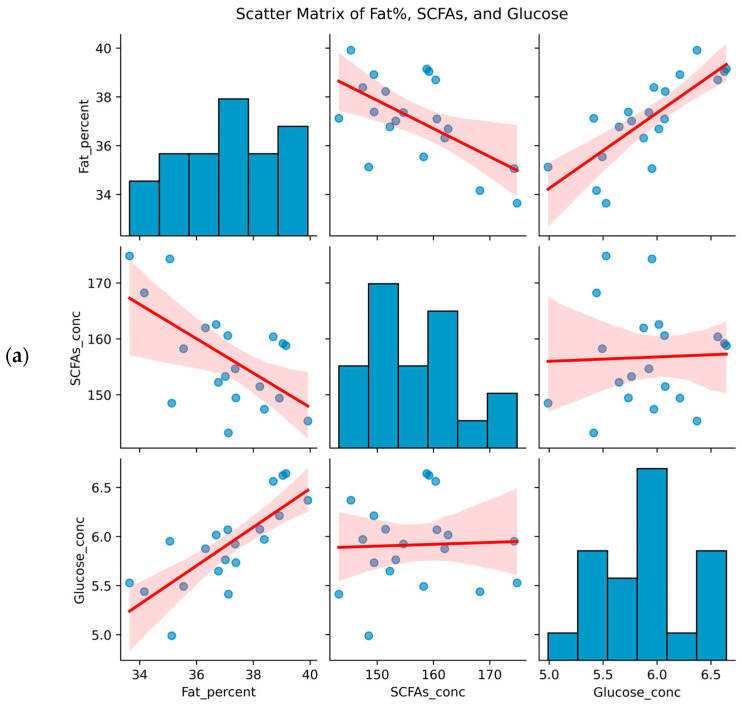 Figure 3
Figure 3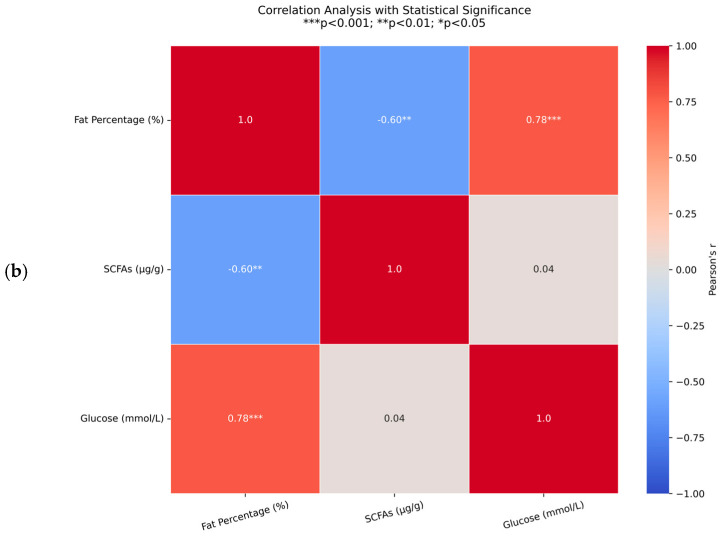 Figure 4
Figure 4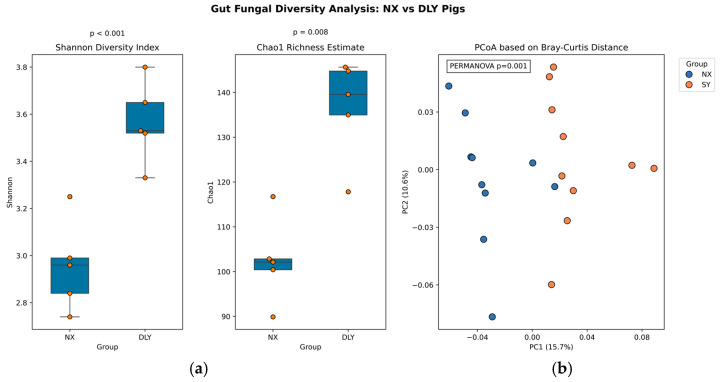 Figure 5
Figure 5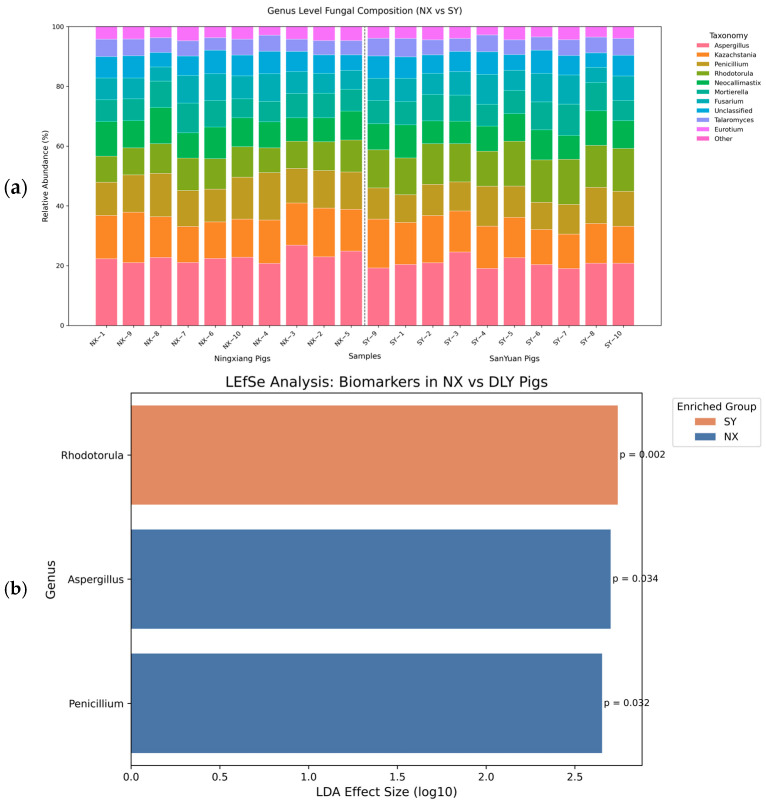 Figure 6
Figure 6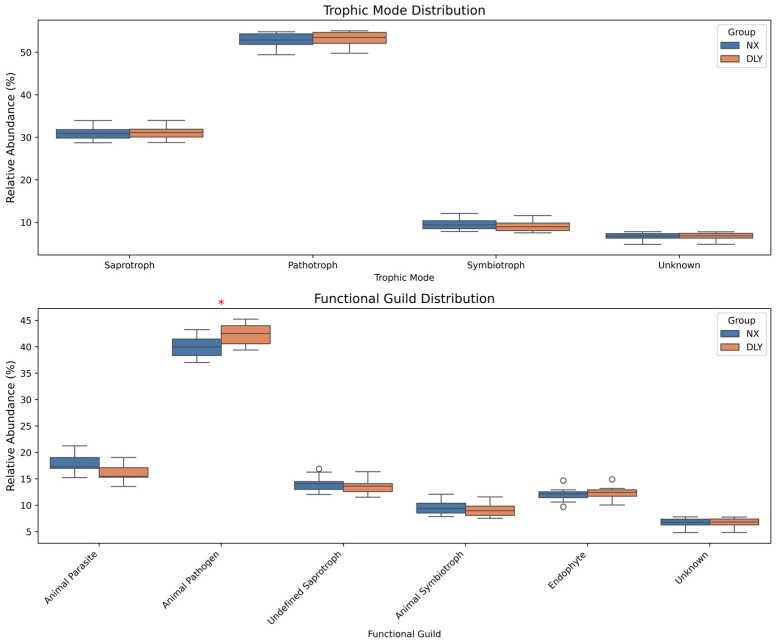 Figure 7
Figure 7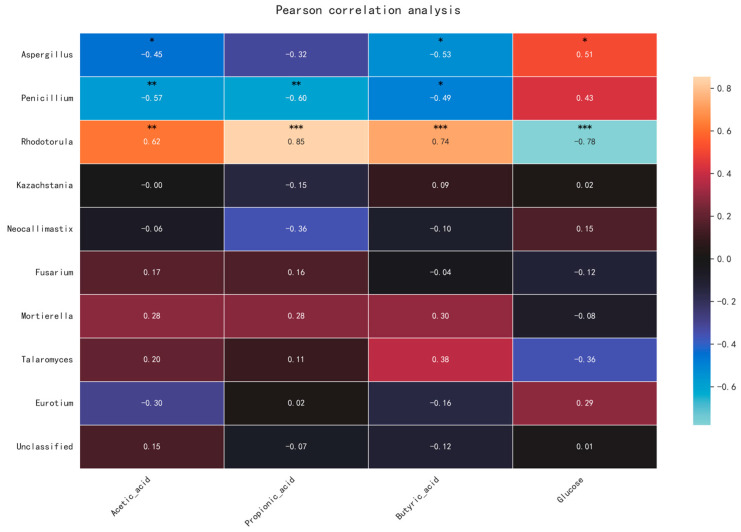 Figure 8
Figure 8- —Education Department of Hunan Province, China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Meat and Animal Product Quality · Fungal Biology and Applications
1. Introduction
Animal breeding significantly contributes to global economies, with pork production accounting for 35% of worldwide meat consumption [1]. The Ningxiang (NX) pig, as an important indigenous pig breed resource in China, is characterized by its remarkable high-fat phenotype. However, the regulatory mechanisms underlying its fat deposition remain incompletely elucidated. In striking contrast, Duroc × Landrace × Yorkshire (DLY) pigs exhibit outstanding lean meat percentage, providing an ideal comparative model for analyzing species differences in lipid metabolism.
