Rhizophorastylosa (Rhizophoraceae) newly recorded from Thailand: lectotypification, leaf anatomy, and pollen morphology

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
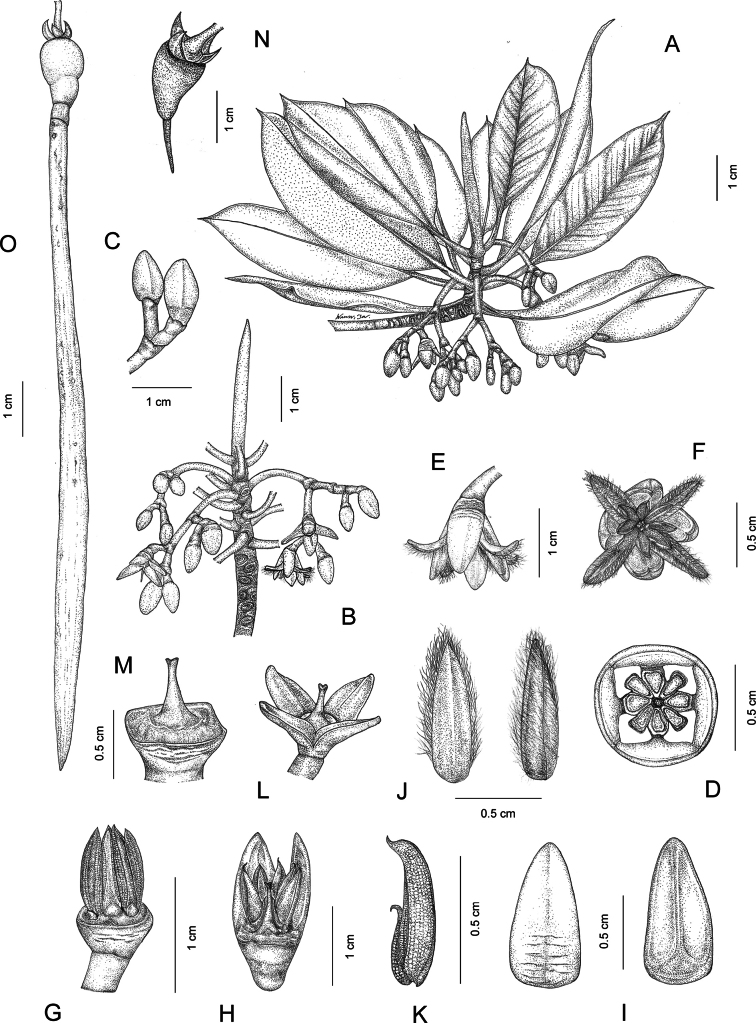 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3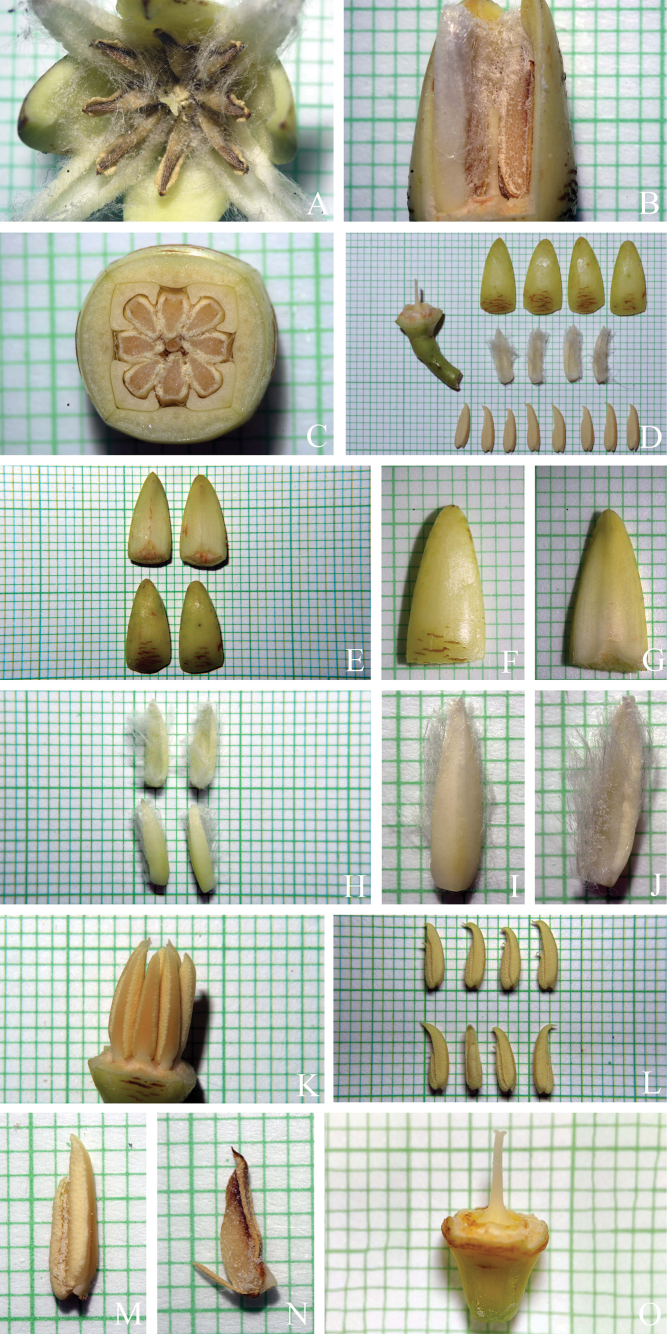 Figure 4
Figure 4 Figure 5
Figure 5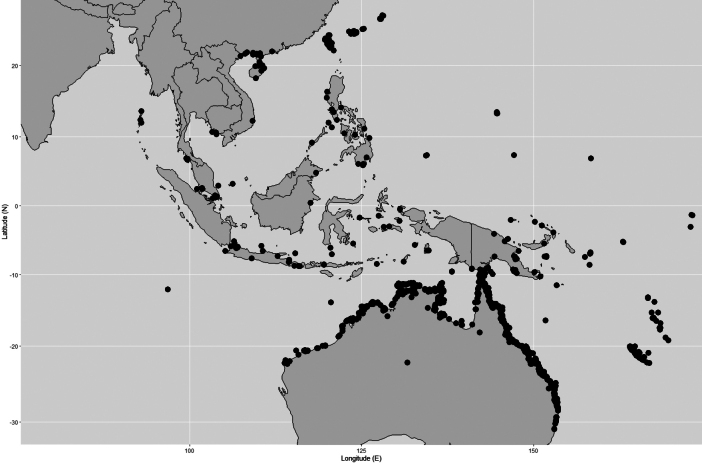 Figure 6
Figure 6 Figure 7
Figure 7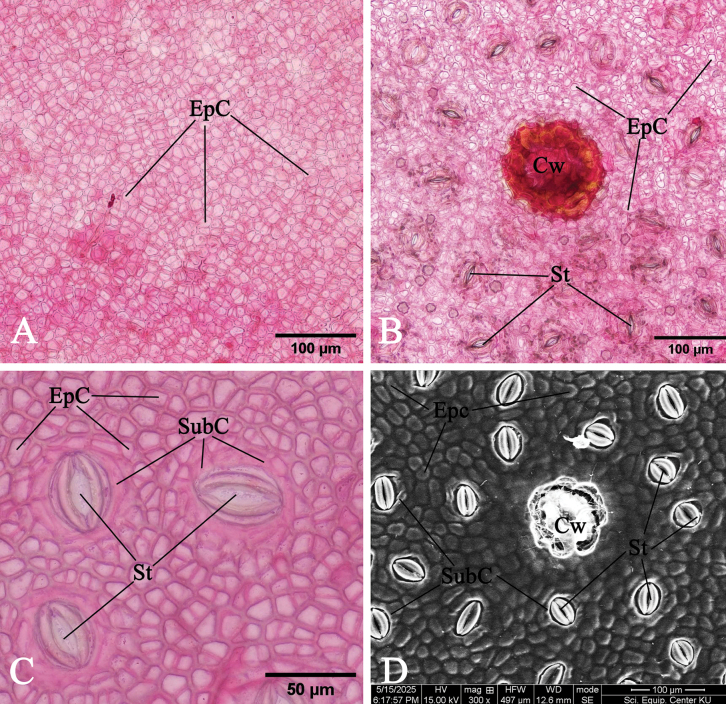 Figure 8
Figure 8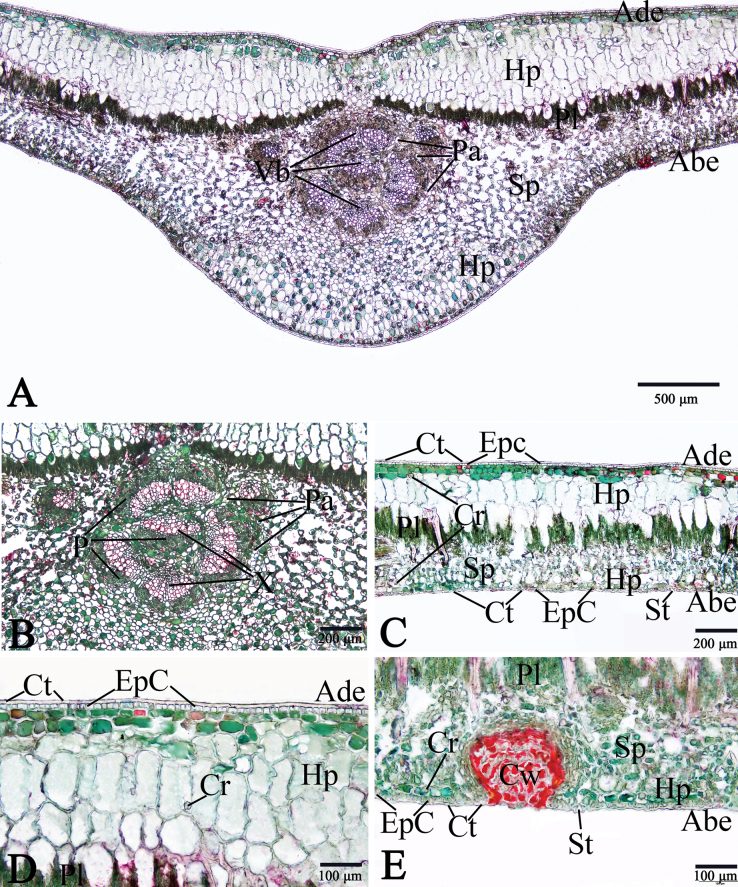 Figure 9
Figure 9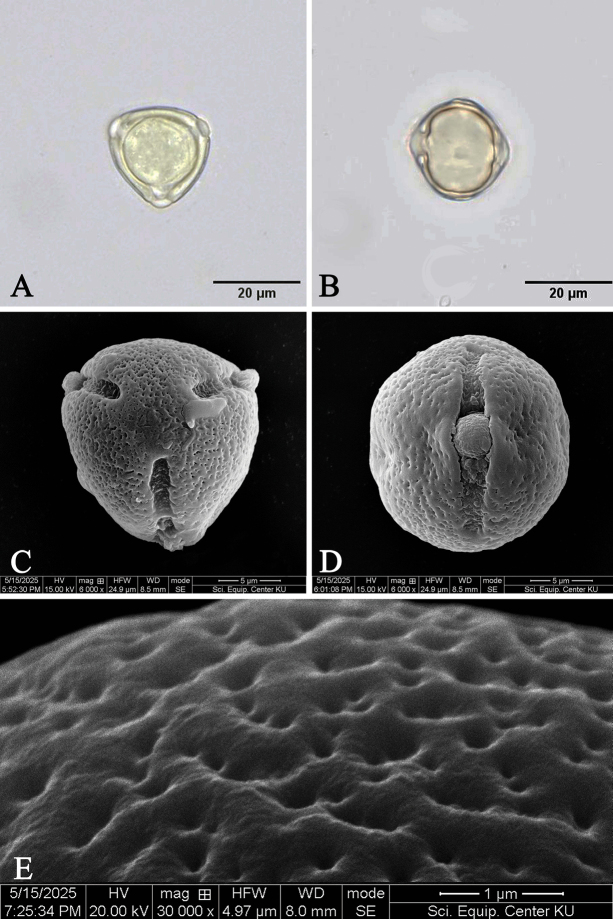 Figure 10
Figure 10| Measurements of vegetative and reproductive parts (units) | Sample sizes | Ranges | Mean ± SD |
|---|---|---|---|
| Stipule length (cm) | 80 | 3.5–7.5 | 5.40 ± 0.92 |
| Stipule diameter at the base (mm) | 80 | 2.5–9.0 | 4.20 ± 1.58 |
| Colleter length (mm) | 50 | 0.5–1.2 | 0.72 ± 0.16 |
| Colleter diameter at the base (mm) | 50 | 0.2–0.5 | 0.35 ± 0.08 |
| Leaf length (cm) | 300 | 8.0–16.0 | 11.43 ± 1.64 |
| Leaf width (cm) | 300 | 3.0–8.5 | 5.20 ± 1.02 |
| Leaf length/width ratio | 300 | 1.7–2.9 | 2.22 ± 0.19 |
| Leaf lamina apex length (mm) | 150 | 2.0–6.0 | 4.30 ± 1.04 |
| Number of secondary veins each side | 300 | 8–14 | 11.20 ± 1.68 |
| Petiole length (cm) | 300 | 2.0–4.8 | 3.25 ± 0.55 |
| Petiole diameter at the middle (mm) | 300 | 2.0–4.5 | 3.19 ± 0.44 |
| Inflorescence length (cm) | 180 | 5.0–12.5 | 7.93 ± 1.35 |
| Peduncle length (cm) | 180 | 2.3–6.0 | 3.80 ± 0.82 |
| Peduncle diameter at the middle (mm) | 180 | 2.0–3.5 | 2.54 ± 0.34 |
| Number of flowers per inflorescence | 180 | 4–13 | 6.70 ± 1.57 |
| Mature flower bud length (cm) | 200 | 0.9–1.5 | 1.25 ± 0.10 |
| Mature flower bud diameter at the basal part (cm) | 200 | 4.0–8.0 | 6.41 ± 0.82 |
| Mature flower bud length/diameter ratio | 200 | 1.5–2.5 | 1.97 ± 0.23 |
| Fully open flower diameter (cm) | 100 | 1.2–1.8 | 1.47 ± 0.15 |
| Sepal length (cm) | 200 | 0.7–1.3 | 1.01 ± 0.14 |
| Sepal width at the base (mm) | 200 | 4.0–6.0 | 4.89 ± 0.59 |
| Sepal length/width ratio | 200 | 1.7–2.7 | 2.06 ± 0.19 |
| Petal length (cm) | 200 | 0.6–1.0 | 0.79 ± 0.12 |
| Petal width at the middle (cm) | 200 | 1.5–3.5 | 2.30 ± 0.51 |
| Petal length/width ratio | 200 | 2.2–5.4 | 3.51 ± 0.62 |
| Anther length (mm) | 200 | 3.5–8.5 | 6.14 ± 1.14 |
| Anther width at the middle (mm) | 200 | 1.2–2.8 | 1.70 ± 0.40 |
| Free part of the ovary length (mm) | 100 | 0.7–1.2 | 0.97 ± 0.12 |
| Free part of the ovary diameter (mm) | 100 | 1.2–1.8 | 1.47 ± 0.15 |
| Style length (mm) | 170 | 3.0–5.0 | 3.80 ± 0.52 |
| Pedicel length (cm) | 100 | 0.6–1.0 | 0.82 ± 0.14 |
| Pedicel diameter (mm) | 100 | 2.0–3.0 | 2.58 ± 0.26 |
| Fruit length (cm) (hypocotyls nearly come off) | 70 | 2.2–4.0 | 2.91 ± 0.43 |
| Fruit diameter at the basal part (cm) | 70 | 1.7–3.0 | 2.04 ± 0.23 |
