Altered Bcl-2/Caspase signaling and hypoxia-induced apoptosis in primary human aniridia limbal stromal cells, in CoCl2 mediated hypoxic stress, in vitro
Shanhe Liu, Shuailin Li, Shao-Lun Hsu, Fabian N. Fries, Zhen Li, Swarnali Kundu, Berthold Seitz, Maryam Amini, Shweta Suiwal, Julia Zimmermann, Simon Trusen, Tanja Stachon, Nóra Szentmáry

TL;DR
Aniridia limbal stromal cells show higher apoptosis and altered Bcl-2/Caspase signaling under hypoxic stress compared to healthy cells.
Contribution
This study identifies specific gene and protein expression changes in aniridia limbal cells under hypoxia, offering insights into disease mechanisms.
Findings
AN-LSCs showed significantly higher apoptosis rates than healthy LSCs under hypoxic stress.
CoCl2 treatment increased apoptosis in AN-LSCs and altered expression of Bcl-2, Caspase, and CDKN1B.
TNF-α protein levels in supernatant increased in AN-LSCs under hypoxia.
Abstract
The aim of this study was to investigate apoptosis in primary aniridia limbal stromal cells (LSCs) and to assess changes in the expression of genes and proteins associated with the apoptotic pathway in response to cobalt chloride (CoCl2)-induced hypoxic stress, in vitro. Primary human limbal stromal cells were isolated from the limbal region of both aniridia (AN-LSCs; n = 8) and healthy (LSCs; n = 8) donors. The cells were treated with 0 µM, 50 µM, and 75 µM CoCl2 for 48 hours. Apoptosis in each group was assessed by Flow cytometry (FC). The expression levels of apoptosis-related genes, including CASP 3/7/8/9/10, BCL2, BID, BAX, CDKN1A (p21), CDKN1B (p27), TNFα, XIAP, and BIRC5 (Survivin), were measured by qPCR. Protein level of these markers was analyzed by FC. TNFα protein expression in the supernatant was quantified using ELISA. Flow cytometry analysis revealed a significantly…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
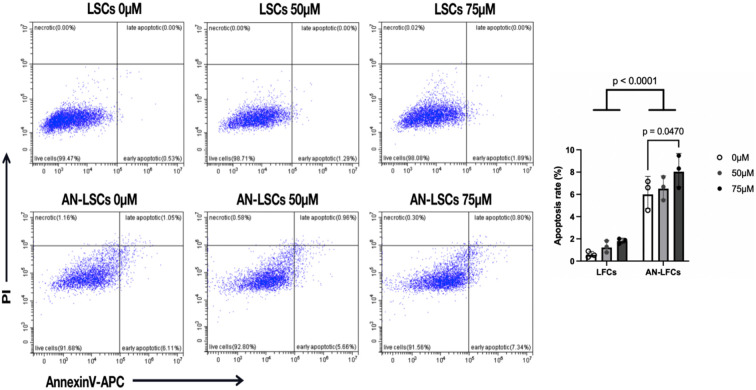 Fig 1
Fig 1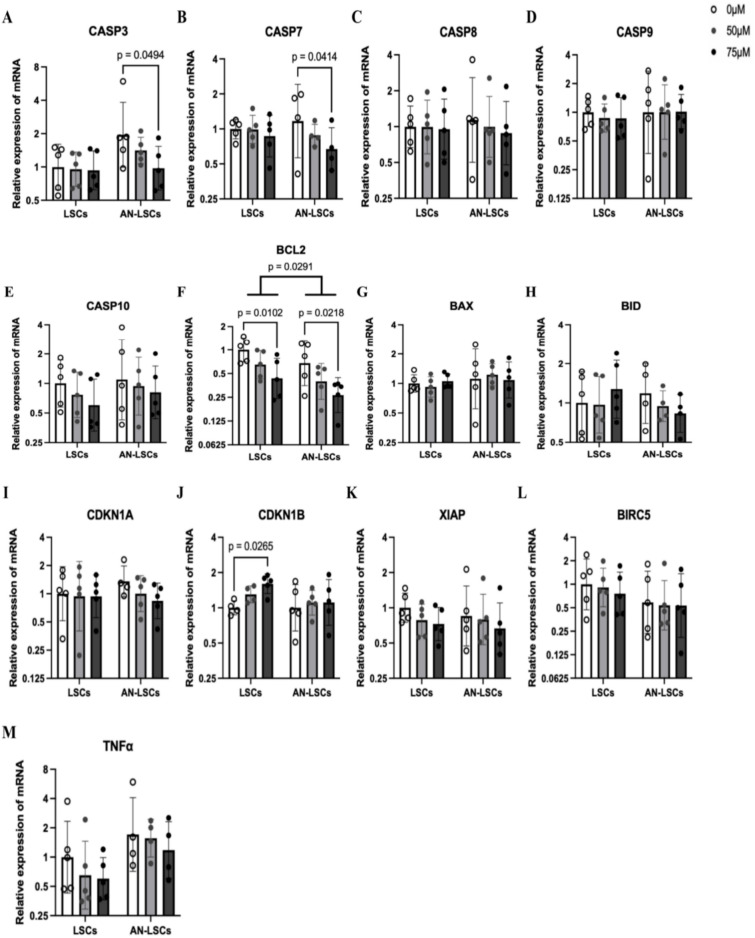 Fig 2
Fig 2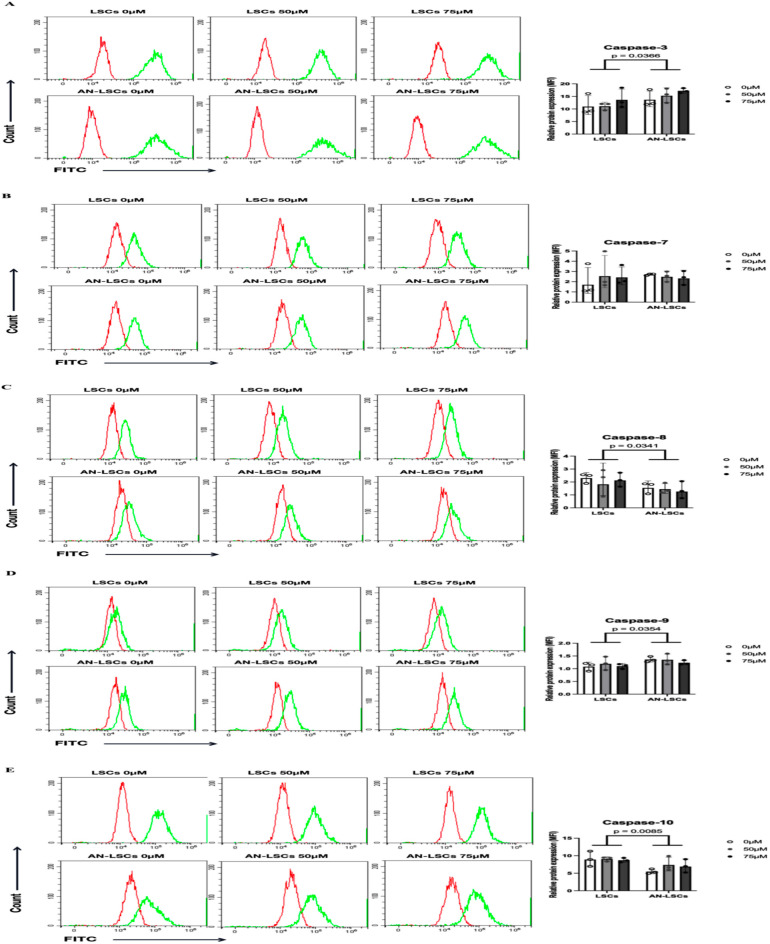 Fig 3
Fig 3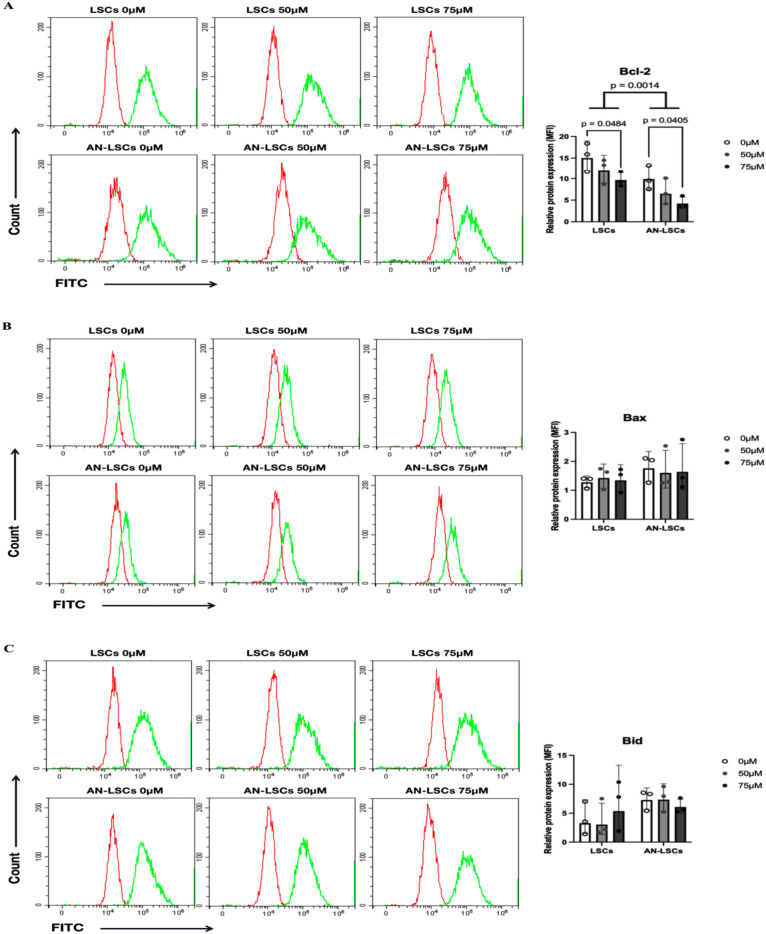 Fig 4
Fig 4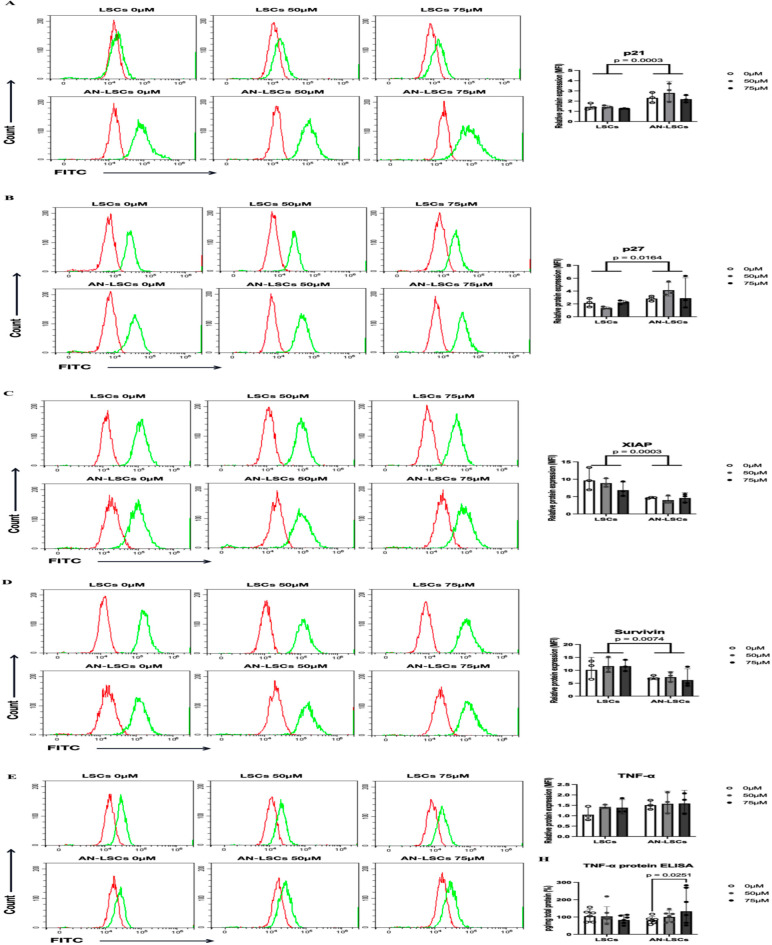 Fig 5
Fig 5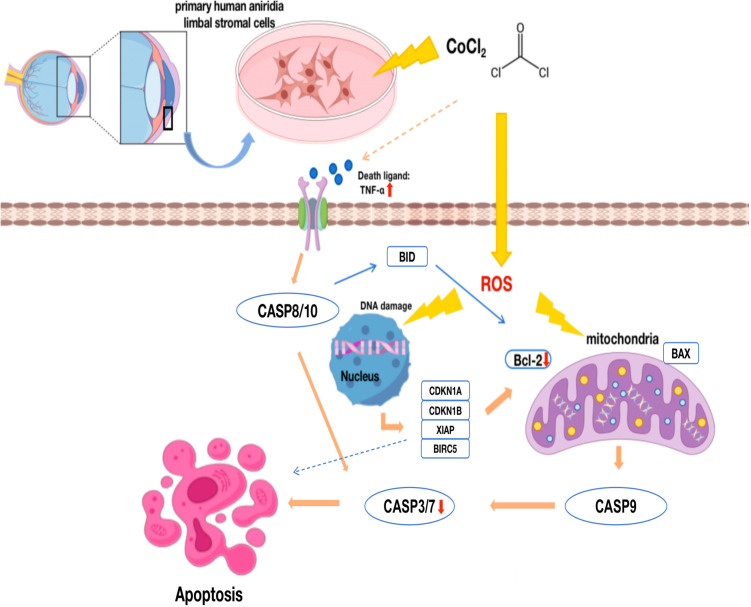 Fig 6
Fig 6- —Dr. Rolf M. Schwiete Foundation
- —Dr. Rolf M. Schwiete Foundation
- —Dr. Rolf M. Schwiete Foundation
- —Dr. Rolf M. Schwiete Foundation
- —Dr. Rolf M. Schwiete Foundation
- —Dr. Rolf M. Schwiete Foundation
- —Dr. Rolf M. Schwiete Foundation
- —Dr. Rolf M. Schwiete Foundation
- —Dr. Rolf M. Schwiete Foundation
- —Dr. Rolf M. Schwiete Foundation
- —Dr. Rolf M. Schwiete Foundation
- —http://dx.doi.org/10.13039/501100004543China Scholarship Council
- —http://dx.doi.org/10.13039/501100004543China Scholarship Council
- —http://dx.doi.org/10.13039/501100004543China Scholarship Council
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCorneal Surgery and Treatments · Marine Invertebrate Physiology and Ecology · Ocular Infections and Treatments
Introduction
Congenital aniridia is a rare panocular condition characterized by the partial or complete absence of the iris (prevalence approximately 1:40,000–1:100,000), with common complications including aniridia-associated keratopathy (AAK), secondary glaucoma, juvenile cataract, macular and optic nerve head hypoplasia [1–3]. AAK is one of the leading causes of progressive vision loss in congenital aniridia. It is characterized by progressive limbal stem cell deficiency, corneal scarring, pannus formation and corneal neovascularization [3]. Abnormal corneal epithelial cell differentiation and migration, altered gene expression in aniridia limbal stromal cells, increased sensitivity to oxidative stress, impaired wound healing, and elevated apoptosis within the limbal niche may all contribute to the development and progression of AAK [4,5]. Interestingly, in cell culture, the viability, proliferation, and migration of aniridia-derived limbal stromal cells did not differ significantly from those of healthy controls [6,7].