Accumulating evidence highlights the critical role of gut microbial metabolites in host energy partitioning. Optimizing fat/lean ratios through microbial interventions could reduce feed costs while meeting consumer preferences for specialty pork [2]. Short-chain fatty acids (SCFAs) have been demonstrated to regulate systemic fat deposition [3,4]. Mechanistic studies reveal that butyrate suppresses fat accumulation by activating G protein-coupled receptor signaling pathways, effectively reducing high-fat diet-induced weight gain [5]. Conversely, propionate promotes lipid oxidative metabolism through PPARγ-dependent pathways, significantly reducing obesity risk [6,7]. Notably, monosaccharides derived from gut microbiota metabolism may influence lipid homeostasis via distinct mechanisms. Excessive glucose tends to be converted into fat storage [8,9], while the hepatic-specific metabolism of fructose markedly enhances lipogenesis and drives abnormal triglyceride accumulation [10].
Current research predominantly focuses on bacterial communities, with limited attention to fungal communities, which constitute only 0.1–5% of the gut microbiota. Emerging evidence suggests that fungal metabolic activities may indirectly modulate host metabolite levels through cross-kingdom interactions. For instance, Aspergillus enhance intestinal SCFA levels by promoting substrate hydrolysis through cellulase secretion [11,12], while Rhizopus elevate peripheral blood monosaccharide levels in hosts by decomposing plant-derived polysaccharides [13,14]. Notably, indigenous Chinese pig breeds like NX possess distinct fat deposition phenotypes, yet the role of gut fungi in regulating such traits remains unexplored. Previous studies on DLY pigs focused on bacterial SCFA producers (e.g., Firmicutes, Bacteroidetes) [15,16], but neglected fungal contributions. This gap limits holistic understanding of microbial regulation in swine lipid metabolism. Our study addresses this by comparatively analyzing fungal communities in high-fat (NX) and lean (DLY) pigs, hypothesizing that fungi modulate fat deposition via the glucose–SCFA balance.
Thus, this study aims to systematically characterize the gut fungal community structures of NX and DLY pigs to uncover their distinct roles in lipid metabolism. By correlating fungal taxa with host glucose and SCFA profiles, we identify key microbial determinants of fat deposition. Ultimately, we propose a novel regulatory paradigm—the gut fungal-mediated Glucose–SCFAs metabolic axis—to explain the divergent adipogenic phenotypes between these breeds.
2. Materials and Methods
2.1. Animal Feeding
Ten healthy NX pigs and ten DLY pigs of similar age and body weight (initial weight range: 70 kg ± 2.5 kg), with equal numbers of males and females, were selected. Sample size (n = 10/group) was determined by power analysis (α = 0.05, β = 0.8) using pilot data, detecting 25% diversity differences (G*Power v3.1). The experiment lasted 30 days. The 30-day duration was based on established gut microbiota stabilization periods in swine, ensuring detectable metabolic differences while minimizing age-related confounders. Pigs had ad libitum access to water and were fed an equal amount of a basal diet (Table 1, formulated according to NRC 2012) twice daily at 08:00 and 16:00, at a rate of 2.0 kg per pig per day.
2.2. Sample Collection
Pigs were fasted for 12 h prior to the end of the experiment. The following morning, 5 mL of blood was collected from the jugular vein. After standing for 30 min, samples were centrifuged at 3000× g for 10 min. The supernatant (serum) was aliquoted and stored at −80 °C. Pigs were stunned by electrical stunning followed by exsanguination. Mid-colon contents were aseptically collected, divided into two portions. One portion was mixed with 0.5% H_3_PO_4_, vortexed, and stored at 4 °C for SCFA analysis. The other portion was flash-frozen in liquid nitrogen and stored at −80 °C for fungal DNA extraction.
