The Effect of APOE ε4 Allele on Dynamic Local Spontaneous Brain Activity and Functional Integration in Alzheimer's Disease
Yi Tan, Dan Yang, Zhihong Ke, Zheqi Hu, Wenting Song, Limoran Tang, Zhixin Zhou, Yuting Mo, Lili Huang, Yun Xu

TL;DR
This study explores how the APOE ε4 gene affects brain activity and cognitive functions in Alzheimer's disease patients.
Contribution
The study identifies how APOE ε4 influences dynamic brain activity and its link to cognitive decline and plasma biomarkers in Alzheimer's disease.
Findings
APOE ε4 carriers showed unique correlations between brain activity in specific regions and cognitive scores.
Voxel-wise concordance in the right caudate nucleus mediated the relationship between plasma Aβ42 and language function.
A combined model using brain metrics and biomarkers effectively distinguished Alzheimer's patients from controls.
Abstract
The apolipoprotein E (APOE) ε4 allele is the most important genetic risk factor for sporadic Alzheimer's disease (AD), yet its mechanisms in AD pathology and cognitive decline remain unclear. Using a sliding‐time window approach to directly quantify the instantaneous fluctuations of various local metrics based on continuous time series and calculate voxel‐wise concordance of these metrics, we explored the impact of APOE ε4 on dynamic local brain activity and functional integration in AD, and its interrelations with plasma biomarkers and cognition. Results showed that APOE ε4 widely affected dALFF, dReHo, dGSCorr, and voxel‐wise concordance. For AD patients, APOE ε4 carriers uniquely exhibited correlations between dALFF in the right angular gyrus/supramarginal gyrus and MoCA scores and orientation function, and between voxel‐wise concordance in the right caudate nucleus (CAU) and general…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
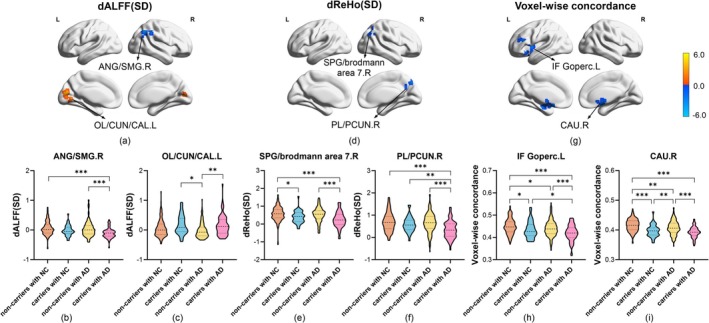 FIGURE 1
FIGURE 1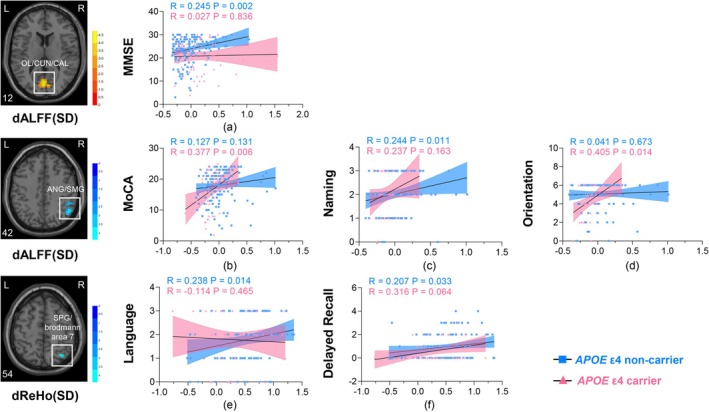 FIGURE 2
FIGURE 2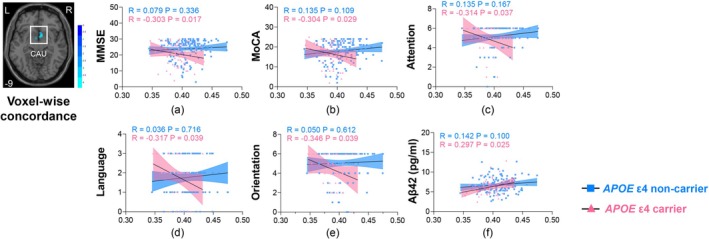 FIGURE 3
FIGURE 3 FIGURE 4
FIGURE 4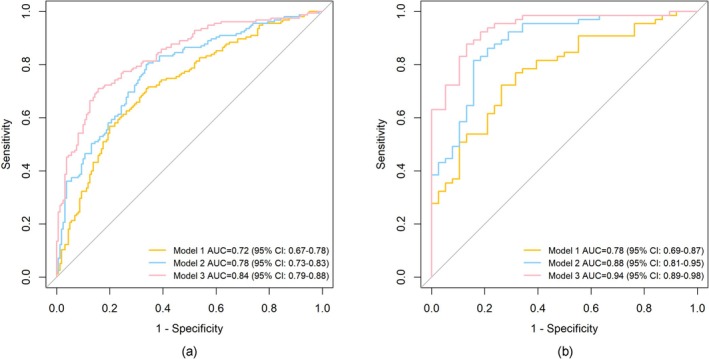 FIGURE 5
FIGURE 5| Ɛ4 non‐carriers with NC | Ɛ4 carriers with NC | Ɛ4 non‐carriers with AD | Ɛ4 carriers with AD |
| |
|---|---|---|---|---|---|
|
|
|
|
| ||
| Demographics | |||||
| Age (years) | 66.68 ± 8.16 | 65.92 ± 10.27 | 69.79 ± 8.70 | 70.66 ± 8.97 | 0.001 |
| Female ( | 80 (50.0) | 25 (65.8) | 91 (58.7) | 35 (53.8) | 0.228 |
| Education level (years) | 12 (9,15) | 12 (8.25,16) | 9 (7,12) | 12 (6,15) | < 0.001 |
| Risk factors | |||||
| Hypertension ( | 78 (48.8) | 17 (44.7) | 69 (44.2) | 35 (53.8) | 0.611 |
| Diabetes mellitus ( | 47 (29.4) | 12 (32.6) | 47 (30.3) | 20 (30.8) | 0.828 |
| Hyperlipidaemia ( | 67 (41.9) | 15 (39.5) | 62 (40.0) | 25 (38.5) | 0.627 |
| General cognition | |||||
| MMSE | 29 (28,30) | 29 (28,30) | 25 (22,28) | 22 (15,26) | < 0.001 |
| MoCA | 26 (25,28) | 26 (24.75,28) | 19 (15,22) | 19 (12,21) | < 0.001 |
| Cognitive domains | |||||
| Executive function | 5 (4,5) | 4 (3,4.75) | 3 (2,3) | 2 (1,4) | < 0.001 |
| Naming ability | 2 (2,3) | 2 (2,3) | 1 (1,2) | 1 (1,2) | < 0.001 |
| Attention | 6 (6,6) | 6 (6,6) | 6 (5,6) | 5.5 (4,6) | < 0.001 |
| Language function | 3 (2,3) | 3 (2.25,3) | 2 (1,3) | 2 (1,3) | < 0.001 |