| Fruit diameter at the apical part (cm) | 70 | 0.9–1.7 | 1.04 ± 0.17 |
| Persistent sepal length (cm) | 150 | 0.9–1.1 | 0.98 ± 0.04 |
| Persistent sepal width at the base (mm) | 150 | 5.0–8.5 | 6.49 ± 0.67 |
| Infructescence stalk length (cm) | 60 | 2.0–6.0 | 3.26 ± 0.85 |
| Infructescence stalk diameter (mm) | 60 | 2.0–3.0 | 2.54 ± 0.33 |
| Fruit stalk length (cm) | 60 | 0.6–1.7 | 1.03 ± 0.25 |
| Fruit stalk diameter (mm) | 60 | 3.0–5.0 | 3.66 ± 0.39 |
| Hypocotyl length (cm) | 75 | 19.5–40.0 | 30.01 ± 3.25 |
| Hypocotyl diameter at the apical part (mm) | 75 | 6.5–9.0 | 7.64 ± 0.65 |
| Hypocotyl diameter at the widest part (cm) | 75 | 1.0–1.6 | 1.29 ± 0.12 |
| Hypocotyl diameter at the basal part (cm) | 75 | 0.8–1.3 | 0.99 ± 0.10 |
| Cotyledonous tube diameter (cm) | 75 | 0.6–1.2 | 0.86 ± 0.14 |
| Morphological characters |
|
|
|---|---|---|
| Habit | Small trees, single- to multi-stemmed, 2.5–8 m tall | Small to large trees, 2–20 (–30) m tall, usually single-stemmed |
| Stipule colours and sizes | Pale green or green, turning whitish pale green or creamish white before the leaves emerging or before falling off, smaller, 3.5–7.5 cm long and 2.5–9 mm diam at the base | Pale green, reddish pale green or pale greenish red when young, turning pale yellow or reddish pale yellow before the leaves emerging or before falling off, larger, 4.5–12.5 cm long, 0.5–1.7 cm diam at the base |
| Leaf shapes and sizes | Elliptic, smaller, 8–16 × 3–8.5 cm, widest at the middle part | Ovate or lanceolate-ovate, larger, 13–23.5 × 6–13 cm, widest at the basal part |
| Petal shapes and hairs | Narrowly elliptic or elliptic, densely and longer villous along margins | Lanceolate-ovate or lanceolate, densely and shorter villous along margins |
| Free part of the ovary length | Shorter, 0.7–1.2 mm long | Longer, 1.5–3 mm long |
| Style length | 3–5 mm long | Very short |
| Fruit sizes (ovoid or conical-ovoid, before seed germination) | Smaller, 2–2.7 cm long, 1–1.7 cm diam. at the basal part | Larger, 3.7–5 cm long, 2.1–2.8 cm diam. at the basal part |
| Fruit sizes (obpyriform, when the hypocotyls nearly come off) | Smaller, 2.2–4 cm long, 1.7–3 cm diam. at the basal part | Larger, 4–7 cm long, 2.4–3.5 cm diam. at the basal part |
| Hypocotyl length | Shorter, 19.5–40 cm long | Longer, 25–70 cm long |
| Morphological characters (units) | Sample sizes | ||||
|---|---|---|---|---|---|
| Ranges | Mean ± SD | Ranges | Mean ± SD | ||
| Stipule length (cm) | A 30/B 50 | 4.0–7.5 | 5.78 ± 1.03 | 3.5–6.5 | 5.16 ± 0.77 |
| Stipule diameter at the base (mm) | A 30/B 50 | 3.0–9.0 | 5.06 ± 2.04 | 2.5–6.0 | 3.68 ± 0.91 |
| Leaf lamina length (cm) | 150 | 9.5–16.0 | 12.72 ± 1.10 | 8.0–12.5 | 10.14 ± 0.94 |
| Leaf lamina width (cm) | 150 | 4.5–8.5 | 6.01 ± 0.76 | 3.0–5.5 | 4.39 ± 0.44 |
| Petiole length (cm) | 150 | 2.5–4.8 | 3.63 ± 0.44 | 2.0–4.0 | 2.88 ± 0.36 |
