Three new species of Agaricus (Agaricaceae, Agaricales) from southern China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
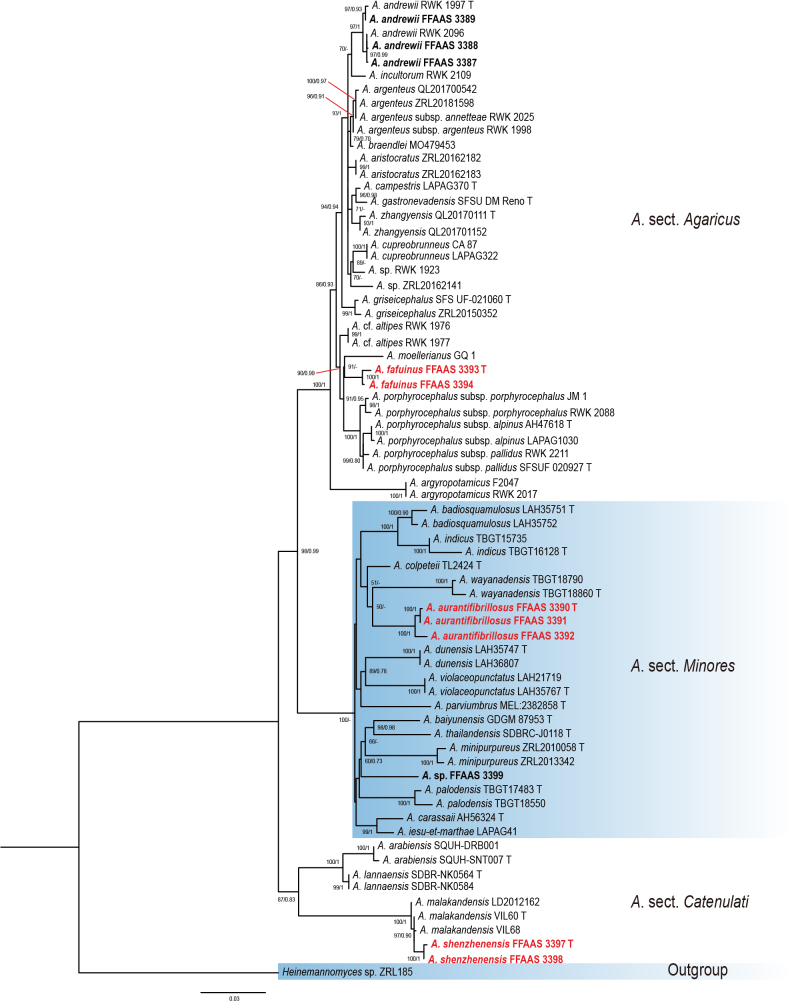 Figure 1
Figure 1 Figure 2
Figure 2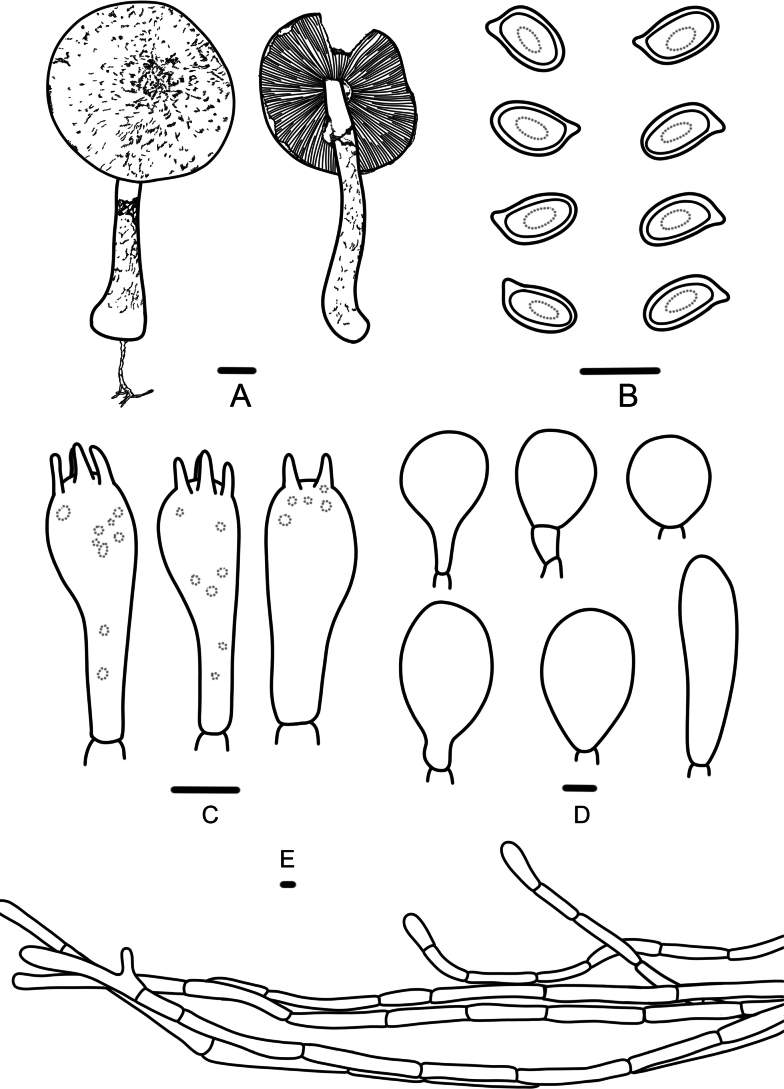 Figure 3
Figure 3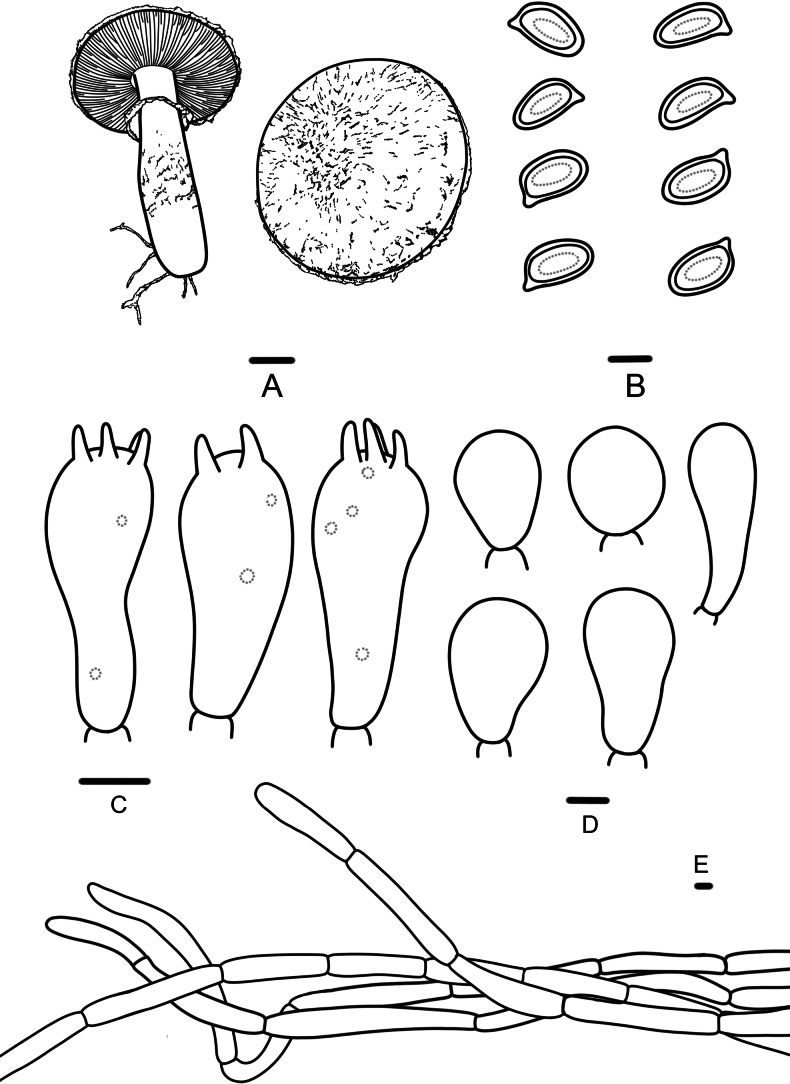 Figure 4
Figure 4 Figure 5
Figure 5| Taxon | Voucher specimen | Country | GenBank accession numbers | References | ||
|---|---|---|---|---|---|---|
| ITS | nrLSU |
| ||||
|
| RWK 2096 | The USA |
| — | — | ( |
|
| RWK 1997 T | The USA |
| — | — | ( |
|
|
|
|
|
|
|
|
|
|
|
|
| — |
|
|
|
|
|
|
| — | — |
|
|
| SQUH-DRB001 | Oman |
| — | — | ( |
|
| SQUH-SNT007 T | Oman |
|
|
| ( |
|
| ZRL20181598 | China |
| — | — | ( |
|
| QL20170054 | China |
| — | — | ( |
|
| RWK 2025 | The USA |
| — | — | ( |
|
| RWK 1998 | The USA |
| — | — | ( |
|
| RWK 2017 | The USA |
| — | — | ( |
|
| F2047 | France |
| — | — | ( |
|
| ZRL20162182 | China |
| — | — | ( |
|
| ZRL20162183 | China |
| — | — | ( |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Pakistan |
|
|
| ( | |
|
| Pakistan |
|
|
| ( | |
|
| GDGM 87953 T | China |
|
|
| ( |
|
| The USA |
| — | — | — | |
|
| LAPAG370 T | Spain |
|
|
| ( |
|
| AH56324 T | Italy |
| — | — | ( |
|
| RWK 1976 | The USA |
| — | — | ( |
|
| RWK 1977 | The USA |
| — | — | ( |
|
| TL2424 T | Australia |
| — | — | ( |
|
| CA 87 | France |
| — | — | ( |
|
| LAPAG322 | Spain |
| — | — | ( |
|
| Pakistan |
|
|
| ( | |
|
| Pakistan |
|
|
| ( | |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| SFSU DM Reno T | The USA |
| — | — | ( |
|
| SFSU F-021060 T | The USA |
| — | — | ( |
|
| ZRL20150352 | China |
| — | — | ( |
|
| LAPAG41 | Spain |
| — | — | ( |
|
| RWK 2109 | The USA |
| — | — | ( |
|
| TBGT16128 T | India |
| — | — | ( |
|
| TBGT15735 | India |
| — | — | ( |
|
| SDBR-NK0564 T | Thailand |
|
|
| ( |
|
| SDBR-NK0584 | Thailand |
|
|
| ( |
|
| ViL-60 T | Pakistan |