After slaughter, the head, feet, and viscera were removed, and carcass weight was recorded. Carcasses were split symmetrically along the spine. The left half-carcass was used for body composition analysis. Subcutaneous fat, intermuscular fat, and perirenal fat were dissected, pooled, and weighed (accuracy: 0.1 g). Skeletal muscle was separated, connective tissue and fat were removed, and the lean tissue was weighed.
2.3. Serum Glucose Analysis (HPLC)
Frozen serum samples were thawed and mixed at 4 °C. A 200 μL aliquot was added to 800 μL of acetonitrile to precipitate proteins, vortexed for 1 min, and centrifuged at 12,000× g for 15 min at 4 °C. The supernatant was filtered through a 0.22 μm organic filter membrane before injection. Analysis was performed using an Agilent 1260 HPLC system (Agilent Technologies, Santa Clara, CA, USA) equipped with a ZORBAX-C18 column (4.6 mm × 250 mm, 5 μm). An external standard calibration curve was established using glucose standards (R^2^ ≥ 0.99). Serum glucose concentration (mmol/L) was calculated based on peak area, with the mean value was obtained from triplicate measurements.
2.4. Colonic SCFA Analysis (GC-MS)
Frozen colon content samples were thawed, and 50 mg was weighed. One milliliter of pre-treatment solution containing 0.5% H_3_PO_4_ was added. Samples were vortexed for 2 min and centrifuged at 12,000× g for 10 min at 4 °C. The supernatant was filtered through a 0.22 μm filter membrane. Analysis was performed using an Agilent 7890B GC system (Agilent Technologies, Santa Clara, CA, USA) coupled with an MS detector, equipped with a DB-FFAP capillary column (30 m × 0.25 mm × 0.25 μm). External standard calibration curves were established using acetate, propionate, and butyrate standards (R^2^ ≥ 0.99). Matrix effects were corrected using 4-methylvaleric acid as the internal standard. SCFA concentrations were calculated based on characteristic ion peak areas and expressed as μg/g wet weight, with the mean value obtained from triplicate measurements.
2.5. DNA Extraction and Sequencing
Total DNA was extracted from 200 mg of colon content using the FastDNA^®^ Spin Kit (MP Biomedicals, Shanghai, China) according to the manufacturer’s instructions. The ITS1 (internal transcribed spacer) region, a fungal-specific rRNA marker, was amplified using primers ITS1F and ITS2R. PCR products were verified by 1.5% agarose gel electrophoresis, purified using AMPure XP beads (Beckman Coulter, Brea, CA, USA), and stored at −20 °C. Purified amplicons were used to construct TruSeq DNA PCR-Free libraries and sequenced (paired-end, 2 × 250 bp) on an Illumina NovaSeq 6000 platform (Illumina, Inc., San Diego, CA, USA). Raw sequence data were processed within QIIME2 (2023.9): sequences with Q-score < 30 were filtered, retaining >5000 reads/sample. Primers were trimmed using cutadapt, and denoising, chimera removal, and generation of the OTU table were performed using DADA2 (v1.26.0).
2.6. Bioinformatics Analysis
Alpha diversity indices (Shannon index and Chao1 index) were calculated based on the OTU table. Beta diversity differences were visualized using Principal Coordinates Analysis (PCoA) based on Bray–Curtis dissimilarity matrices. OTUs were taxonomically classified, and relative abundances (%) at the genus level were calculated within each sample. Linear Discriminant Analysis Effect Size (LEfSe) was employed to identify fungal taxa differentially abundant between groups. Fungal functional guilds were predicted using the FUNGuild database, and results with a confidence rating of “Probable” or higher were retained for analysis.
