Four new Planothidium species (Achnanthidiaceae, Bacillariophyceae) from the Karst region of Guizhou in China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
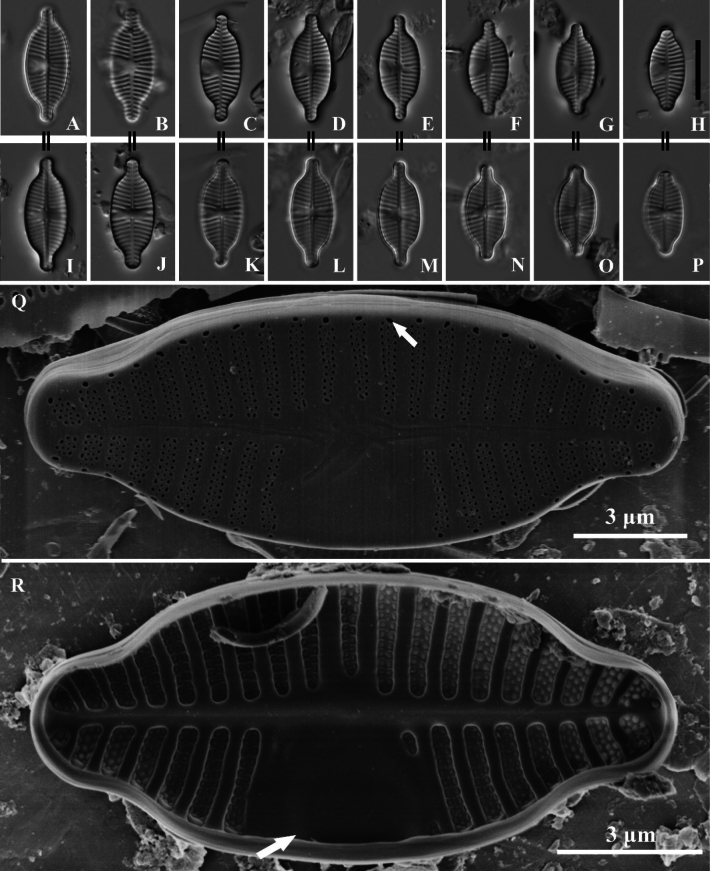 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3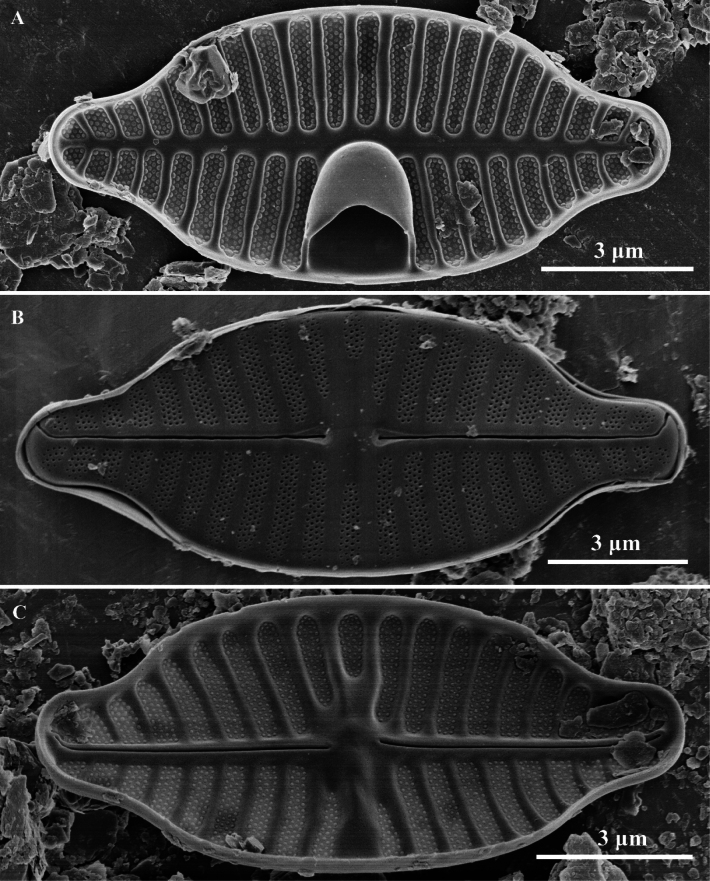 Figure 4
Figure 4 Figure 5
Figure 5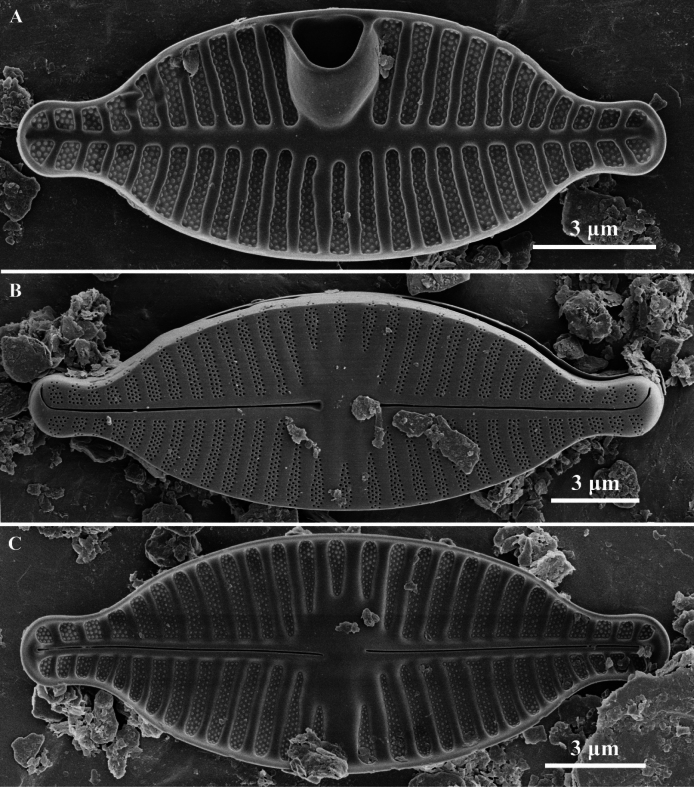 Figure 6
Figure 6 Figure 7
Figure 7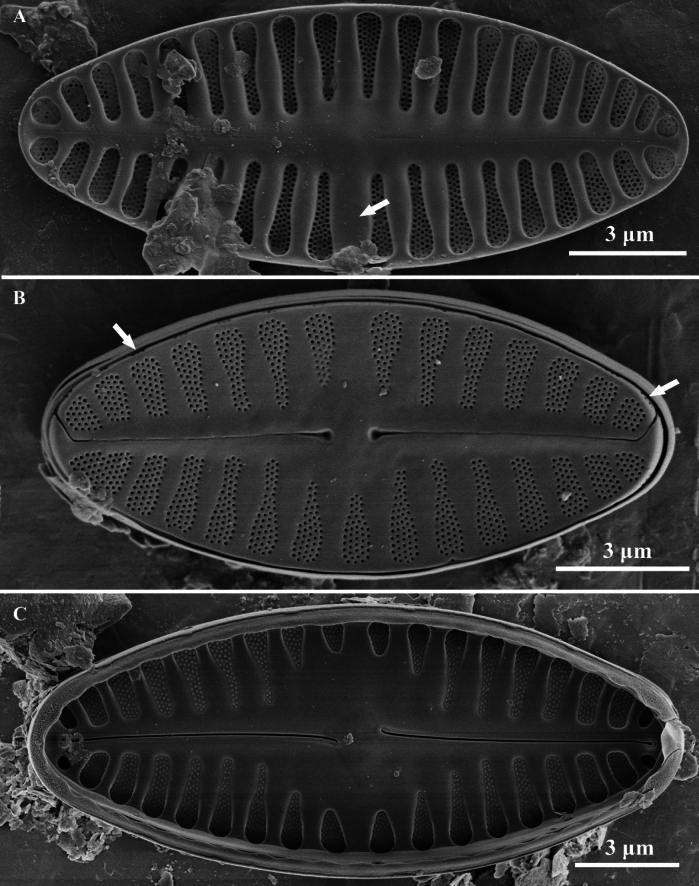 Figure 8
Figure 8| No. of samples | Location | Coordinates | Habitat | Altitude (m) | Water Temp (°C) | pH | Cond. (μS/cm) | Collector | Collection Date |
|---|---|---|---|---|---|---|---|---|---|
| GZ201510041P | Xiaoqikong Scenic Area |
| Attached to rocks in the pond | 780 | 18.0 | 7.5 | 226 | Kociolek & Wang Q.X. | 10.2.2015 |
| GZ201510045 | Xiaoqikong Scenic Area |
| Attached to the stones in the rushing water | 780 | 18.0 | 7.5 | 226 | Wang Q.X. & Kociolek J.P. | 10.2.2015 |
| GZ201510051 | Xiaoqikong Scenic Area |
| Attached to floating things in the pond | 780 | 18.0 | 7.5 | 226 | Wang Q.X. & Kociolek J.P. | 10.2.2015 |
| GZ201510066 | Xiaoqikong Scenic Area |
| Attached to rocks beneath the waterfall | 629 | 19.5 | 8.0 | 215 | Wang Q.X. & Kociolek J.P. | 10.2.2015 |
| GZ201510099 | Maolan Nature Reserve |
| Floating in the slow subsurface stream | 650 | 18.0 | 7.9 | 203 | Wang Q.X. & Kociolek J.P. | 10.4.2015 |
| GZ201510100 | Maolan Nature Reserve |
| Floating in the slow subsurface stream | 650 | 18.0 | 7.9 | 203 | Wang Q.X. & Kociolek J.P. | 10.4.2015 |
| GZ201510108 | Maolan Nature Reserve |
| Attached to rocks in a rapid river. | 811 | 18.0 | 7.8 | 205 | Wang Q.X. & Kociolek J.P. | 10.4.2015 |
| Reference | This study |
| This study |
| |
| Valve outline | Elliptical | Elliptical to linear-elliptical | Elliptical | Elliptical | Elliptical, asymmetrical |
| Apices | Rostrate | Subrostrate to capitate | Rostrate | Short to long rostrate | Short to long rostrate |
| Length | 11.5–19.0 µm | 8–18 µm | 12–18 µm | 6.5–15.0 µm | 12–14 µm |
| Width | 5.5–7.5 µm | 4.0–6.5 µm | 5–7 µm | 4.0–6.5 µm | 6.7–8.2 µm |
| Central area ( | Rectangular to nearly circular | Nd | Rectangular to nearly circular | Rectangular to slightly round | Irregular, rectangular to slightly rounded |
| Axial area ( | Narrow linear | linear | Narrow linear | Narrow, linear, widening towards the central area | Narrow linear |
| Striae ( | 14–18/10 µm | Nd | 14–17/10 µm | 12–16/10 µm | 12–14 µm |
| Areolae composition of striae ( | 3–4 rows areolae | Nd | 4–5 rows areolae | 3–4 rows areolae | 3–4 rows areolae |
| Axial area ( | Narrow, linear to lanceolate | linear | Narrow linear | Narrow, straight, linear | Narrow, linear, expanded in the central area opposite to the cavum |
| Additional structure ( | Sinus | Sinus | Cavum | Cavum | Cavum |
| Striae ( | 14–17/10 µm | Nd | 15–16/10 µm | 12–14/10 µm | 12–14 µm |
| Areolae composition of striae ( | 3–4 rows areolae | Nd | 4–5 rows areolae | 3–4 rows areolae | 1–3 rows areolae |
| Reference | This study |
|
| |