|
|
| ( |
|
| ViL-68 | Pakistan |
|
| — | ( |
|
| LD2012162 | Thailand |
|
|
| ( |
|
| ZRL2010058 T | China |
|
|
| ( |
|
| ZRL2013342 | China |
|
|
| ( |
|
| GQ 1 | The USA |
| — | — | ( |
|
| TBGT17483 T | India |
| — | — | ( |
|
| TBGT18550 | India |
| — | — | ( |
|
| MEL:2382858 T | Australia |
| — | — | ( |
|
| AH47618 T | Italy |
| — | — | ( |
|
| LAPAG1030 | Italy |
| — | — | ( |
|
| SFSU F-020927 T | The USA |
| — | — | ( |
|
| RWK 2211 | The USA |
| — | — | ( |
|
| JM 1 | The USA |
| — | — | ( |
|
| RWK 2088 | The USA |
| — | — | ( |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| SDBR-CJ0118 T | Thailand |
|
|
| ( |
|
| Pakistan |
|
| — | ( | |
|
| Pakistan |
| — |
| ( | |
|
| TBGT18860 T | India |
| — | — | ( |
|
| TBGT18790 | India |
| — | — | ( |
|
| QL20170111 T | China |
| — | — | ( |
|
| QL201701152 | China |
| — | — | ( |
|
|
|
|
|
|
|
|
| ZRL20162141 | China |
| — | — | ( | |
| RWK 1923 | The USA |
| — | — | ( | |
| ZRL185 | Thailand |
|
|
| ( | |
| Sample | ITS | nrLSU |
| ||||||
|---|---|---|---|---|---|---|---|---|---|
| 256 | 273 | 286 | 646 | 410 | 288 | 399 | 439 | 503 | |
|
| |||||||||
| ViL-60 T | C | T | C | C | C | G | A | C | C |
| ViL-68 | C | T | C | C | C | — | — | — | — |
| LD2012162 | C | T | C | C | C | G | A | C | C |
|
| |||||||||
| T | C | T | T | G | A | T | T | T | |
| T | C | T | T | G | A | T | T | T | |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Fungal Biology and Applications · Plant Pathogens and Fungal Diseases
Introduction
Agaricus L. is the type genus of the family Agaricaceae Chevall. (Donk 1962; Parra 2008; Kerrigan 2016; Zhao et al. 2016; Bau 2018). Species within Agaricus are primarily saprophytic fungi, typically inhabiting forest ecosystems or grasslands (Zhao et al. 2011; Chen et al. 2017). However, recent studies have expanded their known ecological niches, with documentation of lignicolous taxa such