| Abstraction | 2 (1,2) | 2 (1,2) | 1 (0,1) | 1 (1,2) | < 0.001 |
| Delayed recall | 3 (2,4) | 3 (2,4) | 0 (0,2) | 0 (0,1.25) | < 0.001 |
| Orientation function | 6 (6,6) | 6 (6,6) | 6 (4.75,6) | 5 (3,6) | < 0.001 |
| AD plasma biomarkers | |||||
| Aβ42 (pg/ml) | 6.94 ± 2.06 | 6.81 ± 1.77 | 6.76 ± 1.85 | 6.09 ± 1.86 | 0.041 |
| Aβ40 (pg/ml) | 103.30 ± 22.73 | 102.65 ± 18.95 | 101.31 ± 20.92 | 102.49 ± 24.49 | 0.901 |
| Aβ42/40 ratio | 0.068 ± 0.016 | 0.067 ± 0.016 | 0.067 ± 0.013 | 0.060 ± 0.012 | 0.003 |
| GFAP (pg/ml) | 118.82 ± 54.84 | 122.86 ± 55.19 | 156.21 ± 150.45 | 210.26 ± 151.06 | < 0.001 |
| NfL (pg/ml) | 24.29 ± 32.92 | 20.79 ± 18.23 | 27.91 ± 29.22 | 28.88 ± 17.84 | 0.398 |
| p‐tau181 (pg/ml) | 2.04 ± 1.04 | 2.18 ± 0.84 | 3.18 ± 1.97 | 4.23 ± 2.52 | < 0.001 |
| Imaging data | |||||
| TIV (ml) | 1354.72 ± 116.93 | 1333.91 ± 128.49 | 1333.48 ± 125.55 | 1324.53 ± 111.51 | 0.262 |
| Dynamic measures | Group/condition effect | Brain regions | Cluster size (voxels) | Peak MNI coordinate | Peak t value | ||
|---|---|---|---|---|---|---|---|
|
|
|
| |||||
| dALFF (CV) | Group effect | R SPG/PCUN/brodmann area 7 | 84 | 15 | −48 | 60 | 4.3350 |
| dALFF (Mean) | Condition effect | R PCUN/brodmann area 7 | 71 | 3 | −63 | 51 | −4.5484 |
| dALFF (SD) | Condition effect | R PCUN/brodmann area 7 | 117 | 3 | −63 | 51 | −5.2175 |
| Group effect | L OL/CUN/CAL | 187 | 0 | −75 | 12 | 4.7950 | |
| Group effect | R ANG/SMG | 123 | 51 | −57 | 42 | −4.6830 | |
| dfALFF (Mean) | Condition effect | L PCUN/brodmann area 7 | 55 | −6 | −66 | 54 | −4.6782 |
| dfALFF (SD) | Condition effect | L SPG/PCUN | 83 | −21 | −60 | 45 | −3.6881 |
| dReHo (SD) | Group effect | R PL/PCUN | 42 | 12 | −72 | 42 | −4.3366 |
| Group effect | R SPG/brodmann area 7 | 65 | 33 | −63 | 54 | −4.3213 | |
| dGSCorr (Mean) | Group effect | L IF Gtriang | 154 | −48 | 27 | 15 | −4.3648 |
| Voxel‐wise concordance | Group effect | R CAU | 184 | 9 | 9 | −9 | −4.3807 |
| Group effect | L IF Goperc | 132 | −48 | 12 | 6 | −4.4705 | |
| Model | Cross‐validated diagnostic performance | |||
|---|---|---|---|---|
| Sensitivity | Specificity | Accuracy | ||
|
| Model 1 | 71.61 | 65.63 | 68.57 |
| Model 2 | 80.65 | 65.63 | 73.02 | |
| Model 3 | 70.97 | 84.38 | 77.78 | |
|
| Model 1 | 72.31 | 73.68 | 72.82 |
| Model 2 | 81.54 | 84.21 | 82.52 | |
| Model 3 | 87.69 | 86.84 | 87.38 | |
- —National Natural Science Foundation of China10.13039/501100001809
- —Jiangsu Province Key Medical Discipline
- —STI 2030 ‐ Major Projects
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFunctional Brain Connectivity Studies · Dementia and Cognitive Impairment Research · Alzheimer's disease research and treatments
Summary
- The APOE ε4 allele widely impacted dynamic local spontaneous brain metrics including dALFF, dReHo, and dGSCorr, and voxel‐wise concordance of these metrics in AD.
- The APOE ε4 allele influenced the correlations of dynamic functional activities with cognition and plasma Aβ pathology in AD.
- The voxel‐wise concordance in the right caudate nucleus partially mediated the relationship between plasma Aβ42 and language function in APOE ε4 carriers with AD.
- The combined model incorporating dynamic metrics, plasma AD biomarkers, and demographic data effectively distinguished AD from NC.
