Role of AdamTS-B in Drosophila wing vein formation
Jana Schulze, Uwe Töpfer

TL;DR
This study shows that AdamTS-B is important for the correct formation of wing veins in fruit flies.
Contribution
The paper identifies AdamTS-B as a novel factor involved in Drosophila wing vein development.
Findings
Knockdown of AdamTS-B causes abnormal vein patterns like extra veins and wandering veins.
The effect is specific to longitudinal veins, with no changes in cross veins.
AdamTS-B appears to influence morphogen spreading during wing development.
Abstract
The development of Drosophila wing veins is a complex morphogenetic process that depends on the interplay of different signaling pathways, including EGFR, BMP, Notch, Hedgehog and Wnt. Basement membranes (BMs) and proteases that process BM components play a crucial role in controlling the morphogen spreading and associated patterning required for proper organ formation. Here we show, that AdamTS-B is required for the proper development of Drosophila wing veins. Knockdown of AdamTS-B results in various phenotypes, including additional veins, delta branches, and wandering veins within different longitudinal veins, though there are no differences in the cross veins.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
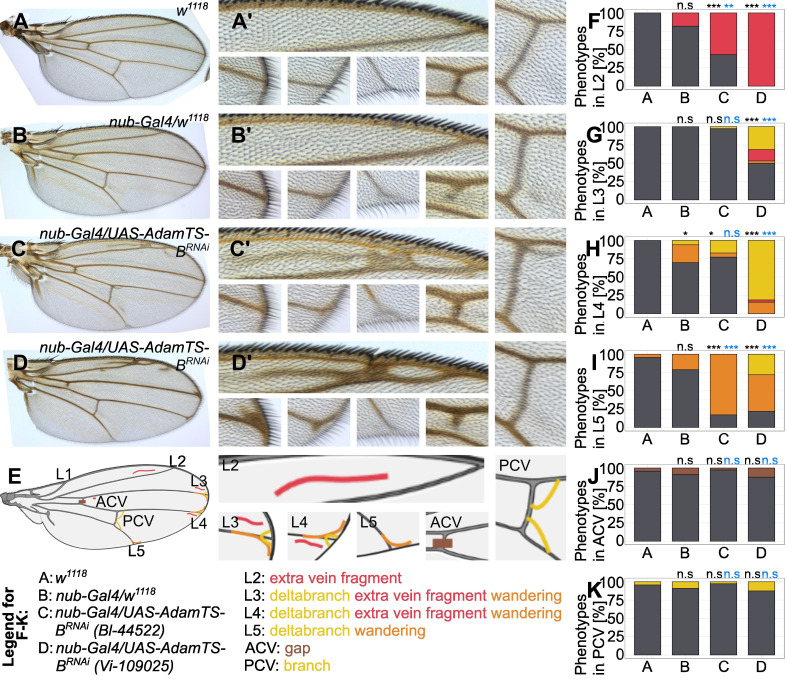 Figure 1
Figure 1|
|
|
|
| |
|
nub-Gal4 |
|
BDSC_25754 |
Bloomington Drosophila Stock Center | |
|
w 1118 |
|
BDSC_3605 |
Bloomington Drosophila Stock Center | |
|
UAS-AdamTS dsRNA |
|
BDSC_44522 |
Bloomington Drosophila Stock Center | |
|
UAS-AdamTS dsRNA |
|
VDRC_109025 |
Vienna Drosophila Resource Center | |
- —Deutsche Forschungsgemeinschaft (Germany)https://ror.org/018mejw64
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDevelopmental Biology and Gene Regulation · Hippo pathway signaling and YAP/TAZ · Neurobiology and Insect Physiology Research
Description
The BM is a thin extracellular matrix that underlies epithelia and surrounds most tissues and organs, thereby mediating biophysical and biochemical cues for cells required for morphogenesis (Khalilgharibi & Mao, 2021; Sherwood, 2021; Töpfer, 2023). Those biochemical cues are mainly regulated by the ability of the BM to bind morphogens that are required for patterning (Patel et al., 2008; Wang et al., 2008). The remodeling of the BM by protease cleavage and subsequent degradation of the BM components is consequently an important step in the limitation of morphogens, patterning and morphogenesis. The Drosophila wing disc is composed of two opposing epithelial layers, which share a common BM that surrounds the organ. The formation of the Drosophila wing veins depends on the strict control of BMP morphogen distribution as well as the antagonism of Notch and EGF signaling, Hedgehog and Wnt pathways (Blair, 2007). The precise regulation of these patterning leads to a fully developed wing with five longitudinal veins (L1-L5) and two cross veins (anterior (ACV) and posterior (PCV)) ( Fig. 1A ). The AdamTS gene group is known to cleave extracellular matrix components and be important for cell migration and organ sculpting (Agarwal et al., 2022; Gonsior & Ismat, 2019; Ismat et al., 2013; Lhamo & Ismat, 2015; Skeath et al., 2017; Töpfer et al., 2024). * AdamTS-B * is one of four genes in Drosophila encoding a ‘A disintegrin and metalloprotease with thrombospondin motif' and an ortholog of human AdamTS-7 and AdamTS12 (Öztürk-Çolak et al., 2024). Knockdown of * AdamTS-B * has been shown to result in wing vein formation phenotypes. Butchar et al. 2012 identified * AdamTS-B * as a downstream gene of EGFR signaling and a knockdown with a ubiquitous Gal4 driver ( Act5C-Gal4 ) led to an extra vein in L2. Based on this, Pham et al 2018 dissected the wing phenotypes of * AdamTS-B * using different driver lines and different UAS-dsRNA lines. In this study, we replicated an experiment form Butchar et al. 2012. and Pham et al. 2018 by knocking down * AdamTS-B , * but with other genetic and environmental conditions.