Oxidative stress refers to the excessive production of highly active intracellular molecules such as reactive oxygen species (ROS) and an imbalance between oxidative and antioxidative pathways. This imbalance often occurs when the organism is exposed to environmental stressors or chemical agents and can lead to cellular damage. ROS not only contribute to oxidative damage but also function as molecular signals, activating various stress-sensitive intracellular pathways and initiating apoptosis [8]. One such chemical agent, Cobalt chloride (CoCl_2_), is widely used as a hypoxia mimetic and has been shown to induce ROS production, apoptosis, and transcriptional changes in genes such as hypoxia-inducible factor-1α (HIF-1α), p53, and p21 [9–11]. Studies have demonstrated that CoCl_2_ induces apoptosis in several types of stem cells [12,13]. Given that the cornea is continuously exposed to oxidative stress—particularly through UV light—ROS-related effects may influence cellular metabolism, migration, and overall corneal function [3,5,14]. In aniridia mouse models, the corneal epithelium has been shown to be particularly vulnerable to oxidative stress, resulting in increased apoptotic cell death [15,16].
Apoptosis is a form of programmed cell death through which cells undergo self-regulated elimination. This process is essential for maintaining tissue homeostasis and for the removal of damaged or abnormal cells. There are two main classical apoptotic pathways: the extrinsic and intrinsic pathways. The extrinsic pathway is initiated by the activation of cell surface death receptors (e.g., Fas, TNF receptors), which trigger apoptosis through the activation of effector caspases, such as Caspase-3. In contrast, the intrinsic pathway is activated by intracellular stressors—including DNA damage, hypoxia, and other cellular insults. This leads to the formation of apoptotic bodies, activation of effector caspases, and ultimately cell death. Although distinct, both pathways are interconnected and function in coordination, regulated by a complex network of signaling molecules to ensure that apoptosis occurs only under appropriate physiological or pathological conditions [17].
Stachon et al. identified alterations in apoptosis-related pathways in the protein profiles of conjunctival impression cytology samples from individuals with aniridia [18]. In addition, in aniridia limbal stromal cells, alterations of miRNA levels regulating apoptotic processes have been identified [19]. However, to the best of our knowledge, apoptotic signaling in limbal stromal cells (LSCs) from patients with congenital aniridia has not yet been investigated.
The purpose of this study was to explore apoptotic pathways in primary aniridia-derived LSCs under CoCl_2_-induced hypoxic stress in vitro. Our goal was to identify potential therapeutic targets within the apoptotic cascade that could be leveraged to favorably modulate the wound healing response.
Materials and methods
Ethics approval and consent to participate
This study was approved by the Ethics Committee of Saarland/ Germany (No. 178/22) and followed the rules of the Declasaion of Helsinki. All patients provided their consent in written form. The recruitment period of the prospective study started from 07.11.2022 to 15.12.2024.
Cell culture
Aniridia limbal samples have been obtained from patients of the Department of Ophthalmology, Saarland University Medical Center, Homburg/Saar, Germany. Before surgery, all aniridia patients underwent a slit-lamp examination to grade AAK according to Lagali et al [20,21]. Control limbal biopsies were obtained from corneal donors of the LIONS Cornea Bank Saar-Lor-Lux, Trier/Westpfalz, Homburg/Saar, Germany.
1.5 mm limbal biopsies of 8 congenital aniridia eyes (age 32.50 ± 17.07 (2–50) years; 3 (37.5%) males) and 12 healthy donor eyes (age 81.17 ± 7.62 (64–90) years; 6 (50%) males) have been used for our experiments. Tables 1 and 2 provide details about all samples.
Table 1: Detailed information of the used healthy corneal donors.
Table 2: Detailed information of the included congenital aniridia subjects. The aniridia-associated keratopathy (AAK) grade according to Lagali et al [23,24] is also included in the Table.
The extracted limbal biopsies were then placed in keratinocyte growth medium (KGM3, PromoCell) containing collagenase A (1 mg/ml) (Roche Diagnostic GmbH, Mannheim, Germany, No.10103578001) and were incubated at 37°C for 24 hours.
Isolation of primary LSCs has been performed as described by Chai et al. [22]. The suspensions were passed through 40 µm Flowmi cell strainers (No. H13680-0040, Bel-Art, Wayne, USA) into a 24-well plate to separate incompletely dissociated cells.
For LSCs cultures, the strainer was rinsed with 500 µl PBS followed by 500 µl of 0.05% trypsin-EDTA, and incubated for 5 minutes to disaggregate cell clusters. The enzyme activity was then halted by adding DMEM supplemented with 5% FCS. The resulting filtrate was centrifuged at 1500g for 5 minutes to pellet the LSCs, and the supernatant was discarded. The cells were then seeded in a 6-well plate using DMEM supplemented with 5% FCS. The cells were maintained at 37°C with 95% relative humidity and 5% CO_2_, with the medium being refreshed every 2–3 days until the LSCs reached confluence.
CoCl2 treatment
LSCs were grown until reaching 80% confluence, before experiments. After 24 hours, the medium was substituted with DMEM solution that included CoCl_2_ (Sigma-Aldrich® GmbH, Geisenheim, Germany, No. 60818). This solution was left for 48 hours to create a hypoxic environment with CoCl_2_ doses of 0 µM, 50 µM, and 75 µM, respectively.