Introduction
1
Alzheimer's disease (AD), a progressive neurodegenerative disease with insidious onset, is the main cause of dementia (Scheltens et al. 2021). AD involves dysregulation of complex cellular and molecular processes (Knopman et al. 2021) and is associated with multiple genetic variations (Schwartzentruber et al. 2021). Among these, the apolipoprotein E (APOE) ε4 allele is the most important genetic risk factor for sporadic ad (Serrano‐Pozo et al. 2021; Uebergang et al. 2023) and plays an important role in AD pathological processes (Haney et al. 2024; Montagne et al. 2020; Blanchard et al. 2022). By age 85, the lifetime risk for AD in APOE ε4 homozygotes reaches 60%, and heterozygotes exhibit semi‐dominant effects of this moderately penetrant gene (Fortea 2024; Genin et al. 2011). However, the mechanism by which the APOE ε4 allele mediates AD pathology remains incompletely understood. Therefore, exploring the underlying pathogenesis of the APOE ε4 allele may provide new therapeutic opportunities for AD patients.
Over the past decades, resting‐state functional magnetic resonance imaging (rs‐fMRI) has been widely used to explore functional abnormalities in the brains of APOE ε4 carriers. The most frequently applied functional metrics include functional connectivity (FC) and local spontaneous brain activity metrics. For example, in healthy human adults, APOE ε4 carriers exhibited increased lateralized connections associated with callosal connections within the default mode, memory, and salience networks compared to non‐carriers (Butt et al. 2022). Decreased FC in the left hippocampus of APOE ε4 carriers was associated with verbal memory decline (Baxter et al. 2023). Regarding local brain activity, APOE ε4 carriers with AD exhibit decreased amplitude of low frequency fluctuations (ALFF) in the left hippocampus (Wang et al. 2015) and decreased fractional ALFF (fALFF) in the left insula, left inferior frontal gyrus, and right precentral gyrus (Lin et al. 2017) compared to non‐carriers. Additionally, cognitively normal young adult APOE ε4 carriers showed increased regional homogeneity (ReHo) in the right superior frontal gyrus which correlated with neuropsychological test scores (Zheng et al. 2017); Cai 2017). also reported altered ALFF and ReHo in the anterior cingulate cortex, medial prefrontal cortex, and precuneus in APOE ε4 carriers with normal cognition (NC), indicating gray matter activity changes in these regions (Cai 2017). Notably, the plasma AD biomarkers such as amyloid beta (Aβ), glial fibrillary acidic protein (GFAP) (Lohman et al. 2024), neurofilament light chain (NfL) (Lohman et al. 2024), and phosphorylated tau (p‐tau) (Zhu et al. 2024) have been associated with APOE ε4‐related functional changes.
However, given that the human brain remains active during the resting state, static rs‐fMRI metrics described previously could not reflect temporal characteristics of local brain activity in APOE ε4 carriers. Fortunately, compared to other dynamic fMRI approaches, such as point process analysis and time frequency analysis, the sliding‐time window method directly quantifies the instantaneous fluctuations of various local rs‐fMRI metrics from continuous time series (Yang et al. 2024; Chen et al. 2023). Since dynamic metrics reflect brain activity from different perspectives, the voxel‐wise concordance derived from them may reveal the interaction among these metrics across states. The sliding‐window approach enables calculation of voxel‐level synchronization of multiple dynamic local metrics, objectively and comprehensively reflecting the mechanism of brain function integration (Yan 2017). A previous study has shown lower global voxel‐wise concordance in patients with AD than in healthy controls (Chen et al. 2023). However, no studies have investigated the effect of the APOE ε4 allele on AD from the perspective of local dynamic metrics and their integration. Therefore, investigating the impact of the APOE ε4 allele on these metrics in AD populations may provide novel insights into AD pathophysiological mechanisms unobtainable from static or single‐metric analyses.
In this study, we compared dynamic ALFF (dALFF), dynamic fALFF (dfALFF), dynamic ReHo (dReHo), and dynamic global signal correlation (dGSCorr), and voxel‐wise concordance of these dynamic metrics across four groups stratified by APOE ε4 allele and cognitive status. We subsequently investigated (1) the effect of APOE ε4 allele on correlations between dynamic metrics and cognition, plasma AD biomarkers in AD; (2) the mediation effect of APOE ε4‐related alterations in dynamic metrics on the relationship between Aβ pathology and cognitive function; (3) the diagnostic performance of combined models in identifying AD patients.
Materials and Methods
2
Participants
2.1
The current study was conducted under the latest version of the Declaration of Helsinki and authorized by the Review board of Nanjing Drum Tower Hospital (Ethical Approval Code: 2022–472‐01). Informed consent was obtained from all participants. All participants were recruited from two medical centers and local communities. Among them, 7 participants were excluded due to excessive head motion (cumulative translation or rotation of > 3.0 mm or 3.0°). 418 individuals were ultimately included in this study. All participants completed APOE genotyping tests and 3.0‐T whole brain MRI scanning, 417 participants completed the Mini‐Mental State Examination (MMSE), 396 participants completed the Montreal Cognitive Assessment (MoCA), 377 participants completed measurements of plasma AD biomarkers including Aβ42, Aβ40, GFAP, NfL, and p‐tau181. Age, sex, and years of education were collected as demographics. Hypertension, diabetes mellitus, and hyperlipidaemia were considered as risk factors. Each participant's blood collection, cognitive tests, and image acquisition were conducted within 1 month.
Dementia and mild cognitive impairment (MCI) were identified by education‐adjusted norms of MMSE and MoCA (Katzman et al. 1988; Gorelick et al. 2011; Lu et al. 2011), respectively. The cut‐off scores of MMSE and MoCA for different education levels were clearly described in previous study (Yang et al. 2023). Dementia and MCI due to probable AD were diagnosed in accordance with the recommendations of the National Institute on Aging‐Alzheimer's Association (Albert et al. 2011; McKhann et al. 2011). Subjects with dementia and MCI due to probable AD were classified into the AD group. APOE ε4 carriers were defined as individuals with at least one ε4 allele. All participants were divided into 160 APOE ε4 non‐carriers with NC, 38 APOE ε4 carriers with NC, 155 APOE ε4 non‐carriers with AD, and 65 APOE ε4 carriers with AD.