| Petiole diameter at the middle (mm) | 150 | 2.5–4.5 | 3.45 ± 0.38 | 2.0–3.5 | 2.93 ± 0.31 |
| Inflorescence length (cm) | 90 | 6.0–12.5 | 8.80 ± 1.18 | 5.0–9.0 | 7.05 ± 0.87 |
| Peduncle length (cm) | 90 | 2.8–6.0 | 4.39 ± 0.65 | 2.3–4.8 | 3.22 ± 0.49 |
| Fruit length (cm) (hypocotyls nearly come off) | A 20/B 50 | 3.0–4.0 | 3.49 ± 0.31 | 2.2–3.0 | 2.68 ± 0.17 |
| Fruit diameter at the basal part (cm) | A 20/B 50 | 1.9–3.0 | 2.25 ± 0.31 | 1.7–2.3 | 1.95 ± 0.11 |
| 1 | Inflorescences on leafless branchlets (in the axils of leaf scars), 2-flowered cymes; flowers sessile; flower buds pale green, usually tinged with brown, finely reticulated, broadly ellipsoid or ellipsoid; bracteoles brown, connate, cup-shaped, shallowly lobed; sepals elliptic-oblong, thickly coriaceous; petals flattened, not involute, glabrous; stamens mostly 12; fruits up to 1.8 cm diam. at the basal part (when the hypocotyls nearly come off); hypocotyls dark green or purplish dark green; cotyledonous cylindrical tubes red, dark greenish red or reddish dark green; stipules bearing bright yellow colleters that produce a sticky, clear exudate; the stiff, pointed tips of the leaves usually less than 3 mm long |
|
| – | Inflorescences axillary, dichotomously branched, many-flowered cymes; flowers pedicelled; flower buds pale green, turning pale yellow when mature, ovoid or conical-ovoid; bracteoles pale green, connate at the base, bilobed; sepals triangular, coriaceous; petals involute, villous along margins; stamens 8; fruits usually more than 1.8 cm diam. at the basal part (when the hypocotyls nearly come off); hypocotyls green; cotyledonous cylindrical tubes pale green or greenish pale yellow; stipules bearing pale yellow colleters that produce a sticky, white exudate; the stiff, pointed tips of the leaves usually more than 3 mm long |
|
| 2 | Leaves ovate or lanceolate-ovate, larger, 13–23.5 × 6–13 cm; petals lanceolate-ovate or lanceolate, densely and shorter villous along margins; free part of the ovaries, longer, 1.5–3 mm long; style very short; fruits (before seed germination) larger, 3.7–5 cm long, 2.1–2.8 cm diam. at the basal part; fruit (when the hypocotyls nearly come off) larger, 4–7 cm long, 2.4–3.5 cm diam. at the basal part; hypocotyls longer, 25–70 cm long; small to large trees, 2–20 (–30) m tall, usually single-stemmed |
|
| – | Leaves elliptic, smaller, 8–16 × 3–8.5 cm; petals narrowly elliptic or elliptic, densely and longer villous along margins; free part of the ovaries, shorter, 0.7–1.2 mm long; style 3–5 mm long; fruits (before seed germination) smaller, 2–2.7 cm long, 1–1.7 cm diam. at the basal part; fruit (when the hypocotyls nearly come off) smaller, 2.2–4 cm long, 1.7–3 cm diam. at the basal part; hypocotyls shorter, 19.5–40 cm long; small trees, single- to multi-stemmed, 2.5–8 m tall |
|
| Anatomical characters | From the author’s observations | Previous studies |