Apoptosis
Apoptosis was detected using an Annexin V apoptosis detection kit (Invitrogen, Germany, No.2734867). Cells were trypsinized, centrifuged, and resuspended in 1xbinding buffer. The samples were incubated with 5 µL of Annexin V-APC at room temperature for 15 minutes, followed by the addition of 5 µL of propidine iodide (PI) and subsequent analysis by flow cytometry. Annexin V-APC and PI fluorescence were detected in the R660 nm and B585 nm channels, respectively. Non-apoptotic and apoptotic cells were distinguished based on Annexin V-APC and PI staining. Non-apoptotic cells showed no staining with either reagents. Apoptotic cells exhibited strong red (APC) fluorescence and low to moderate blue (PI) fluorescence, indicating early and late apoptosis. The increased permeability of late apoptotic cells to PI was due to compromised plasma membrane integrity.
The results were expressed as the percentage of apoptotic cells relative to the total cell population detected. We represented data as mean±SD from three independent experiments.
RNA measurement
The RNA was extracted following the manufacturer’s instructions using the NORGEN RNA Purification Plus Micro Kit. The quantification of proteins and RNA was performed using the Bradford method and a UV/VIS spectrophotometer (Analytik Jena AG, Jena, Germany), respectively. The samples were then kept at a temperature of −80 °C. The synthesis of cDNA was performed using the OneTaq RT-PCR kit (New England Biolabs Inc. Ipswich, USA), 500 ng of total RNA per sample was used as the template. Synthesized cDNA was stored at −20 °C.
Quantitative PCR
Table 3 provides a summary of the primers used for qPCR obtained from Qiagen GmbH (Hilden, Germany). As the manufacturer’s guidelines, the samples were tested using 1 µl of the appropriate primer solution, 5 µl of SYBR Green Mix (Vazyme, Nanjing, China), 3 µl of nuclease-free water, and 1 µl of cDNA. Each gene was measured in duplicate. The PCR Thermocycler QuantStudio 5 Real-Time PCR System, manufactured by ThermoFisher ScientificTM GmbH in Dreieich, Germany, was utilized. The amplification conditions consisted of a denaturation step at 95 °C for 10 seconds, followed by an annealing step at 60 °C for 30 seconds, and a final extension step at 95 °C for 15 seconds. This cycle was repeated 40 times. The cycling threshold (Ct) obtained from the measurements was adjusted by normalizing it to the average values of β-glucuronidase (GUSB) to calculate ΔCt (ΔCt = individual Ct value – Ct value of GUSB). ΔΔCt was then defined as the average ΔCt minus the individual ΔCt. The geometric mean was computed by taking the multiplicative change in ΔΔCt (geometric mean = 2 ^ΔΔCt^), which represents the difference between the Ct value of the target sample and the average Ct value of the reference gene in a squared multiple manner.
Table 3: Primer pairs used for qPCR.
Flow cytometry
For LSCs and AN-LSCs, flow cytometry (FC) analysis was performed in three independent experiments. Cells were detached with trypsin without EDTA (Merck, USA, NO. T4424), counted, washed, and incubated with the appropriate antibodies. For staining with all markers, antibodies not conjugated to a fluorescent dye were used, and the secondary antibody (Antibody online, No. ABIN101988. Biotechne, No. F0118) was only used as a staining control. Stained cells were analysed immediately using a CytoFLEX flow cytometer (Beckman Coulter) and further analysed using CytExpert 2.6 software. Table 4 provides details about all antibodies.
Table 4: Antibodies used for flow cytometry.
ELISA
In this study, TNFα protein level in the supernatant of LSCs and AN-LSCs was also quantified using the DuoSet® ELISA Kit (R&D Systems, Bio-Techne, Minneapolis, USA, NO. DY210). The assay was conducted in accordance with the manufacturer’s protocol, and each measurement was performed in duplicate. Absorbance was recorded at 450 nm using the Tecan Infinite F50 Absorbance Microplate Reader (Tecan Group AG, Männedorf, Switzerland) to determine protein concentrations. To ensure accurate quantification, the obtained data were normalized against the total protein content.
Statistical analysis
Statistical analyses were performed using GraphPad Prism 9.0. Two-way ANOVA followed by Dunnett’s multiple comparisons has been used. Quantitative PCR data were presented as the geometric mean with geometric standard deviation, while FC data were expressed as mean±standard deviation (SD). P-values below 0.05 were considered statistically significant.
Results
The apoptotic ratio of cells
Flow cytometry revealed a significantly higher apoptosis rate in AN-LSCs, than in LSCs (p < 0.0001). In AN-LSCs, 75 µM CoCl_2_ treatment significantly increased apoptosis rate, compared to untreated AN-LSCs (p = 0.0470) (Fig 1).