The exclusion criteria for both NC and AD groups were as follows: (1) ≤ 50 years old; (2) central nervous system diseases that could cause cognitive impairment, such as vascular cognitive impairment, Parkinson's disease dementia, and dementia with Lewy body; (3) a history of stroke and subarachnoid hemorrhage; (4) other severe neurological diseases, such as brain tumor, epilepsy, and multiple sclerosis; (5) psychiatric diseases, such as major depressive disorder, bipolar affective disorder, autistic, and schizophrenia; (6) severe systemic diseases, such as cancer and heart failure; (7) alcoholism.
Cognitive Assessment
2.2
General cognition was evaluated by both MMSE and MoCA scores. Scores of cognitive domains of MoCA were calculated. Among them, executive function was evaluated using the sum score of Alternating Trail Making, Visuoconstructional Skills (cube), and Visuoconstructional Skills (clock). Attention was evaluated using the sum score of Forward Digit Span, Backward Digit Span, Vigilance, and Serial 7 s. Language function was evaluated using the sum score of Sentence repetition and Verbal fluency. All cognitive examinations were completed by two professional neuropsychologists.
MRI Data Acquisition
2.3
All MRI scans were performed on a Philips Medical Systems 3.0 T scanner. The rs‐fMRI data were collected with the following parameters: high‐resolution 3D T1 imaging: echo time (TE) = 4.6 ms, repetition time (TR) = 9.8 ms, flip angle (FA) = 8°, field of view (FOV) = 250 × 250 mm^2^, number of slices = 192, matrix size = 192 × 256 × 256, thickness = 1.0 mm; rs‐fMRI was acquired by a gradient‐echo‐planar imaging sequence: TE = 30 ms, TR = 2000 ms, FA = 90°, FOV = 192 × 192 mm^2^, matrix size = 64 × 64, thickness = 4.0 mm, number of slices = 35, and each functional image contained 240 volumes. Besides, T2‐weighted and diffusion‐weighted imaging were obtained to exclude acute or sub‐acute infarctions. All participants were asked to keep their eyes closed, not to move, not to sleep, and not to think about anything during scanning.
Volume Quantification
2.4
The total intracranial volume (TIV) was calculated with the Statistical Parametric Mapping 8 (SPM8, http://www.fil.ion.ucl.ac.uk/spm) based on 3D‐T1 images.
Rs‐fMRI Data Preprocessing
2.5
Original functional images were preprocessed with a standard pipeline in the toolbox for data processing and analysis of brain imaging (DPARSF, V3.2, http://www.restfmri.net) and SPM 12 (http://www.fil.ion.ucl.ac.uk/spm). The specific process was as follows: (1) The first 10 volumes of data were removed; (2) The slice timing correction and realignment were conducted on the remaining volumes; (3) The re‐alignment was performed to correct the movement between time points; (4) Head motion parameters were calculated by evaluating the translation in each direction and the angular rotation on each axis for each volume, then individuals with cumulative translation > 3.0 mm or rotation > 3.0° were excluded; (5) Single T1‐weighted images were segmented into gray matter, white matter, and cerebrospinal fluid and co‐registered to the mean functional image through a 6° freedom linear transformation without re‐sampling; (6) Linear trends, Friston 24 head motion parameters, the white matter signal and cerebrospinal fluid signal were regressed out from the functional signal as nuisance covariates to reduce head motion artifacts. The global signal regression was not performed due to the attendant controversy (Murphy and Fox 2017); (7) The Diffeomorphic Anatomical Registration Through Exponentiated Lie algebra (DARTEL) tool was used to generate transformations from individual native space to the standard Montreal Neurological Institute (MNI) space (3 × 3 × 3 mm^3^); (8) For ReHo and GSCorr, bandpass filters (0.01 Hz < f < 0.1 Hz) were applied.
Calculation of Dynamic Local Metrics
2.6
(1) ALFF (Yu‐Feng et al. 2007)/fALFF (Zou et al. 2008): ALFF and fALFF were used to measure amplitude by transforming the blood oxygen level dependent time course to the frequency domain via fast Fourier transform. ALFF was computed as the mean of amplitudes within a specific low‐frequency range (0.01–0.1 Hz). fALFF was the ratio of the sum of amplitudes of a given low frequency band (0.01–0.1 Hz) to that across the entire frequency range. In this study, fALFF was chosen to calculate voxel‐wise concordance because of the high co‐linearity between these two metrics and the sensitivity and specificity of fALFF in spontaneous brain activity (Yan et al. 2013; Zuo et al. 2010).
(2) ReHo (Zang et al. 2004): ReHo was used to assess the degree of regional coherence. It was the Kendall's coefficient of concordance or Kendall's W of the time series of a given voxel with those of its nearest neighbors (27 voxels).
(3) GSCorr (Hahamy et al. 2014): GSCorr was calculated as the Pearson's correlation coefficient between the global average signal and each voxel signal within the gray matter mask. These correlation values were then Fisher‐Z transformed.
Dynamic local metrics were calculated via sliding time window method in the Data Processing and Analysis of Brain Imaging (DPABI, V6.1, http://rfmri.org/DPABI). Specifically, the sliding window size of 30 TR and sliding step of 1 TR was first applied to perform window time series analyses, resulting in 181 windows for each subject. Within each window, dynamic local metrics were calculated in a voxel‐wise way. The standard deviation (SD), coefficient of variation (CV), and mean maps of these dynamic metrics were then generated. The SD maps were used to assess stability of each metric, CV maps were used to assess variability of each metric, and mean maps were used to quantify the temporal dynamic characteristics. There was no CV map of dGSCorr because it contained negative values. Finally, these SD, CV, and mean maps were subjected to z‐standardization and smoothed with a 4 × 4 × 4 mm^3^ full‐width at half maximum (FWHM) Gaussian Kernel.