|---|---|---|
| Leaf thickness (μm) | 574.11–881.72 (720.53 ± 92.18, n = 20) | 571.5 ± 21.3 (Australia)[1] 556.0 ± 31.4 (Singapore)[1] |
| Leaf structure type | Bifacial leaves | Dorsiventral leaves[1] (= bifacial leaves) |
| Cuticular wax thickness on the adaxial leaf surface (μm) | 14.13–34.65 (25.46 ± 5.85, n = 20) | 7.04 ± 1.94[1] |
| Cuticular wax thickness on the abaxial leaf surface (μm) | 13.27–33.38 (23.80 ± 6.93, n = 20) | 5.25 ± 1.23[1] |
| Number of epidermal cell layer on the adaxial leaf surface | 1-layered | 1-layered[1] |
| Number of epidermal cell layer on the abaxial leaf surface | 1-layered | 1-layered[1] |
| Epidermal layer thickness on the adaxial leaf surface (μm) | 20.40–43.55 (26.97 ± 5.78, n = 20) | – |
| Epidermal layer thickness on the abaxial leaf surface (μm) | 17.25–37.31 (30.05 ± 5.96, n = 20) | – |
| Epidermal cell shapes on the adaxial leaf surface | Polygonal, sometimes trapezium | Polygonal, 29–35 × 17–25 μm [3] |
| Epidermal cell shapes on the abaxial leaf surface | Polygonal, sometimes trapezium | Polygonal[3] |
| Stomatal types | Sunken cyclocytic | Cyclocytic[1], |
| Stomatal density per mm2 | 77–153 (108.91 ± 18.19, n = 100) | 61.15 (mean), low density, 9–12 (number of stomata in one field of view, 400× magnification)[3] |
| Stomatal length (μm) | 18.31–30.48 (24.65 ± 3.42, n = 100) | 49.50 ± 2.70[1], |
| Stomatal width (μm) | 9.56–15.21 (12.20 ± 1.61, n = 100) | 43.27 ± 2.27[1], |
| Stomatal length/width ratio | 1.39–3.19 (2.05 ± 0.41, n = 100) | 1.14[1] |
| Number of subsidiary cells | 6–8 (6.84 ± 0.81, n = 50) | (5–)6–7[1] |
| Cork wart density per cm2 | 15–52 (31.60 ± 9.85, n = 300) | 10.1 ± 0.4 (mm2)[2] |
| Cork wart density at the apical part per cm2 | 30–52 (41.92 ± 6.11, n = 100) | – |
| Cork wart density at the middle part per cm2 | 24–40 (31.30 ± 4.88, n = 100) | – |
| Cork wart density at the basal part per cm2 | 15–28 (21.33 ± 4.07, n = 100) | – |
| Cork wart diameter (μm) | 41.57–94.58 (67.29 ± 15.69, n = 100) | 196.00 ± 20.86 (Singapore)[1] 252.00 ± 46.11 (Taiwan)[1] |
| Hypodermal layer thickness on the adaxial leaf surface (μm) | 206.41–361.11 (260.49 ± 43.90, n = 20) | – |
| Hypodermal layer thickness on the abaxial leaf surface (μm) | 50.67–110.54 (80.36 ± 18.41, n = 20) | – |
| Number of hypodermal cell layer on the adaxial leaf surface | 4–6-layered (5.25 ± 0.72, n = 20) | 2-layered (small-sized) and 3-layered (large-sized)[1], 5- layered[3] |
| Number of hypodermal cell layer on the abaxial leaf surface | 2–3-layered (2.45 ± 0.51, n = 20) | 1(–3)-layered[1] |
| Hypodermal cell shapes on the adaxial leaf surface (μm) | Subcircular, elliptic, oblong or rounded polygonal | Rounded hexagon [3] |
| Hypodermal cell length on the adaxial leaf surface (μm) | 31.90–99.67 (73.12 ± 20.65, n = 20) | 49–73[3] |