| Valve outline | Elliptic-lanceolate | Lanceolate, elliptic-lanceolate | Lanceolate to broadly elliptic–lanceolate | Elliptical |
| Apices | Narrowly rostrate to subcapitate | Rostrate | Strongly rostrate | Subrostrate |
| Length | 17–21 µm | 20–28 µm | 11.5–14.5 µm | 6.4–13.5 µm |
| Width | 6.5–7.0 µm | 7.0–8.0 µm | 5.0–6.0 µm | 3.8–6.4 µm |
| Central area ( | Subcircular to rectangular | Rectangular | Wide rectangular | Irregular |
| Axial area ( | Narrow linear | Narrow linear | Wide rectangular | Narrow, linear |
| Striae near to central area ( | 3–4 shortened striae on each side | 2–3 shortened striae on each side | 1–3 shortened striae on each side | Usually 1 shortened striae on each side |
| Raphe | Straight | Straight | Straight | Straight |
| Striae ( | 13–14/10 µm | Nd | Nd | 14–17/10 µm |
| Areolae composition of striae ( | 3–4 rows areolae | Nd | 4 rows areolae | 2–3 rows areolae |
| Axial area ( | Narrow linear | Narrow, linear, expanded in the central area | Linear–lanceolate,widening in the central area* | Narrow, straight, linear |
| Additional structure ( | Cavum | Cavum | Cavum | Cavum |
| Striae ( | 13–14/10 µm | 15–17/10 µm | 12–13/10 µm | 13–17/10 µm |
| Areolae composition of striae ( | 3–4 rows areolae | Nd | 3–4 rows areolae | 2–3 rows areolae |
| Reference | This study |
|
|
|
| Valve outline | Elliptical | Linear-lanceolate or elliptic-lanceolate | lanceolate to elliptic-lanceolate in smaller valves with cuneately rounded | Elliptical-lanceolate |
| Apices | Rounded ends not extended | Very weakly protracted, slightly rostrate to broadly rounded in smaller valves | Only very weakly protracted apices | Moderately produced, obtusely rounded |
| Length | 9.5–18.0 µm | 7–30 µm | 7–13 µm | 17–26 µm |
| Width | 5.5–7.5 µm | 4.5–7.0 µm | 4.0–4.5 µm | 6.5–9.5 µm |
| Central area ( | Subcircular | Asymmetrical | Rounded to rectangular | Circular |
| Axial area ( | Narrow linear | Narrow, linear | Very narrow, linear | Narrow, linear |
| Raphe | Straight to slightly curved | Straight to weakly curved | Straight | Straight |
| Striae ( | 10–12/10 µm | 8–10/10 µm | 16–17/10 µm | 12–15/10 µm |
| Areolae composition of striae ( | 5–6 rows areolae | 2–4 rows areolae | 2–3 rows areolae | 4 rows areolae |
| Axial area ( | Narrow linear | Narrow linear | Linear to narrowly lanceolate | Narrow, linear |
| Additional structure ( | Absent | Absent | Absent | Absent |
| Striae ( | 10–12/10 µm | 7–11/10 µm | 15–16/10 µm | 14–16/10 µm |
| Areolae composition of striae ( | 4–5 rows areolae | 3–4 rows areolae | 2 rows areolae | 3–4 rows areolae |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDiatoms and Algae Research · Paleontology and Stratigraphy of Fossils · Marine and coastal ecosystems
Introduction