Calculation of Voxel‐Wise Concordance
2.7
Voxel‐wise concordance was obtained from the Kendall's W of dfALFF, dReHo, and dGSCorr across time windows for each voxel. Maps of voxel‐wise concordance were transformed with z‐standardization to reduce the effect of variability of global activities among subjects. These maps were then spatially smoothed with a Gaussian kernel of 4 × 4 × 4 mm^3^ FWHM.
To verify the robustness of this study, results using different window lengths (window length = 40 TR, window step = 1 TR) as well as different window steps (window length = 30 TR, window step = 2 TR) were analyzed.
Statistical Analysis
2.8
Normality of continuous variables was assessed by the Shapiro–Wilk test. The Levene's test was used to examine the homogeneity of variance. One‐way analyses of variance (ANOVA) tests and Kruskal‐Wallis H‐tests were used to compare continuous numerical variables across the four groups, as appropriate. A Chi‐squared test was used to compare the distribution of sex among the four groups. Bonferroni correction and Kruskal‐Wallis ANOVA were used as post hoc tests for characteristics at baseline. p < 0.05 was considered statistical significance. All the above statistical analyses were conducted in the SPSS software (version 22.0; IBM Corporation, Armonk, NY, USA). Based on the results of the baseline analysis, age, sex, and years of education were included as covariates in all the following analyses to reduce the impact of confounding variables.
Mixed effect analyses of dynamic rs‐fMRI metrics were performed by DPABI (V6.1, http://rfmri.org/DPABI) within a gray matter mask (GreyMask_02_61 × 73 × 61), with age, sex, and years of education as covariates. APOE ε4 status was considered a group effect variable, while cognitive status was considered a condition effect variable. Multiple comparisons correction was performed using Gaussian Random Field (GRF) theory. The voxel‐level threshold was p < 0.001 for dALFF, dfALFF, dReHo, and dGSCorr, and was p < 0.01 for voxel‐wise concordance. The cluster‐level threshold was p < 0.05. For each metric, a cluster size threshold of 40 voxels was applied. Finally, the values of each cluster were extracted. ANOVA tests followed by Bonferroni correction were then used to compare values of these metrics across the four groups.
To explore the effect of APOE ε4 allele on relationship between dynamic rs‐fMRI metrics and cognition, plasma AD pathology, partial correlation analyses were performed in both APOE ε4 carriers and non‐carriers in AD group, adjusting for age, sex, and years of education.
To further explain the interrelations amongst Aβ42, voxel‐wise concordance, and cognition, mediation analyses were conducted among APOE ε4 carriers with AD, adjusting for age, sex, and years of education. The bootstrapping (k = 5000 samples) in PROCESS for SPSS 22.0 was used to obtain the bias‐corrected 95% confidence interval (CI) for the mediating effect. The mediating effect was considered statistically significant if the 95% CI did not contain the value 0.
Finally, receiver operating characteristic curve (ROC) analysis was used to evaluate the discriminative ability of metric models in identifying AD patients under distinct APOE ε4 status. Model 1 was the combination of age, sex, years of education, and APOE ε4‐related alterations in dynamic metrics. Model 2 was the combination of age, sex, years of education, and APOE ε4‐related and cognition‐related alterations in dynamic metrics. Model 3 was the combination of age, sex, years of education, AD plasma biomarkers, and APOE ε4‐related and cognition‐related alterations in dynamic metrics. To prevent model overfitting, we conducted cross‐validation analysis with k‐fold cross‐validation (k = 5) (Poldrack et al. 2020; White and Power 2023). Specifically, the data were randomly divided into 5 subgroups. The holdout approach was then repeated 5 times, with one of the 5 subsets providing the test set and the other 4 subsets forming a training set each time. The performance estimation was averaged over all 5 trials to evaluate overall efficiency. All the above analyses were conducted in the R software (version 4.4.1).
Results
3
Demographic, Risk Factors, Cognitive, Pathologic, and Imaging Characteristics
3.1
Overall, as shown in Table 1, there were 38 (19.19%) and 65 (29.55%) APOE ε4 carriers in NC and AD groups, respectively. There were no group differences in sex distribution, plasma Aβ40, plasma NfL, TIV, or prevalence of hypertension, diabetes mellitus, and hyperlipidaemia across the four groups (all p > 0.05). Compared with the NC group, subjects with AD showed older age and lower education level (Bonferroni and Kruskal‐Wallis ANOVA corrected, p < 0.05). Furthermore, within the AD group, as compared to non‐carriers, APOE ε4 carriers showed worse orientation function, lower plasma Aβ42 and plasma Aβ42/40 ratio, higher plasma GFAP, and plasma p‐tau181 (Bonferroni and Kruskal‐Wallis ANOVA corrected, p < 0.05).
Alterations of Dynamic Local Metrics and Voxel‐Wise Concordance
3.2
Firstly, mixed effect analysis showed that there were significant group effects (i.e., effect of APOE ε4 allele) on dALFF (SD) in the right angular gyrus (ANG)/supramarginal gyrus (SMG), left occipital lobe (OL)/cuneus (CUN)/calcarine fissure and surrounding cortex (CAL) (Table 2 and Figure 1a), as well as dALFF (CV) in the right superior parietal gyrus (SPG)/precuneus (PCUN)/brodmann area 7 (Table 2 and Figure S1). There were significant group effects on dReHo (SD) in the right parietal lobe (PL)/SPG/brodmann area 7/PCUN (Table 2 and Figure 1d) and dGSCorr (Mean) in the triangular part of left inferior frontal gyrus (IF Gtriang) (Table 2 and Figure S1) as well. Secondly, brain regions with voxel‐wise concordance variability were located in the right caudate nucleus (CAU) and opercular part of left inferior frontal gyrus (IF Goperc) (Table 2 and Figure 1g). Thirdly, Table 2 exhibited cognition‐related alterations in dALFF in the right PCUN/brodmann area 7 and dfALFF in the left SPG/PCUN/brodmann area 7. Critically, APOE ε4 allele × cognitive status interaction emerged across groups.