Apoptosis rates in limbal stromal cells (LSCs) and Aniridia-LSCs (AN-LSCs), after treatment with 0 µM, 50 µM and 75 µM Cobalt chloride (CoCl2).LSCs and AN-LSCs were treated with CoCl2 (0, 50 and 75 µM) for 48 h and stained with Annexin V-APC/PI before flow cytometry analysis. Data represent mean±SD from three independent experiments. Two-way ANOVA followed by Dunnett’s test was used, significant p values are indicated. Apoptosis rates were significantly higher in AN-LSCs compared to LSCs (p < 0.0001). Additionally, treatment with 75 µM CoCl2 significantly increased apoptosis in AN-LSCs compared to untreated controls (p = 0.0470).
mRNA levels of apoptosis-related markers
In AN-LSCs, BCL2 mRNA levels were significantly downregulated, compared to LSCs (p = 0.0291). In LSCs, 75 µM CoCl_2_ treatment significantly downregulated BCL2 mRNA levels (p = 0.0102) and upregulated CNKN1B mRNA levels (p = 0.0265). In AN-LSCs, 75 µM CoCl_2_ treatment downregulated CASP3 (p = 0.0494), CASP7 (p = 0.0414) and BCL2 mRNA levels (p = 0.0218). Beyond these findings, there was no significant difference in the expression of other genes across different concentrations of CoCl_2_ treatment (Fig 2).
mRNA levels of CASP family markers (A-E), BCL2 family markers (F-H) and other apoptosis-related markers (I-M) in limbal stromal cells (LSCs) and in Aniridia-LSCs (AN-LSCs), after treatment with 0 µM, 50 µM and 75 µM Cobalt chloride (CoCl2).CASP3, CASP7, CASP8, CASP9 and CASP10 mRNA levels (A-E), BCL2, BAX and BID mRNA levels (F-H) and CDKN1A, CDKN1B, TNFα, XIAP and BIRC5 mRNA levels (I-M) are shown. Values are expressed on a logarithmic scale (Log 2) with geometric mean±geometric standard deviation. Two-way ANOVA followed by Dunnett’s test was used, significant p values are indicated. BCL2 mRNA levels were significantly higher in AN-LSCs compared to LSCs (p = 0.0291). In LSCs, BCL2 mRNA level significantly decreased (p = 0.0102) and CDKN1B level significantly increased using 75 µM CoCl2 treatment. In addition, in AN-LFCs, 75 µM CoCl₂ treatment significantly decreased CASP3, CASP9 and BCL2 mRNA levels (p = 0.0494; p = 0.0414; p = 0.0218).
Protein levels of apoptosis-related markers
Compared to LSCs, the protein levels of Caspase-3 (p = 0.0366), Caspase-9 (p = 0.0354), p21 (p = 0.0003) and p27 (p = 0.0164) were significantly upregulated, while Caspase-8 (p = 0.0341), Caspase-10 (p = 0.00085), Bcl-2 (p = 0.0014), XIAP (p = 0.0003) and Survivin (p = 0.0074) were significantly downregulated in AN-LSCs (Figs 3–5).
Proteins levels of Caspase family markers in limbal stromal cells (LSCs) and in Aniridia-LSCs (AN-LSCs), after treatment with 0 µM, 50 µM and 75 µM Cobalt chloride (CoCl2), using flow cytometry (A-E).Data represent mean±SD from three independent experiments. Two-way ANOVA followed by Dunnett’s test was used, significant p values are indicated. Protein levels of Caspase-3, Caspase-7, Caspase-8, Caspase-9, and Caspase-10 are shown. Representative histograms show primary antibody staining in green, while staining with the corresponding secondary antibody alone (negative control) is shown in red. MFI: mean fluorescence intensity, normalized to the secondary antibody control. In AN-LSCs, Caspase-3 and Caspase-9 protein levels were significantly higher and Caspase-8 and Caspase-10 protein levels were significantly lower than in LSCs (p≤0.0366). Nevertheless, there was no significant difference in protein levels of Caspase family members in any of the analysed subgroups, using different CoCl2 treatment.
Protein levels of Bcl-2 family markers in limbal stromal cells (LSCs) and in Aniridia-LSCs (AN-LSCs), after treatment with 0 µM, 50 µM and 75 µM Cobalt chloride (CoCl2), using flow cytometry (A-C).Data represent mean±SD from three independent experiments. Two-way ANOVA followed by Dunnett’s test was used, significant p values are indicated. Bcl-2, Bax and Bid protein levels are shown. Representative histograms show primary antibody staining in green, while staining with the corresponding secondary antibody alone (negative control) is shown in red. MFI: mean fluorescence intensity, normalized to the secondary antibody control. Bcl-2 protein level was significantly lower in AN-LSCs, than in LSCs (p = 0.0014). In addition, 75 µM CoCl₂ treatment significantly decreased Bcl-2 protein level both in LSCs and AN-LSCs (p = 0.0484; p = 0.0405).
Protein levels of other apoptosis-related markers in limbal stromal cells (LSCs) and in Aniridia-LSCs (AN-LSCs), after treatment with 0 µM, 50 µM and 75 µM Cobalt chloride (CoCl2), using flow cytometry (A-E) and ELISA (H).Data represent mean±SD from three independent experiments. Two-way ANOVA followed by Dunnett’s test was used, significant p values are indicated. p21, p27, XIAP, TNF-α, and Survivin protein levels are shown. Representative histograms show primary antibody staining in green, while staining with the corresponding secondary antibody alone (negative control) is shown in red. MFI: mean fluorescence intensity, normalized to the secondary antibody control. In AN-LSCs, p21 and p27 protein levels were significantly higher (p = 0.0003; p = 0.0164) and XIAP and Survivin protein levels were significantly lower (p = 0.0003; p = 0.0074), than in LSCs, using flow cytometry. In addition, using ELISA, in cell culture supernatant of AN-LSCs, TNF-α protein levels were significantly higher following 75 µM CoCl2 treatment, than in untreated controls (p = 0.0251).