For tissue-specific knockdown, we used * nubbin-Gal4 ( nub-Gal4 ), * which is active in the developing pouch region, the cells that give rise to the adult wing (Azpiazu & Morata, 2000). We analyzed the morphology of wings from * w ^ 1118 ^ * (Fig.1A and A') and * nub-Gal4 /+ * (Fig.1B and B') control flies as well as two different UAS-dsRNA lines for * AdamTS-B * knockdown * , nub-Gal4 /UAS- AdamTS-B ^RNAi^ ( BDSC_44522 ) * (Fig.1C and C') and * nub-Gal4 /UAS- AdamTS-B ^RNAi^ ( VDRC_109025 ) * (Fig.1D and D'). First, we quantified the presence of extra veins in L2 (Fig.1A-F). Knockdown of * AdamTS-B * with the BDSC_44522 (from here on BDSC line) and the VDRC_109025 (from here on VDRC line), which showed full penetrance of the phenotype, resulted in significant differences to * w ^1118^ * as well as to * nub-Gal4 /+ * controls (Fig.1F). In contrast to L2, in the L3-L5 regions, in addition to the extra vein phenotypes, we also observed the presence of delta branches and so-called wandering veins (illustrated in Fig.1E, Fig.1A-D' and G-I). In the L3 region, we observed the presence of all three phenotypes (extra vein, delta branch and wandering vein) with significant differences between the * AdamTS-B * knockdown with the VDRC line compared to both controls, while we found no differences for the BDSC line (Fig.1G). In L4, we found significant differences between the two controls with wandering veins and delta branches in * nub-Gal4 /+ * controls (Fig.1H), as well as a significant difference between * w ^ 1118 ^ * and the BDSC knockdown line. * AdamTS-B * knockdown with the VDRC line resulted in fully penetrance with all three possible phenotypes and significant differences compared to both controls (Fig.1H). In the last region for longitudinal veins, L5, both knockdown lines show significant differences compared to both controls, but only delta branches and wandering veins and no extra veins (Fig.1I). Finally, in the region of ACV and PCV, we found no differences in terms of the phenotypes just described and also presence of gaps, which are present in low penetrance in all genotypes in the AVC (Fig.1J).
In summary we can draw four conclusions. First, our results are consistent with the finding of Butchar et al. 2012 and Pham et al.2018, that knockdown of * AdamTS-B * can lead to extra veins in L2 (Fig.1F). Notably, while Pham et al. 2018 described extra veins in L2 with the MS1069-Gal4 driver, they found a decreased percentage of extra veins in L2 with * AdamTS-B * knockdown using * nub-Gal4 * , which is in contrast to our results. Second, in L3 and L4 the VDRC line shows various phenotypes, namely extra veins, delta branches and wandering veins, that are not significantly different between the * nub-Gal4 * control and the BDSC line (Fig.1G and H). These observations have not been described previously. Third, in L5 we observe delta branches and wandering veins in both * AdamTS-B * knockdown conditions (Fig.1I). Wandering veins in L5 were also described in Pham et al. 2018. Fourth, the cross veins are not affected by * AdamTS-B * knockdown. To conclude, while our results mainly support the findings of Butchar et al. 2012 and Pham et al. 2018, on the one hand we show a result that is in contrast to Pham et al. 2018, specifically the knockdown of * AdamTS-B * led in our results to an increase of extra veins in L2 and in Pham et al. 2018 to a decrease. On the other hand, we also found a much higher penetrance of phenotypes in every longitudinal vein, at least with the VDRC line. One reason might be the difference regarding culturing conditions. In contrast to Pham et al. 2018, we kept our crosses at 29 °C, which might increase RNAi efficiency. In line with Pham et al. 2018 we observed differences regarding the different UAS lines. The VDRC line shows an overall much higher penetrance of phenotypes than the BDSC line. However, it is unclear whether the higher penetrance of the phenotypes in the VDRC line is due to a possible higher knockdown efficiency of this line or whether this line shows off-target effects in contrast to the BDSC.