*Brain regions with significant group effect differences in SD maps of dynamic metrics across the four groups. APOE ε4 allele led to significant differences in dALFF (SD) (a–c), dReHo (SD) (d–f), and voxel‐wise concordance (g–i) across the four groups (thresholds: For dALFF and dReHo, voxel‐level GRF < 0.001, cluster‐level GRF < 0.05, while for voxel‐wise concordance, voxel‐level GRF < 0.01, cluster‐level GRF < 0.05). The color bar indicated the T value. Warm colors indicated increased values, while cold colors indicated decreased values (Ɛ4 carriers versus non‐carriers, or AD versus NC). Age, sex, and years of education were used as covariates. *p < 0.05, **p < 0.01, **p < 0.001.
In post hoc analysis, APOE ε4 carriers with AD showed decreased dALFF (SD) values in the right ANG/SMG (Figure 1b), increased dALFF (SD) values in the left OL/CUN/CAL (Figure 1c) and increased dALFF (CV) values in the right SPG/PCUN/brodmann area 7 (Figure S1) relative to non‐carriers. Similarly, APOE ε4 carriers with AD showed decreased dReHo (SD) values in the right PL/SPG/PCUN/brodmann area 7 in comparison with non‐carriers (Figure 1e,f). We also observed decreased dGSCorr (Mean) values in left IF Gtriang among APOE ε4 carriers (Figure S1). In particular, relative to non‐carriers, APOE ε4 carriers exhibited decreased voxel‐wise concordance in the right CAU and left IF Goperc (Figure 1h,i) independent of cognitive status.
Clusters with significant group effects and condition effects at the window step of 2 TR and the window length of 40 TR were similar to those above (Tables S1 and S2, Figures S2 and S3).
Effect of
APOE ε4 Allele on Correlations Between Dynamic Rs‐fMRI Metrics and Cognition, Plasma AD Biomarkers
3.3
According to the stratified analysis, correlations between dynamic rs‐fMRI metrics and cognition, plasma AD biomarkers among subjects with AD were associated with APOE ε4 mutation status while controlling for age, sex, and years of education. As shown in Figure 2, there were positive associations between dALFF (SD) values in the left OL/CUN/CAL and MMSE scores (R = 0.245, p = 0.002, Figure 2a), dALFF (SD) values in the right ANG/SMG and naming ability (R = 0.244, p = 0.011, Figure 2c), dReHo (SD) values in the right SPG/brodmann area 7 and language function (R = 0.238, p = 0.014, Figure 2e), dReHo (SD) values in the right SPG/brodmann area 7 and delayed recall (R = 0.207, p = 0.033, Figure 2f) in APOE ε4 non‐carriers rather than carriers (all p > 0.05). Meanwhile, dALFF (SD) values in the right ANG/SMG were positively correlated with MoCA scores (R = 0.377, p = 0.006, Figure 2b) and orientation function (R = 0.405, p = 0.014, Figure 2d) in APOE ε4 carriers but not in non‐carriers.
Effect of APOE ε4 allele on correlations between values of SD maps of dynamic rs‐fMRI metrics and cognition in the AD group. APOE ε4 allele weakened correlations between dALFF (SD) values in the left OL/CUN/CAL and MMSE scores (a), dALFF (SD) values in the right ANG/SMG and naming ability (c), dReHo (SD) values in the right SPG/brodmann area 7 and language function (e), delayed recall (f). While APOE ε4 allele enhanced correlations between dALFF (SD) values in the right ANG/SMG and MoCA scores (b), orientation function (d). Age, sex, and years of education were used as covariates. APOE ε4 non‐carriers and carriers were presented as blue and red colors, respectively.
Critically, we observed negative associations between voxel‐wise concordance in the right CAU and MMSE scores (R = −0.303, p = 0.017, Figure 3a), MoCA scores (R = −0.304, p = 0.029, Figure 3b), attention (R = −0.314, p = 0.037, Figure 3c), language function (R = −0.317, p = 0.039, Figure 3d), orientation function (R = −0.346, p = 0.039, Figure 3e) but a positive association with plasma Aβ42 (R = 0.297, p = 0.025, Figure 3f) in APOE ε4 carriers with AD rather than non‐carriers.
Effect of APOE ε4 allele on correlations between voxel‐wise concordance in the right CAU and cognition, Aβ42 in the AD group. APOE ε4 enhanced the correlation between voxel‐wise concordance in the right CAU and MMSE scores (a), MoCA scores (b), attention (c), language function (d), orientation function (e), plasma Aβ42 (f). Age, sex, and years of education were used as covariates. APOE ε4 non‐carriers and carriers were presented as blue and red colors, respectively.
Most of above‐mentioned changes in correlations between dynamic metrics and cognition, Aβ42 in AD subjects within different APOE ε4 mutation status were also observed across multiple window parameters (Figure S4–S7).
Mediation Effect of Voxel‐Wise Concordance on Relationship Between Plasma Aβ42 and Language Function
3.4
Bootstrapped mediation analysis exhibited that voxel‐wise concordance in the right CAU partially mediated the negative association between plasma Aβ42 and language function in APOE ε4 carriers with AD (indirect effect = −0.0456, 95% CI [−0.1359, −0.0009], Figure 4), after adjustment for age, sex, and years of education. This mediation pattern remained consistent across varying window parameters (Figure S8).
Mediation effect of voxel‐wise concordance in the right CAU on relationship between Aβ42 and language function in APOE ε4 carriers with AD. Voxel‐wise concordance in the right CAU partly mediated the relationship between Aβ42 and language function. The red arrow represented negative correlation, and the yellow arrow represented positive correlation. Age, sex, and years of education were used as covariates.
Discriminative Performance of Combined Models for AD Classification
3.5
As shown in Table 3, the k‐fold cross‐validation confirmed superior diagnostic performance of combined models in APOE ε4 carriers than in non‐carriers, with higher values for all performance parameters.
Further cross‐validated confusion matrices showed that among APOE ε4 carriers, 89.23% of AD patients were correctly identified by Model 3, 10.77% were incorrectly classified as NC individuals, and 15.79% of the NC group was misclassified. However, Model 3 showed reduced efficacy in non‐carriers, with the true positive rate of 72.26%, the false positive rate of 27.74%, the true negative rate of 80.00%, and the false negative rate of 20.00%. Model 1 and Model 2 were similar to Model 3 (Figure S9).