as A.subiculosus Miller, Angelini, L.A. Parra & Linda J. Chen growing on decaying wood substrates (Parra et al. 2024a). Although a minority of Agaricus species are toxic and may provoke gastrointestinal distress, the majority exhibit significant edible and medicinal value (Wu et al. 2019; Boxshall et al. 2021; Jaichaliaw et al. 2021; He et al. 2022). Currently, several species within Agaricus are utilized for food and nutraceutical applications, including A.bisporus (J.E. Lange) Imbach, A.flocculosipes R.L. Zhao, Desjardin, Guinb. & K.D. Hyde and A.subrufescens Peck (Wisitrassameewong et al. 2012; Zhao et al. 2012, 2016).
As outlined in our previous studies (Wang 2024; Wang and Bau 2024), the taxonomy of Agaricus has experienced substantial development over the past two decades, due to the incorporation of molecular characters and phylogenetic methodologies to the traditional morphological characters. To date, Agaricus comprises six subgenera, 27 sections (Zhao et al. 2016; Chen et al. 2017; Callac and Chen 2018; He et al. 2018a; Parra et al. 2018a; Ortiz-Santana et al. 2021; Hussain et al. 2022). This study focuses on three subgenera and three sections within Agaricus, viz., A.sect.Agaricus in A.subg.Agaricus, A.sect.Catenulati in A.subg.Pseudochitonia, and A.sect.Minores in A.subg.Minores.
The type species of A.sect.Agaricus is A.campestris L., which shows the following taxonomic characteristics: pileus surface unchanging or slightly yellowing, rarely rufescent on touching; context often turning pink or strongly reddening; odor usually indistinct or mushroomy, sometimes anise; annulus superous or intermediate, simple or double, membranous or fibrillose; KOH and Schäffer’s reactions negative on white areas of the surface of the pileus; cheilocystidia absent or indistinct, basidia-like, some species abundant, large, globose, piriform or ovoid, never catenulate (Zhao et al. 2016).