| Hypodermal cell width on the adaxial leaf surface (μm) | 29.69–73.84 (53.55 ± 12.55, n = 20) | 34–46[3] |
| Hypodermal cell shapes on the abaxial leaf surface (μm) | Broadly elliptic, circular or rounded polygonal | – |
| Hypodermal cell length on the abaxial leaf surface (μm) | 18.78–47.27 (33.53 ± 6.92, n = 20) | – |
| Hypodermal cell width on the abaxial leaf surface (μm) | 19.61–46.08 (31.32 ± 6.21, n = 20) | – |
| Crystal location, type and diameter (μm) | Druse (in hypodermal cells), 14.84–30.06 (22.62 ± 3.98, n = 20) | – |
| Number of palisade cell layer | 1–2-layered (1.40 ± 0.50, n = 20) | 1–4-layered (upper), 0–1-layered (lower)[1] |
| Number of spongy cell layer | 5–7-layered (6.05 ± 0.76, n = 20) | 9–10-layered (Australia), 7–8-layered (Singapore)[1] |
| Pollen characters | From the author’s observations | Previous studies |
|---|---|---|
| Polar axis [P] length (μm) | 17.93–22.36 (20.59 ± 1.35, n = 30) | 19.9 (20.5) 22.7 × 21.3 (22.4) 24.7[1]; 17.29 (19.17) 21.27[2] |
| Equatorial axis [E] length (μm) | 16.79–22.94 (20.08 ± 1.64, n = 30) | 16.61 (18.62) 20.46[2] |
| P/E ratio | 0.88–1.15 (1.03 ± 0.07, n = 30) | 1.03[2] |
| Pollen size classes | Small | – |
| Pollen shapes | Prolate spheroidal or oblate spheroidal | Subprolate, circular to subcircular[1]; prolate-speroidal[2] |
| Pollen aperture | Tricolporate | Tricolporate[1], [2] |
| Colpus length (μm) | 12.92–20.40 (16.48 ± 2.15, n = 30) | 13.4[1] |
| Colpus width (μm) | 2.35–3.85 (3.21 ± 0.37, n = 30) | – |
| Exine thickness (μm) | 0.83–2.43 (1.54 ± 0.39, n = 30) | 1.5[1]; 0.79 (0.95) 1.24[2] |
| Exine sculpturing | Reticulate | Scabrate to slightly granulate to irregularly pitted[1]; perforate[2] |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and Fungal Species Descriptions · Plant Diversity and Evolution · Plant Parasitism and Resistance
Introduction
Rhizophoraceae Pers. is well known as the most species-rich mangrove family, comprising four exclusively mangrove genera and 19 species: Bruguiera Lam. ex Savigny (six species and two natural hybrids), Ceriops Arn. (five species), Kandelia (DC.) Wight & Arn. (two species), and Rhizophora L. (six species and five natural hybrids) (Tomlinson 1986, 2016; Sheue et al. 2012; POWO 2025; WFO 2025). All members of the Rhizophoraceae have large, conspicuous, glabrous and caducous interpetiolar stipules on both vegetative and reproductive shoots, which strongly ensheath the developing leaves and inflorescences (Hou 1958). In species of mangrove Rhizophoraceae, several to hundreds of aggregated finger-like colleters mixed with milky mucilage can be observed with the naked eye at the adaxial base of the stipules (Tomlinson 1986, 2016; Sheue 2003). The structural and mechanical protection provided by stipules and colleter exudates may help shield the young shoots of these mangrove plants from their harsh environmental conditions (Sheue et al. 2012).