Monoraphid diatoms are particularly intriguing due to their structural asymmetry, characterized by the presence of a raphe on one valve and its absence on the other (Kulikovskiy et al. 2016). The genus Planothidium Round & Bukhtiyarova was established to accommodate monoraphid diatoms previously classified within the genus Achnanthidium Kützing (Round and Bukhtiyarova 1996). Currently, approximately 129 accepted taxa of Planothidium are recorded in AlgaeBase (Guiry and Guiry 2024). The genus is characterized by the presence of multiseriate striae on both the raphe valve (RV) and rapheless valve (SV), as well as asymmetrical structures in the central area of the rapheless valve in many species (Morales 2006). Since its establishment, Planothidium has undergone significant taxonomic refinement, with studies describing new species and exploring its morphological and ecological diversity. The application of scanning electron microscopy and molecular techniques has further enhanced the differentiation of Planothidium species and deepened our understanding of their morphological features (Novis et al. 2012; Álvarez-Blanco and Blanco 2013; Van De Vijver et al. 2013; Jahn et al. 2017; Van De Vijver and Bosak 2019; Wetzel et al. 2019).
Morphological studies of Planothidium have focused on key characteristics, such as the structure of the rapheless valve’s central area, valve outline, and striae patterns. Morales (2006) proposed a classification framework for Planothidium species based on the central area morphology of the rapheless valve, dividing them into four groups: (1) species with continuous, uninterrupted striae in the central area, such as P.daui (Foged) Lange-Bertalot; (2) species with a variable, clear space in the central area that does not show any indentation or convexity, such as P.minutissimum (Krasske) Lange-Bertalot; (3) species with a depressed central area without striae, such as P.lanceolatum (Brébisson ex Kützing) Lange-Bertalot; (4) species with a depressed region on one side of the central area, which is capped to form a cave-like structure called a “cavum”, such as P.biporomum (Hohn & Hellerman) Lange-Bertalot (Morales 2006). In the study by Stancheva et al. (2020), Planothidium species were categorized into three groups: those with a sinus (single horse-shoe shaped mark), such as P.lanceolatum; those with a cavum (double horse-shoe shaped mark), such as P.frequentissimum (Lange-Bertalot) Lange-Bertalot; and those lacking both a sinus and a cavum, such as P.minutissimum (Krasske) Morales and P.daui (Foged) Lange-Bertalot. Moreover, beyond classic morphological measurements (such as width, length, length-to-width ratio, and number of striae per 10 µm), several other features observed in the SEM have been suggested as useful for species identification, including (1) the pattern of surface smoothness on the rapheless valve; (2) the number of areolae per stria on the rapheless valve; and (3) whether the striae on the rapheless valve interrupt at the valve margin, thus influencing the number of areolae at the valve edge (Wetzel et al. 2019). In addition to morphological studies, molecular tools have provided new insights into the taxonomy and phylogeny of Planothidium. Molecular research has confirmed the presence of two distinct clades within the genus, one with species possessing a sinus or cavum in the rapheless valve and the other lacking these structures (Jahn et al. 2017).