Also, cross‐validated ROC curves (Figure 5) showed that among APOE ε4 carriers, the area under the curve (AUC) of Model 1 to discriminate AD patients from NC individuals was 0.78 (95% CI [0.69, 0.87]), the AUC of Model 2 was 0.88 (95% CI [0.81, 0.95]), and the AUC of Model 3 was up to 0.94 (95% CI [0.89, 0.98]). Performance was consistently lower in non‐carriers.
Cross‐validated ROC curves for classification between NC and AD. Combined models discriminated AD patients from NC individuals among APOE Ɛ4 non‐carriers (a) and APOE Ɛ4 carriers (b). Yellow color presented Model 1, blue color presented Model 2 and red color presented Model 3.
Discussion
4
To the best of our knowledge, this is the first study to comprehensively examine alterations in dynamic local spontaneous brain activity and functional integration associated with APOE ε4 allele in AD. We found that APOE ε4 allele impacted dALFF, dReHo, and dGSCorr including frontal, parietal, and occipital lobes, as well as voxel‐wise concordance in the right CAU and left IF Goperc. The APOE ε4 allele influenced correlations of dynamic functional activities with both cognition and Aβ pathology in AD. Furthermore, the voxel‐wise concordance in the right CAU partially mediated the relationship between Aβ42 and language function in APOE ε4 carriers with AD. Finally, the dynamic combined model could effectively identified cognitive status, particularly in APOE ε4 carriers. This in‐depth analysis of dynamic local activity, concordance, and plasma AD pathology may provide novel insights into the mechanisms of APOE ε4 in AD pathology.
The present study revealed that APOE ε4 carriers with AD exhibited spatial overlap of dALFF and dReHo alterations in the right SPG/PCUN/brodmann area 7, suggesting increased variability in neural activity intensity and decreased stability of synchronous activity in adjacent voxels in these regions. Previous studies have observed abnormal ALFF and ReHo in these regions in amnestic MCI (Pan et al. 2017; Zhen 2018; Zhong et al. 2022) and subjective cognitive decline (Li et al. 2021), further indicating that aberrant spontaneous brain activity in the right SPG/PCUN/brodmann area 7 may be associated with cognitive decline. Our further analysis showed that dReHo in the right SPG/brodmann area 7 positively correlated with language function and delayed recall in APOE ε4 non‐carriers but not in carriers. Given that SPG is associated with voice severity in essential tremor (De Lima Xavier and Simonyan 2020), auditory integration for vocal pitch regulation (Chen et al. 2021), and long‐term delayed recall (Ma et al. 2023), we speculate that APOE ε4 allele may cause inflexible connectivity between the right SPG/PCUN/brodmann area 7 and neighboring brain regions during the integration, processing, and analysis of cognitive information in AD patients.
Nevertheless, the anatomical distributions of changes in dynamic local metrics were not completely overlapping, indicating that APOE ε4 allele may affect brain activity by physiological complementarity in different regions (Chen et al. 2023). First, the decreased dALFF (SD) of the right ANG/SMG in APOE ε4 carriers suggested that APOE ε4 allele weakens the stability of neuronal activity in the inferior parietal lobule (IPL). The IPL plays a key role in object recognition, social cognition, language function, and orientation (Binder and Desai 2011; Peer et al. 2015). Wei et al. also reported decreased mean dALFF in the IPL in subjective cognitive decline (Wei et al. 2022). Since higher dALFF stability contributes to flexible brain adaptability to environmental demands (Liao et al. 2019), our partial correlation analysis indicated that worse general cognition and orientation function in APOE ε4 carriers with AD likely result from inflexible neuronal activity in the right IPL during time‐varying intrinsic processing. Second, dALFF stability in the left OL/CUN/CAL increased in APOE ε4 carriers. Given the critical role of the OL in various higher cognitive functions (Bettencourt and Xu 2016; Snytte et al. 2022), we speculated that APOE ε4 allele may cause a compensatory increase in neural flexibility that disrupts the relationship between neural stability and general cognition in this region. These regions represent potential therapeutic targets for AD patients carrying APOE ε4 allele.
The voxel‐wise concordance was then used to quantify the harmony of dynamic local metrics during state transitions. Results revealed decreased voxel‐wise concordance in the left IF Goperc and right CAU in APOE ε4 carriers, potentially due to inconsistent synchronization among dynamic local metrics. The IF Goperc constitutes part of Broca's area and serves as the language network hub connecting diverse cortical, subcortical, and cerebellar regions (Bulut 2022). A rs‐fMRI study based on visibility graph demonstrated that left IF Goperc plays a significant role in ad (Gao et al. 2020 ). However, our results indicated that decreased concordance in the left IF Goperc associated with APOE ε4 did not significantly affect cognitive function in AD patients.
Our correlation analysis suggested that the APOE ε4 allele strengthened the correlation between functional integration in the right CAU and cognition. As a key component of the cortico‐basal ganglia‐thalamo‐cortical circuit (Kim, Kim, et al. 2022), the CAU affects multiple cognitive functions including attention and language function (Herman et al. 2020; Tomasi and Volkow 2012). Prior studies have observed functional and structural caudate alterations in APOE ε4 carriers. For example, reduced hippocampal‐caudate FC in APOE ε4 carriers correlates significantly with episodic memory impairment (Li et al. 2014). Additionally, the APOE ε4 allele accelerates gray matter atrophy in the CAU among patients with MCI to AD conversion (Spampinato et al. 2011). Unexpectedly, we observed that higher voxel‐wise concordance was associated with worse cognition in APOE ε4 carriers with AD, which may be explained by synchronous decreases in all dynamic local metrics as AD severity progresses.