The tribe Rhizophoreae Bartl. is characterized by the following features: true mangrove shrubs or trees, with aerial roots; glabrous and caducous interpetiolar stipules that sheath the terminal bud and young leaves; decussate, entire leaves; cymose inflorescences; 4–16-merous flowers; mostly diplostemonous androecia (with twice as many stamens as petals); half-inferior to fully inferior ovaries, 2–3-carpellate with two ovules per carpel; baccate, fibrous fruits usually containing a single seed; and viviparous germination. This pantropical tribe comprises four genera: Bruguiera, Ceriops, Kandelia, and Rhizophora (Henslow 1878; King 1897; Hou 1958; Schwarzbach 2014).
Rhizophora is a group of tropical and subtropical mangrove trees, commonly referred to as true mangroves. It is a small genus belonging to the order Malpighiales Juss. ex Bercht. & J. Presl, the family Rhizophoraceae (The Angiosperm Phylogeny Group 2016), and the tribe Rhizophoreae (Schwarzbach 2014). Eleven taxa have been recognized within the genus, comprising six accepted species and five natural hybrids. Among these, two species and one hybrid occur in the Americas; three species and one hybrid in Africa; three species and two hybrids in the Indian subcontinent, China, and Southeast Asia (POWO 2025); three species and one hybrid in Australia (Duke and Bunt 1979; McCusker 1984; POWO 2025); and four species and three hybrids in the Pacific region (Melanesia, Micronesia, and Polynesia) (Tomlinson 1978; POWO 2025). These three species of Rhizophora found in the Indian subcontinent, China, and Southeast Asian species are R.apiculata Blume, R.mucronata Poir., and R.stylosa Griff. (Hou 1958, 1960; Qin and Boufford 2007; Marchand 2008; POWO 2025). In addition, two natural hybrids occur in the region: R.×annamalayana Kathiresan (R.apiculata × R.mucronata) and R.×lamarckii Montrouz. (R.apiculata × R.stylosa) (Parani et al. 1997; Ng and Chan 2012; Ng et al. 2013; Setyawan et al. 2014; Ragavan et al. 2015; POWO 2025).
Previously, Hou (1970) treated the genus Rhizophora in the “Flora of Thailand”, recognizing two species: R.apiculata and R.mucronata. However, this account was brief, providing limited morphological descriptions and lacking other supporting information. Recently, Ngernsaengsaruay et al. (2024) revised the genus Rhizophora for Thailand, offering updated and detailed morphological descriptions, an identification key, and comprehensive notes on distribution, habitat and ecology, phenology, conservation assessments, and lectotypifications. In addition, leaf anatomical characters and pollen morphology were examined to provide a complete overview of Rhizophora in Thailand.
In 2007, Mr.Nirat Jintana and Associate Professor Dr.Vipak Jintana discovered a plant belonging to the genus Rhizophora on Ko Lidi and Ko Bulon Le, within Mu Ko Phetra National Park, La-ngu District, Satun Province. However, no specimens were collected at the time, and the species remain unidentified. During fieldwork conducted at the same locations in April 2025, and at Ao Talo Wao, Ko Tarutao, Tarutao National Park, Mueang Satun District, Satun Province in May 2025, we collected specimens bearing both vegetative and reproductive parts. These specimens match the type and descriptions found in various taxonomic references of Rhizophorastylosa Griff. (e.g., Griffith 1854; Hou 1958; Backer and Bakhuizen van den Brink 1963; Duke and Bunt 1979), a species previously recorded from India, China to Indo-China, and the Pacific, but not Thailand. Consequently, this represents a new record for Thailand. A detailed morphological description, illustrations, lectotypification, and an updated identification key to the species of Rhizophora in Thailand are provided, along with notes on distribution, habitat and ecology, phenology, preliminary conservation assessment, etymology, vernacular name and specimens examined. Additionally, leaf anatomical characteristics and pollen morphology are presented.
Materials and methods
Plant specimens of Rhizophorastylosa were observed and collected in the peninsular region of Thailand. The collected specimens were identified by consulting taxonomic literature (e.g., Griffith 1854; Henslow 1878; Ridley 1922; Hou 1958, 1960; Backer and Bakhuizen van den Brink 1963; Tomlinson 1978; Duke and Bunt 1979; McCusker 1984; Qin and Boufford 2007; Duke 2010; Ng and Chan 2012; Setyawan et al. 2014; Ragavan 2015; Ragavan et al. 2015, 2021; Kannan et al. 2021) and by comparison with herbarium specimens housed in the following herbaria: BK, BKF, as well as those available through the virtual herbarium databases of A, BR, CAL, E, K, L, P, SING, US, and GBIF (https://www.gbif.org/). All herbarium codes follow Thiers (2025, continuously updated). The taxonomic history of the species was compiled from both published literature and online databases (IPNI 2025; POWO 2025; WFO 2025). Morphological characters, distribution, habitat and ecology, phenology, and uses were described based on newly collected herbarium specimens and the author’s field observations. The distribution map was generated using the R program, based on data from the specimen examined and credible literature sources (e.g., Ragavan 2015; Kannan et al. 2021; Ragavan et al. 2021). Conservation status was assessed according to the IUCN Red List Categories and Criteria (IUCN Standards and Petitions Committee 2024), supplemented with GeoCAT analysis (Bachman et al. 2011) and field data.