Currently, research on the genus Planothidium in China remains limited. In marine environments, one new species has been reported and described (Li et al. 2024). However, in freshwater ecosystems, only a few species have been recorded as new records, and no new species have been reported so far (Liu et al. 2015, 2016; Wang et al. 2019). Karst ecosystems, characterized by carbonate-rich and nutrient-poor waters, remain underexplored. These unique habitats may drive morphological differentiation and adaptive evolution, offering immense potential for diatom taxonomy studies. Recently, several new diatom species have been described from karst regions, such as Sellaphoragologonica Lai, Ector & Wetzel, Germainiellalegionensis Blanco, Borrego-Ramos & Olenici, Achnanthidiummediolanceolatum Yu, You & Kociolek, Amphorabaotuensis Li, Nagumo & Xu, and Fallaciacinariana Sömek, Hamilton, Solak, Beauger & Sevindik et al. The discovery of these species highlights the underexplored diatom biodiversity in karst environments (Borrego-Ramos et al. 2018; Lai et al. 2018; You et al. 2019b; Li et al. 2022; Sömek et al. 2025).
In recent years, we carried extensive surveys of diatoms species diversity of China (Kociolek et al. 2016b, 2016a, 2019; Lowe et al. 2017; You et al. 2017, 2019a; Yu et al. 2017; Zhou et al. 2024). Based on the collections developed for these studies, four new Planothidium species within karst landform of Guizhou have been discovered. This study provides detailed descriptions, as well as light and scanning electron microscopy of these new species: Planothidiumpseudoreichardtii sp. nov., Planothidiumliboensis sp. nov., Planothidiumangustirostratum sp. nov. and Planothidiummaolanensis sp. nov., comparing them with similar species within the genus. This work contributes valuable reference material for the taxonomy and morphology of freshwater Planothidium, while emphasizing the importance of karst habitats as biodiversity hotspots for diatoms.
Materials and methods
Diatom samples were collected from Xiaoqikong Scenic Area and Maolan Nature Reserve (Libo County, Guizhou Province), which is characterized by a subtropical climate and is a typical karst landscape. The sampling sites were distributed in various water bodies, including streams, ponds and underground rivers. The water parameters (pH, Temperature and Conductivity) were measured using a YSI Pro Plus multiparameter meter (YSI, Ohio, USA). Samples were collected using tweezers and a knife, then placed into sealed plastic bottles with the addition of formalin for preservation (final concentration of 4%). Information about the sampling sites is listed in Table 1.
In the laboratory, samples were processed to remove organic matter and excess impurities using a microwave-assisted reaction system (Model MARS, CEM Corporation, Charlotte, USA) with concentrated nitric acid. The specific steps for sample processing and methods of preservation are as described in You et al. (2021). Clean samples after processing were prepared for both light microscopy (LM) and scanning electron microscopy (SEM). For LM, samples were encapsulated in Naphrax and observed using an Olympus BX-53 microscope with DIC optics and a 1.4 numerical aperture, 100× oil immersion objective. For SEM, the same samples were dried on metal stubs and imaged using a SU8010 SEM at 2 kV with a working distance (WD) of less than 6 mm (Hitachi High-Technologies Corp., Tokyo, Japan). To determine the relative abundance of the new species and record co-occurring diatom species, 400 intact and identifiable diatom valves from each sample were identified under light microscopy (1000×). The images were compiled using Adobe Photoshop 2023. The morphological terminology follows (Morales 2006; Van De Vijver et al. 2018; Wetzel et al. 2019; Morais et al. 2020). The holotype images of each species correspond to specimens that have been circled on the permanent slides. All samples and permanent slides are stored at Lab of Algae and Environment, College of Life Sciences, Shanghai Normal University.
Results
Phylum Bacillariophyta
Class Bacillariophyceae Haeckel, 1878
Order Achnanthales Silva, 1962
Family Achnanthidiaceae Mann, 1990
Genus Planothidium Round & Bukhtiyarova, 1996
Planothidium
pseudoreichardtii
Taxon classificationPlantaeAchnanthalesAchnanthidiaceae
Q-M. You, P. Yu & J.P. Kociolek sp. nov.
09DF36F8-ECAA-562C-8006-2B02ACA36692
Holotype.
SHTU! Slide GZ201510045, holotype illustrated in Fig. 1C, J. Diatom samples are housed in the Lab of Algae and Environment, College of Life Sciences, Shanghai Normal University, China.
Planothidiumpseudoreichardtii sp. nov. LM; A–H. Rapheless valves; I–P. Raphe valves. “=” indicates the different valves of the same frustule. Q.SEM external view of an entire rapheless valve. The white arrow points to a larger areola. R.SEM internal view of an entire rapheless valve, showing a distinct sinus in the central area (white arrow). Scale bar as shown. Scale bars: 10 µm (A–P).
Isotype.
COLO! Material 11111, Slides are housed in the Kociolek Collection, University of Colorado, Museum of Natural History Diatom Herbarium, Boulder, U.S.A.
Type locality.
China. Xiaoqikong Scenic Area, Libo County, Guizhou Province, 25°15'36"N, 107°45'16"E, altitude: 780 m, collected by Kociolek J.P. & Wang Q.X., on October 2, 2015.
Description.
Light microscopy (LM) (Fig. 1A–P). Valves elliptical with slightly constricted ends and rostrate extensions. Valve dimensions (n = 60): Length 11.5–19.0 µm, width 5.5–7.5 µm. Rapheless Valve (Fig. 1A–H): Axial area narrow, straight, linear. Central area features a U-shaped hyaline region widening towards the valve margin, on the opposite side, striae extend to the axial area. Striae radiate along the valve outline, 14–17 in 10 µm (measured opposite hyaline area). Raphe Valve (Fig. 1I–P): Axial area narrow, straight, linear. Central area rectangular to nearly circular, bordered by 3–4 shortened, asymmetrical striae on each side. Raphe branches straight. Striae radiate along the valve outline, 14–18 in 10 µm.
Scanning electron microscopy (SEM) (Figs 1Q, R, 2A, B). Rapheless Valve (Fig. 1Q, R): Striae composed of 3–4 rows of circular areolae, the outer two rows larger than those near the center. Striae narrow to 2 rows towards the axial area and may expand to 3–4 rows near the valve margin, a larger areola marks the end of striae near the valve margin (Fig. 1Q, white arrow). The central and axial areas feature irregular, shallow, linear depression. Internally, areolae are covered. A clearly visible sinus is present on one side of the central area, forming a shallow circular depression (Fig. 1R, white arrow). Raphe Valve (Fig. 2A, B): Striae composed of 3–4 rows of circular areolae. Striae narrow towards one end near the axial area. The central area typically displays 2 to 3 shortened striae. Striae almost extend onto the valve margin. Raphe branches are straight, with proximal raphe endings expanding into pores, surrounded by shallow, drop-like depressions. Distal raphe ends are curved in the same direction, briefly extending onto the mantle. Internally, proximal raphe endings are slightly deflected to the opposite side, distal raphe endings terminate in a small helictoglossa. Central nodule is raised, with striae wider raised virgae and sunken between them, and areolae are individually covered.