2.7. Statistical Analysis
All data were tested for normality (Shapiro–Wilk test) and homogeneity of variance (Levene’s test). Non-normally distributed data were log-transformed prior to analysis. Differences between groups (NX vs. DLY) for serum glucose, SCFA concentrations, and alpha diversity indices (Shannon, Chao1) were assessed using independent samples t-tests (for normally distributed data with homoscedasticity) or Mann–Whitney U tests (for non-normally distributed data or heteroscedasticity). Beta diversity differences based on Bray–Curtis distance matrices were evaluated using Permutational Multivariate Analysis of Variance (PERMANOVA) with 999 permutations. For LEfSe analysis, taxa with a Linear Discriminant Analysis (LDA) score > 2.5 and a Kruskal–Wallis test p-value < 0.05 were considered significant, and the Benjamini–Hochberg procedure was used to control the False Discovery Rate (FDR) for multiple hypothesis testing. Differences in predicted FUNGuild functional guild abundances between groups were assessed using the Kruskal–Wallis test (p < 0.05). Spearman’s rank correlation analysis (|r| > 0.5, p < 0.05) was performed to assess relationships between significantly different fungal taxa (or functional guilds) and serum glucose or intestinal SCFA concentrations.
Statistical analyses were performed using R (4.3.2). Data visualization was conducted using the Python Seaborn (0.12.2) and Matplotlib (3.7.1) libraries. Microbiome community data were processed using QIIME2 (2023.9). Statistical significance was set at p < 0.05, and trends were noted for 0.05 ≤ p < 0.10.
3. Results
3.1. Phenotypic Differences
NX pigs exhibited significantly higher fat percentage (36.8% ± 1.9%) and lower lean meat percentage (51.7% ± 2.1%) compared to DLY pigs (fat percentage: 20.2% ± 2.1%; lean meat percentage: 67.8% ± 3.2%) (p < 0.001) (Figure 1).
Further analysis revealed lower colonic SCFA concentrations in NX pigs, with acetate being the most abundant SCFA (91 ± 15 vs. 128 ± 17 μg/g, p = 0.015). Propionate (41 ± 11 vs. 79 ± 12 μg/g, p < 0.001) and butyrate (24 ± 8 vs. 48 ± 11 μg/g, p < 0.001) showed highly significant reductions. Conversely, serum glucose concentration was markedly elevated in NX pigs (5.8 ± 0.4 vs. 4.2 ± 0.3 mmol/L, p < 0.001) (Figure 2). These results suggest divergent metabolic pathways for SCFAs and glucose between the two breeds.
3.2. Associations Between Body Composition and Metabolites
Scatterplot matrices (Figure 3a) revealed complex relationships in NX and DLY pigs: fat percentage showed a significant positive correlation with serum glucose (r = 0.78, p = 0.0056, Figure 3b) and a significant negative correlation with total colonic SCFAs (r = −0.60, p < 0.001, Figure 3b). The weak association between SCFAs and glucose suggests relatively independent regulatory pathways. This indicates that glucose metabolism may synergistically drive high fat deposition in NX pigs, while SCFAs—as key substrates for intestinal energy metabolism—likely promote fat consumption by inhibiting adipocyte differentiation or stimulating lipolysis.
3.3. Alpha and Beta Diversity Analysis
NX pigs harbored significantly higher gut fungal alpha diversity (Shannon index: 3.5 ± 0.2 vs. 2.8 ± 0.3; Chao1 index: 140 ± 13 vs. 97 ± 14; p < 0.05; Figure 4a). Beta diversity analysis (PCoA based on Bray–Curtis distances) revealed clear separation between breeds (PERMANOVA R^2^ = 0.18, p = 0.002), indicating distinct community structures (Figure 4b). While bacterial communities (e.g., Firmicutes/Bacteroidetes ratio) were not sequenced here, prior studies confirm their dominant role in SCFA production [15,16]. Our fungal-focused approach reveals previously overlooked regulatory pathways.
3.4. Fungal Compositional Differences
Taxonomic analysis showed significant enrichment of Aspergillus (relative abundance: 22.2% ± 1.8% vs. 20.2% ± 1.6%) and Penicillium (12.5% ± 1.2% vs. 10.6% ± 1.4%) in NX pigs (p < 0.01), while Rhodotorula dominated in DLY pigs (14.3% ± 1.5% vs. 9.6% ± 1.4%, p < 0.01) (Figure 5a). LEfSe analysis identified Aspergillus (LDA = 2.70, p = 0.01) and Penicillium (LDA = 2.65, p = 0.006) as discriminative taxa for NX pigs, and Rhodotorula for DLY pigs (LDA = 2.74, p < 0.001) (Figure 5b). These compositional differences suggest evolutionary divergence in energy metabolism strategies.