The type species of A.sect.Catenulati is A.arabiensis S. Hussain & Al-Sadi. Species of the section are characterized by: medium-sized basidiomata; white context unchanging when bruised or handled; a phenolic odor; a slightly yellow KOH reaction and negative Schäffer’s reactions; cheilocystidia globose to subglobose or broadly ellipsoid, the anteterminal elements subglobose or cylinderical, forming a chain-shaped structure (Hussain et al. 2022).
Agaricussect.Minores with the type species A.comtulus Fr.. This section has the following phenotypic characters: basidiomes often slender, small-tomedium sized; surface of pileus often discoloring yellow on touching; context often turning yellow on exposure; odor of anise or bitter almonds; annulus superous, simple, thin, fragile; KOH and Schäffer’s reactions positive; cheilocystidia simple, clavate, pyriform, sometimes absent, scattered or rare (Zhao et al. 2016).
The genus Agaricus has attracted increasing attention from taxonomists in recent years. In 2024, 26 novel species were described globally (Arya and Pradeep 2024; Crous et al. 2024; Bashir et al. 2024; Guzmán-Guillermo et al. 2024; Liang et al. 2024; Liu et al. 2024; Nawaz et al. 2024; Palestina-Villa et al. 2024; Parra et al. 2024a, 2024b; Ullah et al. 2024) following our previous publication on 11 January (Wang and Bau 2024). Notably, A.baiyunensis, A.cacainus and A.praeclarefibrillosus were discovered in southern China (Liang et al. 2024). Additionally, A.totalaiiensis M. Ishaq, M. Fiaz & A.N. Khalid is currently under review (https://preprints.arphahub.com/article/133080/, accessed on 4 March 2025).
In this study, we further expand the known diversity of Agaricus in China by describing three new species based on morphological characteristics and molecular phylogenetic analyses, i.e., A.aurantifibrillosus, A.fafuinus and A.shenzhenensis.
Materials and methods
Morphological studies
The macro-morphological characteristics of the species were described using notes and photographs taken from fresh basidiomata collected during field collecting. The color description of the fresh basidiomata follows the Methuen Handbook of Color (Kornerup and Wanscher 1978). Microstructures were observed using a Nikon differential interference contrast (DIC) optical microscope (Nikon Corporation, Japan). Basidiospore descriptions followed the methodologies outlined in previous studies (Wang and Bau 2024). The symbol “(a) b–c (d)” is used to describe the size of basidiospores, where the “b–c” range represents 90% of the measured values, while the “a” and “d” are extreme values. “[Xav = e × f]” indicates the average size of basidiospores. “Q” refers to the ratio of length to width of a single basidiospore from the side view, and “Qav” refers to the average value of “Q” of all specimens. The morphological descriptions of taxa followed the methods outlined in previous studies (Parra 2013; Kerrigan 2016; Zhao et al. 2016). The voucher specimens of this study are deposited in the fungarium of the Fujian Academy of Agricultural Sciences (FFAAS), China.
Phylogenetic studies
Genomic DNA was extracted from specimens using the DNA extraction kit (Fuzhou Meilisha Biotechnology Co., Ltd., Fuzhou, China). The primer pairs ITS1F/ITS4 (White et al. 1990; Gardes and Bruns 1993), LR0R/LR5 (Moncalvo et al. 2000) and EF1-983F/EF1-1567R (Rehner and Buckley 2005) were used to amplify the sequences of three DNA regions, ITS, nrLSU and tef1-α, respectively. The polymerase chain reaction (PCR) procedure was based on the protocol described by Mou and Bau (2021).
The newly generated sequences were deposited in the National Center of Biotechnology Information (NCBI) database (https://www.ncbi.nlm.nih.gov/). Sequences of phylogenetically related taxa within the same section were retrieved from NCBI and incorporated into phylogenetic analyses (Table 1). Finally, species falling within the clades of the species described in this study were selected and integrated to construct new multi-locus phylogenetic trees. Heinemannomyces sp. ZRL185 was used as an outgroup (Zhao et al. 2016; Ortiz-Santana et al. 2021). The ITS, nrLSU, and tef1-α sequences were independently aligned using the Q-INS-i algorithm via the MAFFT v.7.205 (Katoh and Standley 2013) online server (https://mafft.cbrc.jp/alignment/server/). Sequence alignments were manually adjusted in BioEdit v7.1.3.0 (Hall 1999) and subsequently concatenated for the ITS, nrLSU, and tef1-α regions using PhyloSuite v1.2.2 (Zhang et al. 2020). ModelFinder v2.2.0 (Kalyaanamoorthy et al. 2017) was used to select the best-fit model using BIC criterion.
Maximum likelihood (ML) phylogenies were inferred using IQ-TREE for 5000 ultrafast bootstraps, as well as the Shimodaira–Hasegawa–like approximate likelihood-ratio test (Guindon et al. 2010; Nguyen et al. 2015). Bayesian Inference (BI) phylogenies were inferred using MrBayes 3.2.6 (Ronquist et al. 2012) under partition model, in which the initial 25% of sampled data were discarded as burn-in. Analyses were run until convergence criteria were met (average standard deviation of split frequencies <0.01). The phylogenetic tree was visualized using FigTree v1.4.3 (http://tree.bio.ed.ac.uk/software/figtree/) and edited using Adobe Illustrator 2021 (Adobe, San Jose, CA, USA). Bootstrap support (BS) values ≥ 50% and Bayesian posterior probability (PP) values ≥ 0.70 are indicated on branches (BS/PP).