In LSCs and in AN-LSCs, 75 µM CoCl_2_ treatment significantly downregulated Bcl-2 protein levels (p = 0.0484; p = 0.0405) (Fig 4). In the cell culture supernatant, TNF-α protein levels increased significantly (p = 0.0251) (Fig 5).
Discussion
In this study, we systematically identified significant dysregulation of apoptosis in corneal limbal stromal cells from patients with congenital aniridia, along with their distinct response patterns to hypoxic stress. Firstly, our study revealed that AN-LSCs exhibit a significantly higher apoptotic rate than healthy LSCs, accompanied by a significant upregulation of pro-apoptotic proteins (Caspase-3/9, p21/p27) and a widespread downregulation of anti-apoptotic molecules (Bcl-2, XIAP, Survivin). Secondly, we observed that 75 µM CoCl_2_-induced hypoxic environment further exacerbated apoptosis in AN-LSCs, while LSCs exhibited only downregulation of BCL2 mRNA and protein levels. Notably, AN-LSCs showed unexpected downregulation of pro-apoptotic genes such as CASP 3/7 and BCL2 mRNA levels after hypoxia, suggesting that there may be a compensatory feedback mechanism for the regulation of their apoptotic pathway. Finally, we also found that the changes in mRNA and protein levels were not completely consistent, suggesting that post-transcriptional modification or protein stability may play an important role in the apoptotic regulation of limbal stromal cells in AAK. This could be observed regarding CASP3 mRNA downregulation in AN-LSCs after hypoxia, although its basal protein levels were significantly increased.
Previous studies have shown that dysfunction or depletion of corneal limbal epithelial stem cells and limbal stromal cells leads to impaired renewal of the corneal epithelium, resulting in a range of ocular surface disorders [23,24]. The significant downregulation of Bcl-2 and XIAP, accompanied by the activation of Caspase-3 and 9, may contribute to the depletion of corneal limbal stromal cells via mitochondria-dependent apoptotic pathways in AN-LSCs [25]. Interestingly, in AN-LSCs, CoCl_2_ treatment suppressed some pro-apoptotic genes (CASP3/7 and BCL2 mRNA) although it further elevated the apoptosis rate. This paradoxical phenomenon may result from negative feedback regulatory mechanisms activated in cells undergoing prolonged apoptosis, such as cell cycle arrest mediated by p21/p27, which serves to limit further amplification of apoptotic signaling [26,27]. Liu et al. reported in their study on herpetic eye disease that inflammation can cause significant damage to the corneal limbal microenvironment [28]. In the present study, the marked increase in TNF-α protein levels in the supernatant of AN-LSCs following hypoxic stress suggests that the inflammatory microenvironment exacerbated changes through the extrinsic apoptotic pathway involving Caspase-8 and 10. These findings provide experimental support for the hypothesis that inflammation-induced apoptosis contributes to the pathogenesis of AAK (Fig 6).
Schematic representation of the CoCl2-mediated apoptotic pathway in primary human aniridia limbal stromal cells (AN-LSCs).In AN-LSCs, CoCl2 induces a hypoxic environment and stimulates reactive oxygen species (ROS) production, which in turn affects mitochondrial function, causes DNA damage, and triggers apoptosis. This is accompanied by differential expression of related genes, including downregulation of the anti-apoptotic Bcl-2 and the pro-apoptotic CASP3 and CASP7. CoCl2 also increases extracellular TNFα levels and activates the extrinsic apoptotic pathway, further promoting apoptosis.
In corneal epithelial cells, Caspase-3 expression is closely associated with inflammation and cell death [29]. For example, in dry eye disease models, Caspase-3 has been shown to exacerbate disease progression by promoting pyroptosis, a form of programmed cell death in corneal epithelial cells [30]. Similarly, elevated Caspase-3 expression has been observed in corneal alkali burns, where it contributes to inflammation and tissue damage. Notably, inhibition of Caspase-3 in these models reduced both inflammation and corneal damage, thereby promoting tissue healing [31]. Further evidence of Caspase-3’s role in corneal pathology comes from studies showing its upregulation in response to oxidative stress, again linking it to apoptosis in corneal epithelial cells [31]. Its expression is also markedly increased in viral keratitis, suggesting a central role in corneal injury caused by viral infections [32]. Beyond its apoptotic functions, Caspase-3 also modulates corneal immune responses and tissue repair, indicating its broader significance in inflammation-related corneal damage [33].
Nevertheless, the role of Caspase-3 in limbal stromal cells has not been analysed, yet. The present study confirmed elevated Caspase-3 expression in aniridia-derived limbal stromal cells in vitro, in a congenital hereditary eye disease model [30]. In addition, in contrast to diabetic keratopathy models, AN-LSCs exhibited significant Bcl-2 depletion even under normoxic conditions, suggesting that genetic defects may predispose these cells to a lower apoptotic threshold, independent of environmental triggers [34]. Importantly, this study is the first to report abnormal downregulation of pro-apoptotic genes in AN-LSCs under hypoxic stress—a phenomenon not previously described in other corneal disease models. This unique response may point to AAK-specific mechanisms of apoptosis regulation at mRNA and protein levels and offer novel insights into potential therapeutic targets in AAK.