The leaf anatomical features of Rhizophorastylosa were investigated through transverse sectioning using a sliding microtome at thickness of 15–20 µm. To study the epidermal cells, the leaves were peeled and mounted. Permanent slides were prepared according to the standard methods described by Johansen (1940) and Kermanee (2008). Anatomical characteristics were observed and recorded photographically using an Olympus BX53 microscope equipped with an Olympus DP74 digital camera at the Department of Botany, Faculty of Science, Kasetsart University (KU). Anatomical terminology follows the conventions outlined by Metcalfe and Chalk (1957).
Samples of pollen grains were examined and recorded photographically using an Olympus BX53 microscope equipped with an Olympus DP74 digital camera. For scanning electron microscopy (SEM), materials were prepared at the Scientific Equipment Centre, Faculty of Science, Kasetsart University, by mounting the pollen grains on stubs with double-sided sellotape, sputter-coating them with gold, and examining them using an FEI Quanta 450 SEM (Hillsboro, OR, USA) at 15.00 kV. The characteristics of the pollen grains were examined and measured following the methods of Erdtman (1945, 1952) and Simpson (2010). Pollen morphology terminology follows that of Punt et al. (2007).
Results and discussion
Taxonomic treatment
Rhizophora
stylosa
Taxon classificationPlantaeMalpighialesRhizophoraceae
Griff., Not. Pl. Asiat. 4: 665. 1854; G. Hensl. in Hook. f., Fl. Brit. India 2: 436. 1878; Ridl., Fl. Malay Penins. 1: 693. 1922; Ding Hou, Fl. Males., Ser. 1, Spermat. 5(4): 456, figs. 5, 8a–f, 13. 1958 et Blumea 10(2): 629. 1960; Backer & Bakh. f., Fl. Java 1: 380. 1963; Toml., J. Arnold Arbor. 59: 163, figs. 3, 4D–F. 1978; N. C. Duke & J. S. Bunt, Aust. J. Bot. 27: 672. fig. 6a–b. 1979; A. McCusker in A. S. George et al., Fl. Australia 22: 2. 1984; W. Giesen et al., Mangrove Guideb. S.E. Asia: 239, fig. 239. 2007; H. Qin & Boufford, Fl. China 13: 296, fig. 316(1–9). 2007; N. C. Duke, Blumea 55: 179, fig. 8. 2010; A. D. Setyawan et al., Nusantara Bioscience 6(1): 77, figs. 6A.b–G.b, 8. 2014; Ragavan, Taxon. Mangroves Andaman and Nicobar Isl. PhD Thesis: 115, fig. 43. 2015; Ragavan et al., Indian Mangroves: 122–123. 2021; S. Kannan et al., Bot. Mar. 64(3): 7–8, figs. 3D, 5B, D, F, G, J, L, M (right), O, 2021.
BF9B24D7-9F6E-58BC-B854-545544BDE3D6
≡ RhizophoramucronataPoir.var.stylosa (Griff.) A. Schimp., Bot. Mitt. Tropen 3: 92. 1891; Guillaumin in Lecomte et al., Fl. Indo-Chine 2(6): 724. 1920.
Type.
Peninsular Malaysia • Malacca (Melaka, Malay), s.d., W. Griffth s.n. (lectotype K digital image! [K005752944] (Fig. 5A), designated here; isolectotype: BR digital image! [BR0000031831338] (Fig. 5B).
Description.
Habit small evergreen trees, single- to multi-stemmed, 2.5–8 m tall, 10–60 cm GBH. Stilt roots much-branched, descending from the base of the stem, bearing numerous lenticels. Branches and branchlets decussate, growing upward at acute angles; branches often with pendulous aerial roots; branchlets terete, glabrous, with conspicuous annular stipular scars and leaf scars at the nodes. Bark grayish brown or dark brown, varies from smooth, rough or shallowly longitudinally and transversely fissured. Stipules interpetiolar, in pairs enclosing the young shoot (including the terminal bud and young leaves) and young inflorescences, pale green or green, turning whitish pale green or creamish white before the leaves emerging or before falling off, reddish brown when dry, linear-lanceolate, gradually narrowing towards the apex, 3.5–7.5 cm long, 2.5–9 mm diam. at the base, apex acute, glabrous and caducous, outline in transverse section depressed orbicular basally and suborbicular to orbicular apically, with dense colleters aggregated in a basal band, producing a sticky white exudate; colleters sessile, pale yellow, narrowly conical, 0.5–1.2 mm long, 0.2–0.5 mm diam. at the base, apex obtuse. Leaves decussate and crowded at the apical part of branchlets, elliptic, 8–16 × 3–8.5 cm, apex mucronate, a short terminal stiff point pale green, turning black before they come off, 2–6 mm long, base cuneate, margin entire, coriaceous, shiny dark green above, pale green below, glabrous on both surfaces, with numerous conspicuous, scattered tiny black cork warts below, midrib pale green (paler than lamina), flattened above, raised below, secondary veins 8–14 on each side, curving towards the margin and connected in distinct loops and united into an intramarginal vein, visible above, obscure below, with intersecondary veins, veinlets reticulate, visible above, obscure below; petioles green, 2–4.8 cm long, 2–4.5 mm diam., glabrous; fresh leaves brittle when crushed; young leaves shiny pale green; mature leaves turning greenish bright yellow and bright yellow before falling off; dry leaves yellowish brown. Inflorescences axillary, opposite, compound dichasia, dichotomously branched, 4–13-flowered cymes, 5–12.5 cm long; peduncles pale green, 2.3–6 cm long, 2–3.5 mm diam., glabrous. Bracts 2, pale green, concave, 2.5–4 × 2.7–3.5 mm, apex rounded. Bracteoles at the base of the flower, pale green, connate at the base, bilobed, 3–5 × 4–6 mm. Flowers 4-merous; flower buds pale green, turning pale yellow when