Results
Phylogenetic analyses
In this study, 30 sequences were generated including 11 ITS sequences, 9 nrLSU sequences, and 10 tef1-α sequences. The multi-locus dataset (ITS + nrLSU+ tef1-α) of Agaricus had an aligned length of 2157 bp (ITS subset: 1–725 bp; nrLSU subset:726–1595 bp; tef1-α subset: 1596–2157 bp) total characters including gaps. Alignment has 70 sequences with 2157 columns, 637 distinct patterns 407 parsimony-informative, 155 singleton sites, 1595 constant sites. The alignment was submitted to Figshare (https://figshare.com/s/778e265dd87f6d6f83b0).
In this study, Bayesian inference (BI) and maximum likelihood (ML) phylogenetic trees were reconstructed using a concatenated dataset comprising ITS, nrLSU, and tef1-α sequences. BI and ML analysis resulted in a very similar topology, so the ML tree is provided in this study (Fig. 1). For the construction of the maximum likelihood (ML) phylogenetic tree, the best-fit substitution model (TIM2+F+I+G4, without partitioning) was selected using BIC criterion. Similarly, for the Bayesian inference (BI) phylogeny, a partitioned model (edge-linked) was optimized using BIC, with the following partitions: HKY+F+G4 for ITS, GTR+F+I+G4 for nrLSU, and K2P+G4 for tef1-α.
Multi-locus phylogenetic tree of Agaricus obtained from the maximum likelihood analysis (ML) based on ITS, nrLSU, and tef1-α sequence data. “T” refers to the type specimen. Bold refers to the sequences produced from this study. Red font refers to the new species.
The phylogenetic tree revealed three major clades, corresponding to three sections: A.sect.Agaricus, A.sect.Catenulati and A.sect.Minores. A.fafuinus and A.moellerianus Bon formed a sister clade (BS/PP = 91/-) in A.sect.Agaricus. Within A.sect.Catenulati, A.shenzhenensis and A.malakandensis formed a sister clade with a strong support value (BS/PP = 97/0.90). Within A.sect.Minores, A.aurantifibrillosus is phylogenetically close to A.colpeteii T. Lebel and A.wayanadensis. Despite low support values, A.aurantifibrillosus forms a distinct clade.
Taxonomy
Agaricus
aurantifibrillosus
Taxon classificationFungiAgaricalesAgaricaceae
P.H. Liu & S.E. Wang sp. nov.
E376667B-C689-5DE7-95D9-EF79C43A28CC
858060
Etymology.
aurantifibrillosus (Latin), referring to the pileus covered with arranged orange (5A8) or brownish yellow (5C8) fibrillose squamules.
The photographs of fresh basidiomata of Agaricus species in this study. A.A.aurantifibrillosus (FFAAS 3390); B.A.aurantifibrillosus (FFAAS 3391); C.A.fafuinus (FFAAS 3393); D.A.fafuinus (FFAAS 3394); E.A.shenzhenensis (FFAAS 3397); F.A.shenzhenensis (FFAAS 3398). Scale bars: 1 cm.
Holotypus.
China • Fujian Province, Fuzhou City, Fuzhou National Forest Park, 2 October 2024, 26°10'41"N, 119°16'19"E, alt. 280 m, Shi-En Wang, E2410232 (FFAAS 3390).
Agaricusaurantifibrillosus (FFAAS 3390, FFAAS 3391); A. Basidiomata; B. Basidiospores; C. Basidia; D. Cheilocystidia; E. Pileipellis. Scale bar: 1 cm (A); 5 μm (B–E).
Diagnosis.
This species is characterized by its pileus covered with orange (5A8) or brownish yellow (5C8) scattered fibrillose squamules, elongate basidiospores (Qav = 1.70), and abundant cheilocystidia.
Description.
Pileus 3.6–6.2 cm in diameter, 0.2–0.3 cm thick at the center, truncate conical to plane, surface dry, white (5A1) or orange white (5A2), covered with scattered arranged orange (5A8), brownish yellow (5C8) fibrillose squamules, denser at disc, showing brownish orange (6C8) or brown (6E8), margin appendiculate by annulus remnants. Context of the pileus white (6A1), with no special odor. Lamellae 0.2–0.4 cm broad, pastel red (9A4) then reddish brown (8E8) later brownish black (8F8), free, crowded, intercalated with numerous lamellulae. Stipe 5.6–8.6 × 0.4–1.3 cm, hollow, clavate, with white (6A1) rhizomorphs, provided with an annulus in its upper third, above the annulus white (6A1), below the annulus with white (6A1), orange (5A8) floccose squamules, becoming dark yellow (4C8) on touching or bruising. Annulus superior, white (6A1), simple, membranous, easy falling out.