Planothidiumpseudoreichardtii sp. nov. SEM; A. External view of an entire raphe valve. B. Internal view of an entire raphe valve, scale bar as shown. Scale bar as shown.
Etymology.
The species was named for its outline being similar to P.reichardtii.
PhycoBank registration.
Distribution and ecology.
This species has currently only been found at its type locality, where it was collected from rocks in a rapid stream (water temperature 18.0 °C, elevation 780 m). The species was observed in sample GZ201510041P (2.0%), GZ201510045 (2.5%). In the type material (GZ201510045), P.pseudoreichardtii was rare. The associated diatom flora included Naviculalundii Reichardt (19.5%), Achnanthidiumdelmontii Pérès, le Cohu & Barthès (12.0%), Planothidiumrostratum (Østrup) Lange-Bertalot (6.0%), Platessahustedtii Lange-Bertalot (5.5%), and Achnanthidiumminutissimum (Kützing) Czarnecki (5.0%), and other taxa with lower abundance (less than 5%) such as Naviculacapitatoradiata Germain, Achnanthidiumrivulare Potapova & Ponader, Staurosiraconstruens Ehrenberg, Punctastriatanyingchiensis Luo & Wang, Cocconeisplacentula Ehrenberg, and Nitzschiapalea Smith. Additional ecological information is presented in Table 1.
Remarks.
Due to their similar valve outlines, Planothidiumpseudoreichardtii sp. nov. and Planothidiumliboensis sp. nov. were compared with several morphologically similar taxa, including Planothidiumreichardtii Lange-Bertalot & Werum, Planothidiumrostratum (Østrup) Lange-Bertalot and Planothidiumxinguense Morais, Wetzel & Bicudo (Table 2). However, P.pseudoreichardtii sp. nov. can be easily distinguished from P.liboensis sp. nov. because the former possesses a sinus, while the latter has a cavum on the center of the rapheless valve. Meanwhile, P.pseudoreichardtii can be differentiated from P.reichardtii by several distinct features: (1) in LM, the central area of the rapheless valve in P.pseudoreichardtii has a broader hyaline area outlined by 3–5 striae, whereas in P.reichardtii, the hyaline area is outlined by only 1–2 striae; (2) The striae on the rapheless valve of P.pseudoreichardtii consist of 3–4 rows of circular areolae, with the outermost two rows being larger than the central ones, whereas P.reichardtii has uniformly sized areolae in its striae; (3) the striae of P.pseudoreichardtii terminates at the valve margin with a distinctly enlarged areola, while P.reichardtii doesn’t have it. (4) irregular shallow slit-like depressions are present in the central and axial areas of the rapheless valve in P.pseudoreichardtii, whereas no depressions were observed on the rapheless valve of P.reichardtii.
Planothidium
liboensis
Taxon classificationPlantaeAchnanthalesAchnanthidiaceae
Q-M. You, P. Yu & J.P. Kociolek sp. nov.
233C8D09-6BE2-5097-A314-A5F236BF6C22
Holotype.
SHTU! Slide GZ201510051, holotype illustrated in Fig. 3F, N. Diatom samples are housed in the Lab of Algae and Environment, College of Life Sciences, Shanghai Normal University, China.
Isotype.
COLO! Material 11117, Slides are housed in the Kociolek Collection, University of Colorado, Museum of Natural History Diatom Herbarium, Boulder, U.S.A.
Type locality.
China. Xiaoqikong Scenic Area, Libo County, Guizhou Province, 25°15'36"N, 107°45'16"E, altitude: 780 m, collected by Wang Q.X. & Kociolek J.P., on October 2, 2015.
Description.
Light microscopy (LM) (Fig. 3A–P). Valves elliptical with slightly constricted ends and rostrate extensions. Valve dimensions (n = 75): Length 12–18 µm, width 5–7 µm. Rapheless Valve (SV) (Fig. 3A–H): Axial area narrow, straight, linear. Central area with a large, unilateral, horseshoe-shaped hyaline area containing a clearly visible cavum, on the opposite side, striae extend to the axial area. Striae radiate along the valve outline, 15–16 in 10 µm (measured opposite the cavum). Raphe Valve (RV) (Fig. 3I–P): Axial area narrow, straight, and linear, widening slightly towards the central area. Central area irregular, rectangular to slightly rounded, bordered by 2–3 shortened, asymmetrical striae on each side. Raphe branches straight, with proximal raphe endings drop-like. Striae radiate along the valve outline, 14–17 in 10 µm.
Planothidiumliboensis sp. nov. LM; A–H. Rapheless valves; I–P. Raphe valves. “=” indicates the different valves of the same frustule. Q.SEM external view of an entire rapheless valve, scale bar as shown. Scale bars: 10 µm (A–P).
Scanning electron microscopy (SEM) (Figs 3Q, 4A–C). Rapheless Valve (Figs 3Q, 4A): Striae composed of 4 to 5 rows of circular areolae, typically parallel to each other. Near the central area, striae often narrow to 3 areolae adjacent to the axial area. Striae extend onto the valve margin, and no areolae exist on the mantle. The axial area and central area exhibit irregular depressions. The cavum opening is broad, striae are distinctly wider than the virgae and sunken between them, areolae are covered by individual hymenes. Raphe Valve (Fig. 4B, C): Striae composed of 4 to 5 rows of circular areolae, with some striae consisting of 3 areolae near the axial area and valve margin. Striae extend onto the valve margin, and no areolae exist on the mantle. The central area typically displays 2 to 3 shortened areolae. Raphe branches are straight, with proximal raphe endings expanding into pores, surrounded by shallow, drop-like depressions. Distal raphe ends are curved in the same direction, briefly extending onto the mantle. Internally, proximal raphe endings are slightly deflected to the opposite side, distal raphe endings terminate in a small helictoglossa. The central nodule is raised, striae are markedly wider than the virgae and sunken between them, and areolae are covered by individual hymenes.