To address impaired corneal epithelial regeneration in patients with congenital aniridia, various therapeutic strategies are under investigation, including limbal stem cell transplantation and gene therapy. These approaches aim to restore or replace damaged stem cell function, thereby promoting normal corneal epithelial renewal [35,36]. However, their clinical application remains challenging due to issues such as limited survival of transplanted cells, immune rejection, and uncertain long-term efficacy [23,35]. In terms of clinical translational value, our findings suggest that elevated Caspase 3 protein levels, in combination with dual downregulation of Bcl-2 at both mRNA and protein levels, may serve as a potential biomarker set for early AAK diagnosis. Therapeutically, Caspase inhibitors (e.g., z-VAD-fmk) and Bcl-2 analogues (e.g., ABT-199) may help restore AN-LSC viability [37,38]. Additionally, blocking TNFα signaling (e.g., with etanercept) may help interrupt the vicious cycle of hypoxia, inflammation, and apoptosis [39]. The observed upregulation of p21 and p27 further suggests that CDK inhibitors may provide a novel approach by modulating cell cycle progression in the limbal niche.
While this study yielded important findings, it also has limitations. First, the in vitro CoCl_2_ model does not fully replicate the in vivo hypoxic microenvironment. Second, the limited sample size may affect statistical power. Moreover, the specific regulatory mechanisms behind the discrepancies between mRNA and protein expression such as miRNA-mediated translational repression remain to be elucidated. In future research, we plan to expand upon the CoCl_2_ data using alternative hypoxia-induced oxidative stress models, and to explore the functional reversal of the apoptotic phenotype by repairing the Bcl-2 gene in AN-LSCs using CRISPR/Cas9 technology. Future studies should aim to increase sample sizes, integrate functional assays, and utilize in vivo models to more effectively translate our findings into clinical applications.
Conclusions
The present study confirms the activation of apoptotic pathways in corneal limbal stromal cells in AAK. Specifically, the combined effect of mitochondria-dependent apoptosis and suppression of anti-apoptotic molecules contributes to impaired corneal regeneration. Hypoxic stress further exacerbates this pathological process, in part through the synergistic influence of inflammatory mediators such as TNFα. These findings suggest that targeted modulation of the Bcl-2/Caspase axis or intervention in the hypoxia-inflammation-apoptosis cascade may represent promising therapeutic strategies to restore corneal homeostasis in patients with AAK.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Landsend ECS, Lagali N, Utheim TP. Congenital aniridia - A comprehensive review of clinical features and therapeutic approaches. Surv Ophthalmol. 2021;66(6):1031–50. doi: 10.1016/j.survophthal.2021.02.011 33675823 · doi ↗ · pubmed ↗
- 2Lim HT, Kim DH, Kim H. PAX 6 aniridia syndrome: clinics, genetics, and therapeutics. Curr Opin Ophthalmol. 2017;28(5):436–47. doi: 10.1097/ICU.0000000000000405 28598868 · doi ↗ · pubmed ↗
- 3Ramaesh K, Ramaesh T, Dutton GN, Dhillon B. Evolving concepts on the pathogenic mechanisms of aniridia related keratopathy. Int J Biochem Cell Biol. 2005;37(3):547–57. doi: 10.1016/j.biocel.2004.09.002 15618012 · doi ↗ · pubmed ↗
- 4DoràN, Ou J, Kucerova R, Parisi I, West JD, Collinson JM. PAX 6 dosage effects on corneal development, growth, and wound healing. Dev Dyn. 2008;237(5):1295–306. doi: 10.1002/dvdy.21528 18386822 PMC 2655055 · doi ↗ · pubmed ↗
- 5Ramaesh T, Ramaesh K, Martin Collinson J, Chanas SA, Dhillon B, West JD. Developmental and cellular factors underlying corneal epithelial dysgenesis in the Pax 6+/- mouse model of aniridia. Exp Eye Res. 2005;81(2):224–35. doi: 10.1016/j.exer.2005.02.002 16080917 · doi ↗ · pubmed ↗
- 6Li Z, Stachon T, Zimmermann J, Trusen S, Fries FN, Berger M, et al. Expression of PAX 6 and Keratocyte-Characteristic Markers in Human Limbal Stromal Cells of Congenital Aniridia and Healthy Subjects, In Vitro. Curr Eye Res. 2025;50(4):362–72. doi: 10.1080/02713683.2025.2449915 39791356 · doi ↗ · pubmed ↗
- 7Li S, Stachon T, Liu S, Li Z, Hsu S-L, Kundu S, et al. Increased sensitivity of primary aniridia limbal stromal cells to travoprost, leading to elevated migration and MMP-9 protein levels, in vitro. P Lo S One. 2025;20(6):e 0326967. doi: 10.1371/journal.pone.0326967 40570009 PMC 12200743 · doi ↗ · pubmed ↗
- 8Ke H, Augustine CK, Gandham VD, Jin JY, Tyler DS, Akiyama SK, et al. CYLD inhibits melanoma growth and progression through suppression of the JNK/AP-1 and β1-integrin signaling pathways. J Invest Dermatol. 2013;133(1):221–9. doi: 10.1038/jid.2012.253 22832488 PMC 3485435 · doi ↗ · pubmed ↗