Basidiospores (4.6)4.7–5.7(5.9) × (2.7)2.8–3.3(3.6) μm, [Xav = 5.2 × 3.1 μm], Q = 1.50–1.93, Qav = 1.70, ellipsoid to elongate-ellipsoid, smooth, thick-walled, brown, guttulate. Basidia 14–18 × 5–7 μm, clavate, 4(2)-spored, sterigmata 2–4 µm long. Cheilocystidia abundant, nearly globose, oblong, sphaeropedunculate, or broadly clavate, 12–34 × 10–20 μm, with pale yellowish intracellular pigment. Pleurocystidia absent. Pileipellis a cutis of cylindrical, slightly constricted at the septa, pale yellowish hyphae, 4–9 μm in diameter.
Habitat and distribution.
Gregarious or scattered in broad-leaved and bamboo forests during autumn. Currently, it has only been documented in Fujian Province, China.
Additional specimens measured.
China • Fujian Province, Fuzhou City, Fujian Agriculture and Forestry University, 5 October 2024, Shi-En Wang, E2410524 (FFAAS 3391) and E2410525 (FFAAS 3392).
Notes.
Agaricusaurantifibrillosus belongs to A. (subg. Minores) sect. Minores. Agaricusaurantipileatus T. Bau & S.E. Wang in A.sect.Arvenses shares morphological similarities with A.aurantifibrillosus. However, A.aurantipileatus can be distinguished by its double annulus, smaller spore Q value (Q = 1.17–1.36), and sometimes catenulate cheilocystidia (Wang and Bau 2024).
In the multi-locus phylogenetic tree (Fig. 1), A.aurantifibrillosus clusters with A.colpeteii T. Lebel and A.wayanadensis, albeit with low statistical support, and this may be due to the lack of sequences in the related taxa. Agaricuscolpeteii is a gasteroid Agaricus species with basidia not observed (Lebel 2013). Agaricuswayanadensis differs in having a pileus surface covered with brown squamules and a smaller spore Qav value (Qav = 1.58) (Arya and Pradeep 2024).
Agaricus
fafuinus
Taxon classificationFungiAgaricalesAgaricaceae
P.H. Liu & S.E. Wang sp. nov.
5F9C3FAF-FE6A-578F-A022-5F090B7BFB5A
858061
Etymology.
Derived from the acronym FAFU (Fujian Agriculture and Forestry University), where the type specimens of this species were collected.
Agaricusfafuinus (FFAAS 3393, FFAAS 3394); A. Basidiomata; B. Basidiospores; C. Basidia; D. Cheilocystidia; E. Pileipellis. Scale bar: 1 cm (A); 5 μm (B–E).
Holotypus.
China • Fujian Province, Fuzhou City, Fujian Agriculture and Forestry University, 4 March 2024, 26°08'N, 119°24'E, alt. 30 m, Si-Ang Chen, CSA299 (FFAAS 3393).
Diagnosis.
This species is characterized by its pileus and stipe covered with brown (7D5) or fox red (8D7) fibrils or fibrillose squamules, elongate basidiospores (Qav = 1.63), and abundant cheilocystidia.
Description.
Pileus 3.4–5.5 cm in diameter, 0.2–0.4 cm thick at the center, truncate conical to plane, surface dry, white (5A1), brownish gray (7C2), covered with brown (7D5) or fox red (8D7) fibrils or with fibrillose squamules, concentrically arranged, denser and reddish brown (8E8) at disc, margin appendiculate by annulus remnants. Context of the pileus, white (6A1), with no special odor. Lamellae 0.2–0.3 cm broad, first pastel red (9A4), then reddish brown (8E8), later brownish black (8F8), free, crowded, intercalated with numerous lamellulae. Stipe 2.2–4.5 × 0.6–1.5 cm, nearly cylindrical, hollow, with white (6A1) rhizomorphs, provided with an annulus in its upper half, above the annulus white (6A1), below the annulus covered towards the base with concolorous squamules with the pileus surface. Annulus superior, white (6A1) to reddish brown (8E8), simple, membranous, persistent.
Basidiospores (5.7)6.0–7.9(8.2) × (3.5)3.7–5.0(5.1) μm, [Xav = 6.8 × 4.2 μm], Q = 1.46–2.03, Qav = 1.63, ellipsoid to cylindrical, smooth, thick-walled, brown, guttulate. Basidia 17–25 × 6–9 μm, clavate, 4(2)-spored, sterigmata 1–3 µm long. Cheilocystidia abundant, nearly globose, broadly clavate, or pyriform, 10–23 × 7–13 μm, hyaline. Pleurocystidia absent. Pileipellis a cutis of cylindrical, slightly constricted at septa, light brown hyphae, 6–12 μm wide.
Habitat and distribution.
Gregarious in bamboo forests or grass during spring. Currently, it has only been known from Fujian Province, China.
Additional specimens measured.
China • Fujian Province, Fuzhou City, Fujian Agriculture and Forestry University, 23 March 2024, Si-Ang Chen, CSA314 (FFAAS 3394).
Notes.
Agaricusfafuinus belongs to A. (subg. Agaricus) sect. Agaricus. Agaricusfafuinus exhibits variable pileus surface: specimen CSA299 possesses fibrils, while CSA314 displays fibrillose squamules. However, molecular data confirm their conspecificity, suggesting that these morphological differences may result from humidity variations. Specimen CSA299 was collected post-rainfall and likely it was exposed to precipitation, whereas specimen CSA314 was obtained under dry conditions. Initially, we thought we had mixed up the specimen numbers, but after thorough verification, this turned out to be accurate.