3.5. Fungal Functional Prediction and Metabolic Correlations
FUNGuild annotation revealed similar nutritional modes but significant differences in specific functional guilds. DLY pigs had significantly higher abundance of the “Animal Pathogen” guild (p < 0.05) (Figure 6), suggesting greater pathogenic colonization risk. Conversely, NX pigs may possess enhanced resistance to pathogens.
Correlation analysis showed that the dominant NX genera (Aspergillus, Penicillium) strongly correlated negatively with SCFAs (Penicillium-propionate: r = −0.72, p < 0.01) and positively with serum glucose (Aspergillus: r = 0.51, p < 0.01). In DLY pigs, Rhodotorula correlated positively with SCFAs (propionate: r = 0.85, p < 0.001) and negatively with glucose (r = −0.78, p < 0.001) (Figure 7), indicating co-evolution between fungal function and host metabolic phenotype.
4. Discussion
This study reveals a novel mechanism by which the gut fungal community regulates fat deposition in NX pigs through microbial metabolites. Notably, NX pigs exhibited a significantly higher fat deposition rate compared to DLY pigs. Their serum glucose levels were elevated by 38.1%, while the concentration of total SCFAs in colonic contents was 38.8% lower than in DLY pigs. Specifically, acetate, propionate, and butyrate decreased by 29.0%, 48.1%, and 50.0%, respectively. Further analysis revealed a significant negative correlation between the fat rate and total colonic SCFAs concentration, and a significant positive correlation with serum glucose concentration. This association suggests that the high fat deposition in NX pigs may be closely linked to the accumulation of glucose in their serum. Furthermore, butyrate, a key substrate in energy metabolism, might drive fat consumption by inhibiting adipocyte differentiation or promoting lipolysis.
Analysis of ITS sequencing revealed a significant specificity in the gut fungal community of NX pigs: the relative abundance of Aspergillus and Penicillium increased compared to DLY pigs, while the abundance of Rhodotorula decreased. Studies indicate that Penicillium spp. can secrete glucose oxidase, leading to increased glucose consumption in the local microenvironment and consequently elevated circulating blood glucose levels [17,18]. The cellulase systems of Aspergillus spp. may enhance polysaccharide breakdown within the host gut, increasing glucose bioavailability [19,20]. Our study also found a strong positive correlation between the abundance of the Aspergillus/Penicillium complex and serum glucose levels, suggesting that these gut fungi promote fat deposition, at least partly, through a monosaccharide supply regulatory axis. Mechanistically, serum glucose likely serves as a substrate for de novo lipogenesis in the liver. It may upregulate the expression of key enzymes such as fatty acid synthase and acetyl-CoA carboxylase via the insulin signaling pathway [21,22], thereby explaining the characteristic subcutaneous fat deposition in NX pigs. This proposed glucose-driven mechanism aligns with findings in Jinhua pigs, where an Aspergillus-enriched microbiota correlated with elevated lipogenesis [23].
Complementing this glucose-mediated pathway, our data also implicate reduced SCFA biosynthesis, linked to lower Rhodotorula abundance, in promoting adiposity. We observed a significant positive correlation between Rhodotorula abundance and colonic SCFAs concentration. Although existing research does not directly establish a link between Rhodotorula abundance and gut SCFA levels, it may influence SCFA production through interactions with other gut microbes. For instance, an in vitro study evaluated the nutritional quality of microbial samples from bacteria, filamentous fungi, and yeasts, including a yeast strain (Rhodotorula) [24]. This study measured in vitro protein digestibility and carbohydrate colonic fermentation, including SCFA production. Thus, the reduction in Rhodotorula observed in NX pigs may impair gut SCFA biosynthesis, particularly butyrate. As a crucial regulator of lipid metabolism, butyrate can inhibit adipocyte differentiation by activating AMP-activated protein kinase phosphorylation [25,26,27]. The consequent reduction in butyrate concentration in NX pigs may therefore alleviate the inhibition of adipocyte differentiation, creating a microenvironment conducive to lipid deposition. Collectively, this ‘Glucose–SCFAs axis’ mediated by gut fungi provides a dual mechanism promoting fat deposition in NX pigs, complementing known bacterial contributions such as acetate production by Bacteroides spp. for fat storage and butyrate-induced leptin expression by Roseburia [28,29].