Planothidiumliboensis sp. nov SEM; A. Internal view of an entire rapheless valve; B. External view of an entire raphe valve; C. Internal view of an entire raphe valve. Scale bar as shown.
Etymology.
The species was named for the type locality, Libo County.
PhycoBank registration.
Distribution and ecology.
This species has currently only been found at its type locality, where it was collected from rocks and floating debris in ponds, as well as from rocks beneath a waterfall (water temperature 18.0–19.5 °C, elevation 629–780 m). The species was observed in samples GZ201510045 (1.0%), GZ201510051 (5.8%), and GZ201510066 (1.5%). In the type material (GZ201510051), P.liboensis exhibited a relatively high abundance. The associated diatom flora included Sellaphora sp. (15.5%), Planothidiumrostratum (12.5%), Fallacia sp. (5.0%), Achnanthidiumminutissimum (5.0%), Nitzschiapalea (5.0%), and other taxa with lower abundance (less than 5%) such as Planothidiumellipticum (Cleve) Round & Bukhtiyarova, Sellaphorarotunda Wetzel, Ector, Van de Vijver, Compère & Mann, Achnanthidium spp., Punctastriatanyingchiensis, Staurosiraconstruens, Cocconeisplacentula, and Platessahustedtii. Additional ecological information is presented in Table 1.
Remarks.
In LM, P.liboensis, P.rostratum, and P.xinguense exhibit broadly similar valve outlines, and all display a cavum. However, P.liboensis can be distinguished from other taxa by the following features: (1) P.liboensis has a denser arrangement of striae, typically consisting of 4–5 rows of similarly-sized round areolae, and the striae are distinctly wider than the virgae. In contrast, the striae of P.rostratum in the rapheless valve consist of 3–4 rows of areolae, where the two middle rows are smaller than the outer rows. The striae of P.xinguense in rapheless valve are composed of 1–3 rows of areolae; (2) The central area of rapheless valve in P.liboensis exhibits distinctive irregularly shaped shallow depressions, whereas P.rostratum features shallow, slit-like depressions. P.xinguense also exhibits irregularly shaped shallow depressions in the central area, but their shape differs from those in P.liboensis. Additionally, P.xinguense has slit-like depressions in the axial area; (3) compared to P.rostratum, the cavum of P.liboensis has a more open aperture; (4) compared to P.xinguense, P.liboensis exhibits weaker striae radiation and a narrower central area on the rapheless valve.
Planothidium
angustirostratum
Taxon classificationPlantaeAchnanthalesAchnanthidiaceae
Q-M. You, P. Yu & J.P. Kociolek sp. nov.
50A32D80-7886-5F32-AC92-846E8B398F8E
Holotype.
SHTU! Slide GZ201510051, holotype illustrated in Fig. 5F, N. Diatom samples are housed in the Lab of Algae and Environment, College of Life Sciences, Shanghai Normal University, China.
Planothidiumangustirostratum sp. nov. LM; A–H. Rapheless valves; I–P. Raphe valves. “=” indicates the different valves of the same frustule. Q.SEM external view of an entire rapheless valve, scale bar as shown. Scale bars: 10 µm (A–P).
Isotype.
COLO! Material 11117, Slides are housed in the Kociolek Collection, University of Colorado, Museum of Natural History Diatom Herbarium, Boulder, U.S.A.
Type locality.
China. Xiaoqikong Scenic Area, Libo County, Guizhou Province, 25°15'36"N, 107°45'16"E, altitude: 780 m, collected by Wang Q.X. & Kociolek J.P., on October 2, 2015.
Description.
Light microscopy (LM) (Fig. 5A–P). Valves elliptic-lanceolate with narrowly rostrate to subcapitate apices. Valve dimensions (n = 40): Valve length 17–21 µm, width 6.5–7.0 µm. Rapheless Valve (Fig. 5A–H): Axial area narrow, linear, slightly broadened in the middle. A horseshoe-shaped hyaline area present on one side of the central area, containing a cavum. Striae weakly radiate along the valve outline, 13–14 in 10 µm (measured opposite the hyaline area). Raphe Valve (Fig. 5I–P): Axial area narrow, straight, linear. Central area subcircular to rectangular, bordered by 3–4 slightly shortened striae on each side. Raphe branches straight, with proximal raphe endings drop-like. Striae radiate along the valve outline, 13–14 in 10 µm.
Scanning electron microscopy (SEM) (Figs 5Q, 6A–C). Rapheless Valve (Figs 5Q, 6A): Striae composed of 3–4 rows of circular areolae, narrowing near the central area adjacent to the axial area. Striae extend onto the valve mantle. Irregular circular depressions present between the axial area, central area, even between striae. Internally, areolae covered individually, striae wider than virgae and sunken between them. Cavum opening slightly constricted. Raphe Valve (Fig. 6B, C): Striae composed of 3–4 rows of areolae, striae near the central area regularly shortened and narrowing towards the axial area. Striae extend onto the valve mantle. Raphe branches straight, with proximal raphe endings expanding into pores surrounded by drop-like depressions. Distal raphe ends are curved in the same direction, slightly extending onto the valve mantle. Internally, proximal raphe endings slightly deflected to the opposite side, and distal raphe endings terminating in a faint helictoglossa. Internally, areolae covered individually, striae wider than virgae and sunken between them.
Planothidiumangustirostratum sp. nov. SEM; A. Internal view of an entire rapheless valve; B. External view of an entire raphe valve; C. Internal view of an entire raphe valve. Scale bar as shown.
Etymology.
The species was named for its narrowly rostrate valve apices.
PhycoBank registration.
Distribution and ecology.