In the multi-locus phylogenetic tree (Fig. 1), A.fafuinus and A.moellerianus Bon form a sister clade with good support. However, A.moellerianus is distinguished by its subglabrous white pileus, smooth stipe, rounder basidiospores (Qav = 1.22), and broadly clavate cheilocystidia resembling basidioles (Kerrigan 2016).
Agaricus
shenzhenensis
Taxon classificationFungiAgaricalesAgaricaceae
P.H. Liu & S.E. Wang sp. nov.
9957CD28-9E14-588F-A514-BE678DB58C09
858063
Etymology.
shenzhenensis (Latin), meaning from shenzhen city where the holotype specimen was collected.
Agaricusshenzhenensis (FFAAS 3397, FFAAS 3398); A. Basidiomata; B. Basidiospores; C. Basidia; D. Cheilocystidia; E. Pileipellis. Scale bar: 1 cm (A); 5 μm (B–E).
Holotypus.
China • Guangdong Province, Shenzhen City, Lianhuashan Park, 29 April 2024, 22°33'23"N, 114°3'13"E, alt. 100 m, Cheng-Cheng, 20240430 (FFAAS 3397).
Diagnosis.
Distinguished by the pileus adorned with brownish gray (6E2) fibrillose squamules, context becoming yellow (3B8) on cutting, stipe base tapering and covered with brownish gray (6E2) fibrillose squamules, predominantly 2-spored basidia.
Description.
Pileus 2.5–6.5 cm in diameter, 0.5–0.8 cm thick at the center, hemispherical, truncate conical to plane, applanate with a slightly depressed center when mature, surface dry, white (2A1), grayish white (2B1), entirely covered with appressed, concentrically arranged, triangular, brownish gray (6E2) fibrillose squamules, scattered towards the margin in age, denser taupe (4F1), black (2F1) at the disc, margin entire, appendiculate by annulus remnants. Context of the pileus white (6A1), becoming yellow (3B8) on cutting, odor unknown. Lamellae 0.5–0.7 cm broad, first pale red (9A3) then reddish brown (8E8), later brownish black (8F8), free, crowded, intercalated with numerous lamellulae. Stipe 4.5–11.5 × 0.5–2.0 cm, cylindrical, tapering downwards, hollow, with short white (6A1) rhizomorphs, provided with an annulus in its upper part, above the annulus white (6A1), below the annulus the upper half white and the lower half covered with dense adpressed brownish grey (6E2) squamules concolorous with the pileus surface. Annulus superior, white (6A1) to brownish black (8F8), simple, membranous, persistent.
Basidiospores (5.8)6.1–7.3(7.5) × (3.8)4.0–4.3(4.4) μm, [Xav = 6.8 × 4.2 μm], Q = 1.41–1.87, Qav = 1.61, ellipsoid to elongate-ellipsoid, smooth, brown, thick-walled, guttulate. Basidia 25–33 × 8–10 μm, clavate, 2(4)-spored, sterigmata 2–3 µm long. Cheilocystidia abundant, broadly clavate, oblong or pyriform, 22–50 × 10–22 μm. Pleurocystidia absent. Pileipellis a cutis of cylindrical, slightly constricted at the septa, light brown hyphae, 4–7 μm wide.
Habitat and distribution.
Gregarious or clustered in grass or broad-leaved forests during spring. Currently, it is only known from Guangdong Province, China.
Additional specimens measured.
China • Guangdong Province, Shenzhen City, Lianhuashan Park, 29 April 2024, Cheng-Cheng, 20240430-1 (FFAAS 3398).
Notes.
Agaricusshenzhenensis belongs to A. (subg. Pseudochitonia) sect. Catenulati, which currently comprises three species: A.arabiensis S. Hussain & Al-Sadi, A.lannaensis N. Suwannarach, J. Kumla & S. Lumyong and A.malakandensis.
Agaricusshenzhenensis differs from all three in both morphological and molecular characters. Agaricusarabiensis exhibits a reddish-brown to dark reddish-brown pileus, context unchanged on handling, smooth stipe, and globose to subglobose or broadly clavate, regularly catenulate cheilocystidia (Hussain et al. 2022). Agaricuslannaensis differs in having brown pileus, reddish brown context when cut, fibrillose stipe white below the annulus to the base, and smaller basidia (19–26 × 5.5–8.5 µm) (Jaichaliaw et al. 2021). Agaricusmalakandensis possesses a dark brown to reddish brown pileus, smooth or fibrillose to slightly squamulose stipe, and multiseptate cheilocystidia with clavate to pyriform terminal element (Nawaz et al. 2024). Notably, catenulate cheilocystidia were not observed in this species, even though this feature is the primary morphological diagnostic characteristic for taxa within this section.
The genetic distinctions between A.shenzhenensis and A.malakandensis remain unequivocal, with four nucleotide differences in the ITS region, one in nrLSU, and four in tef1-α (Table 2). To ensure the accuracy of the results, we rechecked the quality of the sequences and confirmed their compliance with the required standards.
Notably, this species represents the first record of A.sect.Catenulati in China, expanding the known biogeographic range of this section.