Notably, this study challenges the traditional view in prior research that emphasizes the dominant role of bacterial SCFAs in lipid metabolism [30], uncovering the potential contribution of gut fungi in monogastric animals. Unlike ruminants where fungi degrade fiber in foreguts [31,32], porcine fungi directly modulate host glucose metabolism—a distinction arising from monogastric physiology and high-starch diets. This species-specific mechanism necessitates breed-customized microbial modulation strategies for the livestock industry. Practically, manipulating gut fungi offers targeted solutions: for NX pigs, reducing Aspergillus/Penicillium (e.g., via antifungal feed additives) may decrease excessive fat deposition; for DLY pigs, supplementing Rhodotorula could enhance intramuscular fat for superior meat quality. This approach is particularly valuable for indigenous breeds—optimizing NX pigs’ pathogen resistance with fat modulation could maximize their use in specialty pork production. Industrial applications could include balancing lipogenesis in fatty breeds (Rhodotorula enhancement) or improving flavor in lean-type pigs (targeted Aspergillus/Penicillium use). However, while bioinformatic correlations link fungal metabolism to fat deposition, direct causality requires validation through fungal isolation, culture, and in vitro studies.
5. Conclusions
The high fat rate in NX pigs is associated with glucose enrichment and the dominance of cellulose-degrading fungi (Aspergillus/Penicillium), whereas DLY pigs’ leanness associates with Rhodotorula-linked SCFA production. Targeted modulation of the gut fungal community could potentially optimize pig production performance and provide a theoretical basis for the utilization of indigenous breed resources. Targeting key fungi (e.g., probiotics/antifungals) could fine-tune fat/lean ratios in swine production, improving economic efficiency and product diversity.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bordbar F. Mohammadabadi M. Jensen J. Xu L. Li J. Zhang L. Identification of Candidate Genes Regulating Carcass Depth and Hind Leg Circumference in Simmental Beef Cattle Using Illumina Bovine Beadchip and Next-Generation Sequencing Analyses Animals 202212110310.3390/ani 1209110335565529 PMC 9102740 · doi ↗ · pubmed ↗
- 2Mohammadabadi M. Bordbar F. Jensen J. Du M. Guo W. Key Genes Regulating Skeletal Muscle Development and Growth in Farm Animals Animals 20211183510.3390/ani 1103083533809500 PMC 7999090 · doi ↗ · pubmed ↗
- 3He J. Zhang P. Shen L. Niu L. Tan Y. Chen L. Zhao Y. Bai L. Hao X. Li X. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism Int. J. Mol. Sci.202021635610.3390/ijms 2117635632887215 PMC 7503625 · doi ↗ · pubmed ↗
- 4Portincasa P. Bonfrate L. Vacca M. De Angelis M. Farella I. Lanza E. Khalil M. Wang D.Q.H. Sperandio M. Di Ciaula A. Gut Microbiota and Short Chain Fatty Acids: Implications in Glucose Homeostasis Int. J. Mol. Sci.202223110510.3390/ijms 2303110535163038 PMC 8835596 · doi ↗ · pubmed ↗
- 5Lu Y. Fan C. Li P. Lu Y. Chang X. Qi K. Short Chain Fatty Acids Prevent High-fat-diet-induced Obesity in Mice by Regulating G Protein-coupled Receptors and Gut Microbiota Sci. Rep.201663758910.1038/srep 3758927892486 PMC 5124860 · doi ↗ · pubmed ↗
- 6Den Besten G. Bleeker A. Gerding A. van Eunen K. Havinga R. van Dijk T.H. Oosterveer M.H. Jonker J.W. Groen A.K. Reijngoud D.J. Short-Chain Fatty Acids Protect Against High-Fat Diet–Induced Obesity via a PPARγ-Dependent Switch From Lipogenesis to Fat Oxidation Diabetes 2015642398240810.2337/db 14-121325695945 · doi ↗ · pubmed ↗
- 7Giovanni T. Vincenzo C. What are the common downstream molecular events between alcoholic and nonalcoholic fatty liver?Lipids Health Dis.2024234110.1186/s 12944-024-02031-138331795 PMC 10851522 · doi ↗ · pubmed ↗
- 8Milton-Laskibar I. Marcos-Zambrano L.J. Gómez-Zorita S. de Santa Pau E.C. Fernández-Quintela A. Martínez J.A. Portillo M.P. Involvement of microbiota and short-chain fatty acids on non-alcoholic steatohepatitis when induced by feeding a hypercaloric diet rich in saturated fat and fructose Gut Microbiome 20223 e 510.1017/gmb.2022.239295781 PMC 11406367 · doi ↗ · pubmed ↗