This species has currently only been found at its type locality, where it was collected from floating debris in a pond (water temperature 18.0 °C, elevation 780 m). The species was observed only in sample (GZ201510051) at a low relative abundance (0.75%). In the type material, P.angustirostratum was rare. The associated diatom flora included Sellaphora sp. (15.5%), Planothidiumrostratum (12.5%), Planothidiumliboensis (5.75%), Fallacia sp. (5.0%), Achnanthidiumminutissimum (5.0%), Nitzschiapalea (5.0%), and other taxa with lower abundance (less than 5%) such as Planothidiumellipticum, Sellaphorarotunda, Achnanthidium spp., Punctastriatanyingchiensis, Staurosiraconstruens, Cocconeisplacentula, and Platessahustedtii. Additional ecological information is presented in Table 1.
Remarks.
Due to the broadly similar valve outline in LM and the presence of a cavum, Planothidiumangustirostratum sp. nov. can be compared with Planothidiumbrasiliense Wetzel & Blanco, Planothidiumpotapovae Wetzel & Ector and Planothidiumrostratoholarcticum Lange-Bertalot & Bąk (Table 3). In addition to differences in striae density and valve length and width, Planothidiumangustirostratum sp. nov. also can be distinguished from these similar species by the following features: (1) with narrower rostrate to subcapitate apices and a narrower cavum aperture; (2) a smaller central area, ranging from rectangular to nearly circular; (3) its striae denser on the raphe valve, consist of 3–4 rows of circular areolae; (4) circular shallow depressions are present in the central area, axial area, and between the striae, unlike the slit-like depressions observed in P.potapovae and P.rostratoholarcticum.
Planothidium
maolanensis
Taxon classificationPlantaeAchnanthalesAchnanthidiaceae
Q-M. You, P. Yu & J.P. Kociolek sp. nov.
C92B2E99-9C1F-5D70-A6AC-6EF828CC24C1
Holotype.
SHTU! Slide GZ201510099, holotype illustrated in Fig. 7D, L. Diatom samples are housed in the Lab of Algae and Environment, College of Life Sciences, Shanghai Normal University, China.
Planothidiummaolanensis sp. nov. LM; A–H. Rapheless valves; I–P. Raphe valves. “=” indicates the different valves of the same frustule. Q.SEM external view of an entire rapheless valve, scale bar as shown. Scale bars: 10 µm (A–P).
Isotype.
COLO! Material 11165, Slides are housed in the Kociolek Collection, University of Colorado, Museum of Natural History Diatom Herbarium, Boulder, U.S.A.
Type locality.
China. Maolan Nature Reserve, Libo County, Guizhou Province, 25°17'32"N, 108°04'16"E, altitude: 650 m, collected by Kociolek J.P. & Wang Q.X., on October 4, 2015.
Description.
Light microscopy (LM) (Fig. 7A–P). Valves elliptical, with rounded ends not extended. Valve dimensions (n = 50): Length 9.5–18.0 µm, width 5.5–7.5 µm. Rapheless Valve (Fig. 7A–H): Axial area linear-lanceolate, with central area slightly broadened. One side of the central area has two striae with widened spacing, opposite side striae slightly shortened. Striae weakly radiate along the valve outline, narrowing towards axial area, 10–12 in 10 µm (measured opposite hyaline area). Raphe Valve (Fig. 7I–P): Axial area linear-lanceolate, slightly widened in the central area. Central area subcircular, bordered by 2–3 slightly shortened striae on each side. One side of the central area has two striae with widened spacing. Striae radiate weakly, 10–12 in 10 µm. Raphe branches straight, occasionally slightly curved.
Scanning electron microscopy (SEM) (Figs 7Q, 8A–C). Rapheless Valve (Figs 7Q, 8A): Striae composed of 4–5 rows of circular areolae, markedly narrow near the axial area, reducing to 2–3 rows of areolae, striae almost extend onto the valve margin, and some areolae exist on the mantle. Irregular depressions present on the axial area, central area and even on the virgae. Internally, areolae covered. Virgae widen gradually from the ends towards the center, with striae sunken between virgae, a distinctly wider virgae present on one side of the central area (Fig. 8A, White arrow). Raphe Valve (Fig. 8B, C): Striae composed of 5–6 rows of small circular areolae, markedly narrowing near the central area, reducing to 2–3 rows of areolae. striae don’t extend onto the valve margin, and some areolae exist on the mantle (Fig. 8B, White arrows). Raphe branches straight, occasionally slightly curved. Externally, proximal raphe endings expand into pores, distal raphe ends are curved in the same direction, and extend onto the mantle. Internally, proximal raphe endings slightly deflected to the opposite side, and distal raphe endings terminate in a small helictoglossa. Internally, areolae covered individually by membranes, striae sunken between virgae.
Planothidiummaolanensis sp. nov SEM; A. Internal view of an entire rapheless valve. The white arrow points to a distinctly wider virgae; B. External view of an entire raphe valve. White arrows indicate the areolae on the mantle. C. Internal view of an entire raphe valve. Scale bar as shown.
Etymology.
The species was named for the Maolan Nature Reserve, where the type specimen was collected.
PhycoBank registration.
Distribution and ecology.
This species has currently only been found at its type locality, where it was collected from rocks in a subsurface stream and in rapidly flowing river water (water temperature 18.0 °C, elevation 650–811 m). The species was observed in samples GZ201510099 (3.75%), GZ201510100 (0.25%), and GZ2015108 (0.75%). In the type material (GZ201510099), P.maolanensis was rare. The associated diatom flora included Cocconeisplacentula (23.0%), Achnanthidiumdelmontii (7.5%), Naviculalundii (7.5%), Naviculaantonii Lange-Bertalot, Navicula sp. (6.0%), Achnanthidiumminutissimum (5.5%), and other taxa with lower abundance (less than 5%) such as Platessahustedtii, Achnanthidium spp., Encyonemahophense Krammer, and Achnanthesprominula Levkov & Tofilovska. Additional ecological information is presented in Table 1.
Remarks.