Discussion
Of the 26 recently described Agaricus species, five, A.calolepidotus, A.xalapensis, A.karakensis, A.palodensis and A.wayanadensis, were delineated solely through ITS sequence data (Arya and Pradeep 2024; Guzmán-Guillermo et al. 2024; Ullah et al. 2024), without multi-locus phylogenetic analyses. The same applies to A.totalaiiensis, which is currently under review. Notably, the original description of A.karakensis lacks explicit GenBank accession numbers for its molecular data (Ullah et al. 2024). However, phylogenetic analyses of Agaricus based exclusively on ITS sequences lack methodological rigor (Cao et al. 2020). To ensure taxonomic reliability, the description of novel Agaricus species should integrate analyses of multi-locus (e.g., ITS + nrLSU + tef1-α). Furthermore, resolving higher-level phylogenetic relationships using single-copy orthologous genes derived from genomic data (He et al. 2024; Wang et al. 2024) is emerging as a trend.
Convergent evolution is a notable phenomenon within Agaricus, exemplified by striking macro-morphological similarities between A.aurantifibrillosus and A.aurantipileatus. Remarkably, this convergence extends beyond intra-generic boundaries, as seen in A.sinoagrocyboides T. Bau & S.E. Wang, a species our team previously discovered in China, which exhibits macroscopic traits strongly resembling those of Agrocybe species (Wang and Bau 2024). Such morphological variability becomes more complex in certain identifications due to the ecological plasticity of Agaricus. For instance, the fact that A.subiculosus is growing on decaying wood challenges traditional habitat assumptions for Agaricus (Parra et al. 2024a). These observations underscore the necessity of collecting morphologically ambiguous specimens during field surveys, as overlooked specimens may represent cryptic lineages or novel ecological adaptations within Agaricus.
During our field surveys targeting Agaricus specimens, we inadvertently collected specimens of the genera Leucoagaricus, Leucocoprinus, Micropsalliota and Xanthagaricus. These specimens will undergo comprehensive morphological and molecular analyses to explore potential novel discoveries.
Supplementary Material
XML Treatment for Agaricus aurantifibrillosus
XML Treatment for Agaricus fafuinus
XML Treatment for Agaricus shenzhenensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arya CP Pradeep CK (2024) Agaricussection Minores: New and noteworthy species from India.Phytotaxa 634: 255–273. 10.11646/phytotaxa.634.3.5 · doi ↗
- 2Bashir H Asif M Ghafoor A Niazi AR Khalid AN Parveen G Harun N Afshan N-S Bibi A Callac P (2024) Multigene phylogeny and morphological descriptions of five species of Agaricussect.Minores from subtropical climate zones of Pakistan. PLOS ONE 19: e 0302222. 10.1371/journal.pone.0302222 PMC 1123906038990811 · doi ↗ · pubmed ↗
- 3Bau T (2018) Mushroom Taxonomy.Science Press, Beijing, China, 303 pp.
- 4Boxshall AG Birch JL Lebel T Symonds MRE Callahan DL (2021) A field-based investigation of simple phenol variation in Australian Agaricusxanthodermus. Mycologia 113: 1123–1135. 10.1080/00275514.2021.193685134494944 · doi ↗ · pubmed ↗
- 5Callac P Chen J (2018) Tropical species of Agaricus. In: Sánchez JE Mata G Royse DJ (Eds) Tropical Mushrooms.El Colegio de la Frontera Sur, Chiapas, Mexico, 25–38.
- 6Cao B He MQ Ling ZL Zhang MZ Wei SL Zhao RL (2020) A revision of Agaricussection Arvenses with nine new species from China.Mycologia 113: 191–211. 10.1080/00275514.2020.183024733326360 · doi ↗ · pubmed ↗
- 7Chen J Callac P Parra LA Karunarathna SC He MQ Moinard M De Kesel A RaspéO Wisitrassameewong K Hyde KD Zhao RL (2017) Study in Agaricussubgenus Minores and allied clades reveals a new American subgenus and contrasting phylogenetic patterns in Europe and Greater Mekong Subregion.Persoonia 38: 170–196. 10.3767/003158517 X 69552129151632 PMC 5645183 · doi ↗ · pubmed ↗
- 8Crous PW Wingfield MJ JurjevićŽBalashov S Osieck ER Marin-Felix Y Luangsa-ard JJ Mejía LC Cappelli A Parra LA Lucchini G Chen J Moreno G Faraoni M Zhao RL WeholtØBorovička J Jansen GM Shivas RG Tan YP Akulov A Alfenas AC Alfenas RF Altés A Avchar R Barreto RW Catcheside DEA Chi TY Esteve-Raventós F Fryar SC Hanh LTM Larsbrink J Oberlies NH Olsson L Pancorbo F Raja HA Thanh VN Thuy NT Ajithkumar K Akram W Alvarado P Angeletti B Arumugam E Khalilabad AA Bandini D Baroni TJ Barreto GG Boertmann D Bose T Castañeda Ruiz RF Couceiro A Cykowska-Marzencka B Dai YC Darmostuk Vda Silva SBG Dearnaley JD Wde Aze · doi ↗ · pubmed ↗