AdamTS-B * was identified as a target gene of EGFR pathway using microarray experiments, where ectopic activation of EGFR results in increased AdamTS-B expression (Butchar et al., 2012). The fact that reduction of * AdamTS-B * through RNAi led to the presence of extra veins (Butchar et al., 2012; Pham et al., 2018) and ectopic expression of * AdamTS-B * to the loss of veins (Pham et al., 2018) are reminiscent to the modulation of the EGFR signaling pathway (Brentrup et al., 2000; Sturtevant et al., 1993; Sturtevant & Bier, 1995). In addition, a genetic interaction analysis in which * AdamTS-B * knockdown phenotypes were attempted to be rescued revealed that genetic reduction of the EGFR ligands kern in combination with either vein or spitz reduces the proportion of wings with extra veins, suggesting a direct or indirect involvement of * AdamTS-B * in EGFR signaling at the ligand level (Butchar et al., 2012). Our results support the idea that * AdamTS-B * negatively regulates EGFR pathways, hence in our condition * AdamTS-B * knockdown shows stronger phenotypes in all longitudinal veins, which is evident since EGFR affects all longitudinal veins (Fig.1A-I). The exact mechanism by which * AdamTS-B * modulates the EGFR pathway and longitudinal vein formation is unknown. We propose a model in which * AdamTS-B * influences activation of EGFR signaling, controlling the morphogenesis of all five longitudinal veins, by remodeling the BM during larval stages when cell fate is determined by morphogen gradients (De Celis, 1998) and before the BM is degraded during wing growth and elongation (Diaz-de-la-Loza et al., 2018).
Methods
** Drosophil a genetics and husbandry **
The fly stocks used were * nub-Gal4 * [Bloomington Drosophila Stock Center (BDSC), BDSC_25754], * w ^1118^ * (BDSC_3605), * UAS- AdamTS-B ^dsRNA^ * (BDSC_44522) and UAS- * UAS- AdamTS-B ^dsRNA^ * [Vienna Drosophila Resource Center (VDRC), VDRC_109025]. Flies were kept at 25°C on standard food. Crosses were raised at 29°C.
Wing dissection
Flies were stunned with CO 2 and the desired female flies were transferred to a block dish containing isopropanol. The wings were dissected from the body using forceps. The wings were transferred to tubes containing isopropanol using a cut pipette tip and stored until mounting in a mixture of isopropanol and Euparal (ratio 1:1).
Wing quantification
The pictures were taken using the bright field of the Observer.Z1 (air objective: 5x). For wings, that can't be captured with one photograph, two pictures were made and afterwards stitched together using ImageJ (Preibisch et al., 2009). Each wing was analysed for all veins.
Statistical analysis
To evaluate significances Chi-square testin R was used (R Core Team, 2021).
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agarwal Priti Shemesh Tom Zaidel-Bar Ronen 202291 Directed cell invasion and asymmetric adhesion drive tissue elongation and turning in C. elegans gonad morphogenesis Developmental Cell 57171534-580721112126.e 610.1016/j.devcel.2022.08.00336049484 · doi ↗ · pubmed ↗
- 2Azpiazu Natalia Morata Ginés 2000615 Function and regulation of homothorax in the wing imaginai disc of Drosophila Development 127120950-19912685269310.1242/dev.127.12.268510821766 · doi ↗ · pubmed ↗
- 3Blair Seth S. 2007111 Wing Vein Patterning in Drosophila and the Analysis of Intercellular Signaling Annual Review of Cell and Developmental Biology 2311081-070629331910.1146/annurev.cellbio.23.090506.12360617506700 · doi ↗ · pubmed ↗
- 4Brentrup Doris Lerch Hans-Philipp Jäckle Herbert Noll Markus 2000111 Regulation of Drosophila wing vein patterning: net encodes a b HLH protein repressing rhomboid and is repressed by rhomboid-dependent Egfr signalling Development 127210950-19914729474110.1242/dev.127.21.472911023875 · doi ↗ · pubmed ↗
- 5Butchar Jonathan P Cain Donna Manivannan Sathiya N Mc Cue Andrea D Bonanno Liana Halula Sarah Truesdell Sharon Austin Christina L Jacobsen Thomas L Simcox Amanda 201281 New Negative Feedback Regulators of Egfr Signaling in Drosophila Genetics 19141943-26311213122610.1534/genetics.112.14109322595244 PMC 3416002 · doi ↗ · pubmed ↗
- 6De Celis JF 1998 Positioning and differentiation of veins in the Drosophila wing.Int J Dev Biol 4230214-62823353439654017 · pubmed ↗
- 7Diaz-de-la-Loza Maria-del-Carmen Ray Robert P. Ganguly Poulami S. Alt Silvanus Davis John R. Hoppe Andreas Tapon Nic Salbreux Guillaume Thompson Barry J. 201871 Apical and Basal Matrix Remodeling Control Epithelial Morphogenesis Developmental Cell 4611534-58072339.e 510.1016/j.devcel.2018.06.00629974861 PMC 6035286 · doi ↗ · pubmed ↗
- 8Gonsior Madison Ismat Afshan 20191024 sli is required for proper morphology and migration of sensory neurons in the Drosophila PNS Neural Development 1411749-810410.1186/s 13064-019-0135-z PMC 681307831651354 · doi ↗ · pubmed ↗