Due to the broadly similar valve outline in LM and the absence of cavum, Planothidiummaolanensis sp. nov. can be compared with P.hauckianum (Grunow) Bukhtiyarova, P.kaetherobertianum Van de Vijber & Bosak, and P.iberense Rovira & Witkowski (Table 4). P.maolanensis can be easily distinguished from P.kaetherobertianum and P.iberense by valve outline and striae density, while it shows greater overall similarity to P.hauckianum. However, P.maolanensis can still be differentiated by the following features: (1) In P.maolanensis, only two striae on one side of the central area show slightly increased spacing, while the remaining striae are evenly arranged. In contrast, P.hauckianum has more striae with increased spacing on both sides of the central area. (2) The striae of P.maolanensis consist of 4–5 rows of circular areolae on rapheless valve and 5–6 rows on raphe valve. In P.hauckianum, the striae consist of 3–4 rows of circular areolae on the rapheless valve and 2–4 rows on the raphe valve. (3) Although the striae in P.maolanensis and P.hauckianum both generally narrow toward the axial area, the narrowing in P.maolanensis is not gradual or consistent. Sometimes, the middle of striae contracts, making it narrower than the ends near the axial area. (4) P.maolanensis has irregularly shaped shallow depressions in axial area and between the striae, while P.hauckianum exhibits slit-like depressions.
Discussion
The four new species described in this study exhibit multiseriate striae on both the raphe valve and the rapheless valve. On one side of the central area of the rapheless valve, Planothidiumpseudoreichardtii sp. nov. features an uncovered depression known as a “sinus”, Planothidiumliboensis sp. nov. and Planothidiumangustirostratum sp. nov. possess a cavum, and Planothidiummaolanensis sp. nov. lacks both sinus and cavum. The four new species were compared with several related and morphologically similar taxa (Tables 2, 3, 4).
As a complement to morphometry, Wetzel et al. (2019) proposed several distinguishing features in SEM, such as the pattern of valve surface smoothness on the rapheless valve, the number of areolae per stria, and whether the striae on the rapheless valve are interrupted towards the valve mantle. During the observation of these new Planothidium species, we found that the morphology of the striae and the number of areolae rows per stria on the rapheless valve are relatively stable within the same species. For instance, the rapheless valve striae of P.pseudoreichardtii sp. nov. are composed of two outer rows of larger areolae and 1–2 smaller rows in the center, with an enlarged areola present at the end of each stria. The presence or absence of striae on the rapheless valve mantle is also stable within the same species. Striae are present on the valve mantle of P.angustirostratum sp. nov. and P.maolanensis sp. nov., whereas no striae are observed on the valve mantle of P.liboensis sp. nov. and P.maolanensis sp. nov. Furthermore, we observed that the feature of valve surface smoothness on the rapheless valve is consistent within each species. Based on the shape of the depressions, these features can be roughly classified into three types: P.angustirostratum sp. nov. exhibits circular shallow depressions on the valve surface; P.liboensis sp. nov. and P.maolanensis sp. nov. display irregularly shaped depressions; while P.pseudoreichardtii sp. nov. and P.rostratum, also observed in the Xiaoqikong Scenic Area, exhibit slit-like shallow depressions.
Currently, numerous species of the genus Planothidium have been discovered in various aquatic environments and habitats worldwide (Blanco et al. 2013; N’Guessan et al. 2014; Wetzel and Ector 2014a, 2014b; Kopalová et al. 2016; Riaux-Gobin et al. 2018; Stancheva 2019; Cantonati et al. 2021; Lai et al. 2021). Additionally, some studies have investigated the ecological distribution and physiological characteristics of Planothidium species (Sbihi et al. 2014; Stancheva et al. 2020; Juchem et al. 2023). Despite significant advancements over the years, the taxonomy of Planothidium is relatively well-developed, but several questions remain open for further exploration. Future research may require additional populations or increased taxa sampling to examine the stability of certain morphological features within the genus. Furthermore, integrating more ecological data and molecular techniques could provide deeper insights into the variability of these features under different environmental conditions and their taxonomic applicability.
Supplementary Material
XML Treatment for Planothidium pseudoreichardtii
XML Treatment for Planothidium liboensis
XML Treatment for Planothidium angustirostratum
XML Treatment for Planothidium maolanensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Álvarez-Blanco I Blanco S (2013) Planothidiumgalaicum sp. nov. (Bacillariophyta, Achnanthidiaceae), a new diatom species from Galician coast, Spain. Phytotaxa 151: 44. 10.11646/phytotaxa.151.1.4 · doi ↗
- 2Bąk M Lange-Bertalot H (2014) Four small-celled Planothidium Species from Central Europe proposed as new to science.Oceanological and Hydrobiological Studies 43: 346–359. 10.2478/s 13545-014-0152-9 · doi ↗
- 3Blanco SÁlvarez‐Blanco I Cejudo‐Figueiras C Espejo JMR Barrera CBBécares E Del Olmo FD Artigas RC (2013) The diatom flora in temporary ponds of Doñana National Park (southwest Spain): Five new taxa.Nordic Journal of Botany 31: 489–499. 10.1111/j.1756-1051.2013.01691.x · doi ↗
- 4Borrego-Ramos M Blanco S Olenici A (2018) Diatoms from the valporquero cave (Leon, NW Spain), with the description of Germainiellalegionensis sp. nov.Journal of Caves and Karst Studies 80: 181–189. 10.4311/2017 MB 0128 · doi ↗
- 5Cantonati M Bilous O Angeli N Van Wensen L Lange-Bertalot H (2021) Three new diatom species from spring habitats in the Northern Apennines (Emilia-Romagna, Italy). Diversity 13: 549. 10.3390/d 13110549 · doi ↗
- 6Guiry MD Guiry GM (2024) Algae Base. World-wide electronic publication, National University of Ireland, Galway. https://www.algaebase.org [Searched on 05 December 2024]
- 7Jahn R Abarca N Gemeinholzer B Mora D Skibbe O Kulikovskiy M Gusev E Kusber W-H Zimmermann J (2017) Planothidiumlanceolatum and Planothidiumfrequentissimum reinvestigated with molecular methods and morphology: Four new species and the taxonomic importance of the sinus and cavum.Diatom Research 32: 75–107. 10.1080/0269249 X.2017.1312548 · doi ↗
- 8Juchem DP Schimani K Holzinger A Permann C Abarca N Skibbe O Zimmermann J Graeve M Karsten U (2023) Lipid degradation and photosynthetic traits after prolonged darkness in four Antarctic benthic diatoms, including the newly described species Planothidiumwetzelii sp. nov. Frontiers in Microbiology 14: 1241826. 10.3389/fmicb.2023.1241826 PMC 1050092937720158 · doi ↗ · pubmed ↗