Furthermore, we found that voxel‐wise activity in the right CAU was positively associated with plasma Aβ42 exclusively in APOE ε4 carriers. Previous studies have indicated that APOE ε4 carriers exhibited accelerated Aβ deposition and significant degeneration of serotonergic synapses in the ventromedial caudate nucleus (Kim, Chun, et al. 2022; Postupna et al. 2017; Ichimata et al. 2024). Notably, Aβ positivity in the caudate nucleus was significantly correlated with global cognitive decline in APOE ε4 carriers (Brugulat‐Serrat et al. 2023), aligning with our findings. Our mediation analysis further indicated that plasma Aβ42 acts as an upstream driver of functional alterations in the right CAU. Researchers have reported an interactive effect between smoking and functional connectivity of the right CAU with the bilateral anterior cingulate cortex on language function in MCI patients due to ad (Qiu et al. 2022 ). Furthermore, the bilateral CAU was proven to be associated with advanced semantic fluency strategies, particularly during the MCI stage (Ahn et al. 2022; Kwak et al. 2022). Integrating our mediation model results with these prior findings, we propose that the right CAU critically influences language function in AD. However, spatial relationships between voxel‐wise concordance and Aβ deposition require validation via positron emission tomography, and causal relationships among these factors need longitudinal investigation.
Ultimately, ROC analysis demonstrated that these dynamic metrics could serve as potential early imaging biomarkers and prognostic markers for AD, and the corresponding brain regions may represent effective targets for neuroregulatory therapy in early AD.
Several limitations should be mentioned in the present study. First, recent studies suggest that APOE ε4 homozygotes may have distinct functional and pathological trajectories, requiring separate analysis from heterozygotes (Fortea 2024). Due to the substantial sample size disparity between these groups, we did not conduct subgroup analysis, limiting this study's applicability for screening or diagnostic purposes. We will improve this shortcoming in the following research. Second, given that dfALFF, dReHo, and dGSCorr are more developed in characterizing local intrinsic activity, only these three dynamic local metrics were applied to calculate voxel‐wise concordance. Nevertheless, the optimal combination for comprehensive characterization remains undetermined. Third, as a cross‐sectional study with a limited sample size, we are continuing to recruit new subjects and follow up with them to validate our findings.
Conclusion
5
In conclusion, our findings indicated that the APOE ε4 allele broadly influenced dynamic local spontaneous brain activity and functional integration in AD. It also modified the relationships between alterations in dynamic metrics and both cognition and plasma AD biomarkers. Importantly, voxel‐wise concordance in the right CAU partially mediated the relationship between plasma Aβ and language function. Our research highlights the effects of APOE ε4 on dynamic metric‐cognition relationships in AD, providing new insights into pathological mechanisms. Moreover, functionally altered regions may suggest therapeutic targets for AD patients with APOE ε4 allele. However, the cross‐sectional design and lack of subgroup analysis limit the application potential of this study in screening or diagnosis.
Author Contributions
Yi Tan: conceptualization, methodology, formal analysis, data curation, and writing – original draft; Dan Yang: conceptualization, methodology, and writing – review and editing; Zhihong Ke: data curation; Zheqi Hu: data curation; Wenting Song: data curation and formal analysis; Limoran Tang: data curation; Zhixin Zhou: data curation; Yuting Mo: data curation; Lili Huang: data curation; Yun Xu: supervision, project administration, and funding acquisition.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Data S1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahn, H. , D. Yi , K. Chu , et al. 2022. “Functional Neural Correlates of Semantic Fluency Task Performance in Mild Cognitive Impairment and Alzheimer's Disease: An FDG‐PET Study.” Journal of Alzheimer's Disease 85, no. 4: 1689–1700. 10.3233/JAD-215292.PMC 921029134958036 · doi ↗ · pubmed ↗
- 2Albert, M. S. , S. T. De Kosky , D. Dickson , et al. 2011. “The Diagnosis of Mild Cognitive Impairment due to Alzheimer's Disease: Recommendations From the National Institute on Aging‐Alzheimer's Association Workgroups on Diagnostic Guidelines for Alzheimer's Disease.” Alzheimer's & Dementia 7, no. 3: 270–279. 10.1016/j.jalz.2011.03.008.PMC 331202721514249 · doi ↗ · pubmed ↗
- 3Baxter, L. C. , M. Limback‐Stokin , K. J. Patten , et al. 2023. “Hippocampal Connectivity and Memory Decline in Cognitively Intact APOE ε4 Carriers.” Alzheimer's & Dementia 19, no. 9: 3806–3814. 10.1002/alz.13023.PMC 1110501836906845 · doi ↗ · pubmed ↗
- 4Bettencourt, K. C. , and Y. Xu . 2016. “Decoding the Content of Visual Short‐Term Memory Under Distraction in Occipital and Parietal Areas.” Nature Neuroscience 19, no. 1: 150–157. 10.1038/nn.4174.26595654 PMC 4696876 · doi ↗ · pubmed ↗
- 5Binder, J. R. , and R. H. Desai . 2011. “The Neurobiology of Semantic Memory.” Trends in Cognitive Sciences 15, no. 11: 527–536. 10.1016/j.tics.2011.10.001.22001867 PMC 3350748 · doi ↗ · pubmed ↗
- 6Blanchard, J. W. , L. A. Akay , J. Davila‐Velderrain , et al. 2022. “APOE 4 Impairs Myelination via Cholesterol Dysregulation in Oligodendrocytes.” Nature 611, no. 7937: 769–779. 10.1038/s 41586-022-05439-w.36385529 PMC 9870060 · doi ↗ · pubmed ↗
- 7Brugulat‐Serrat, A. , G. Sánchez‐Benavides , R. Cacciaglia , et al. 2023. “APOE‐ε4 Modulates the Association Between Regional Amyloid Deposition and Cognitive Performance in Cognitively Unimpaired Middle‐Aged Individuals.” EJNMMI Research 13, no. 1: 18. 10.1186/s 13550-023-00967-6.36856866 PMC 9978048 · doi ↗ · pubmed ↗
- 8Bulut, T. 2022. “Meta‐Analytic Connectivity Modeling of the Left and Right Inferior Frontal Gyri.” Cortex 155: 107–131. 10.1016/j.cortex.2022.07.003.35985123 · doi ↗ · pubmed ↗