mature, ovoid or conical-ovoid, 0.9–1.5 cm long, 4–8 mm diam., apex obtuse; fully open flowers 1.2–1.8 cm diam.; sepals 4, erect or patent after anthesis, pale yellow on both sides, triangular, 0.7–1.3 cm × 4–6 mm, apex acute, coriaceous, glabrous; petals 4, creamish white, narrowly elliptic or elliptic, 0.6–1 cm × 1.5–3.5 mm, thin, involute, densely long-villous along margins; stamens 8, 4 antesepalous and 4 antepetalous (each petal enclosing 1 stamen in flower buds); anthers falcate-like, 3.5–8.5 × 1.2–2.8 mm, apex mucronate, triangular in outline in transverse section; filaments very short; free part of the ovary 0.7–1.2 mm long, 1.2–1.8 mm diam.; style 3–5 mm long; stigma 2-lobed; pedicels pale green, 0.6–1 cm long, 2–3 mm diam. Fruits brownish green, greenish brown or brown, ovoid or conical-ovoid, 2–2.7 cm long, 1–1.7 cm diam. at the base, apex obtuse (before seed germination); obpyriform, 2.2–4 cm long, 1.7–3 cm diam. at the basal part, 0.9–1.7 cm diam. at the apical part (when the hypocotyls nearly come off), roughened surface; persistent sepals reflexed, triangular, 0.9–1.1 cm × 5–8.5 mm; infructescence stalks 2–6 cm long, 2–3 mm diam.; fruit stalks 0.6–1.7 cm long, 3–5 mm diam. Seeds 1, viviparous. Hypocotyls green, slightly glossy, cylindrical-clavate, 19.5–40 cm long, 6.5–9 mm diam. at the apical part, 1–1.6 cm diam. at the widest part, 0.8–1.3 cm diam. at the basal part, acute at the basal end, roughened surface, with numerous, scattered lenticels; cotyledonous cylindrical tubes pale green or greenish pale yellow, 0.6–1.2 cm diam. (can be seen when the hypocotyls nearly falling off).
Rhizophorastylosa. A, B. Flowering branchlets showing a terminal interpetiolar stipule, leaves, and inflorescences with flower buds and fully open flowers; C. Flower buds; D. Flower bud in transverse section; E. Fully open flower in lateral view; F. Fully open flower in top view; G. Flower bud showing stamens with sepals and petals removed; H. Flower bud in longitudinal section; I. Sepals; J. Petals with densely long-villous along margins; K. Stamen; L–M. Pistils showing a long style; N–O. Fruits showing successive stages in development of hypocotyl. Drawn by Wanwisa Bhuchaisri.
Rhizophorastylosa. A–D. Habitats and habit; E. Stems supported by numerous branched stilt roots; F. Stem, outer bark and inner bark; G. Leaves showing adaxial and abaxial surfaces: Population A (left) and Population B (right); H. Branchlets, leaves and young inflorescences; I. Leaf scar with many tiny vascular bundle strands, arranged in a crescent moon; J. Stipule with dense colleters aggregated in a basal band, producing a sticky white exudate. Photos: Chatchai Ngernsaengsaruay (A–C, E–J) and Rachanee Pothitan (D).
Rhizophorastylosa. A, B. Flowering branchlets showing inflorescences with flower buds and fully open flowers; C. Flowering branchlets showing flower buds and fully open flowers and fruiting branchlet showing young fruits; D. Inflorescence with flower buds and fully open flower; E, G. Flowering and fruiting branchlets showing leaves, inflorescences with flower buds and fully open flowers, and hypocotyls. F. Seedling. Photos: Chatchai Ngernsaengsaruay.
Rhizophorastylosa. A. Fully open flower; B. Flower bud in longitudinal section; C. Flower bud in transverse section; D. Dissected flower showing sepals, petals, stamens, and pistil; E–G. Sepals; H–J. Petals with densely long-villous along margins; K–N. Stamens; O. Pistil showing a long style. Photos: Pichet Chanton (A–N), Apinya Sukantatul (O).
Lectotype and isolectotype of Rhizophorastylosa from Malacca, Peninsular Malaysia. A. Lectotype, W. Griffth s.n. (K [K005752944]); B. Isolectotype, W. Griffth s.n. (BR [BR0000031831338]). Photos: © The Board of Trustees of the RBG, Kew, https://records.data.kew.org/occurrences/f573f797-57cc-48c7-b444-c6a9b8f085c3 (A), Meise Botanic Garden, Meise, Belgium, https://www.botanicalcollections.be/specimen/BR0000031831338 (B).
The measurements of the vegetative and reproductive parts of Rhizophorastylosa in Thailand are presented in Table 1.
Distribution.
India (Andaman and Nicobar Islands), China (South Guangxi, South Guangdong, Hainan), Taiwan (Taipei), Japan (Ryukyu Islands, also known as Nansei-shoto), Vietnam, Cambodia, Peninsular Thailand, Peninsular Malaysia (also called Malaya), Singapore, Indonesia [Java, Lesser Sunda Islands, Sulawesi (also called Celebes), Moluccas (also called Maluku)], Philippines, Australia (Northern Territory, Queensland, New South Wales, Western Australia), Melanesia (Bismarck Archipelago, New Guinea, Solomon Islands, Vanuatu, Fiji, New Caledonia), Micronesia (Mariana Islands, Caroline Islands, Gilbert Islands), Polynesia (Tuvalu, Tonga, Society Islands) (Fig. 6).
Distribution of Rhizophorastylosa. It is widely distributed from India (Andaman and Nicobar Islands), China to Indo-China, and the Pacific Islands. In Thailand, this species is known only from Ko Lidi and Ko Bulon Le within Mu Ko Phetra National Park, located in La-ngu District, and from Ao Talo Wao, Ko Tarutao, in Tarutao National Park, Mueang Satun District, Satun Province. Map: Pichet Chonton & Chatchai Ngernsaengsaruay.
Distribution in Thailand.
Rhizophorastylosa is known only from Ko Lidi and Ko Bulon Le within Mu Ko Phetra National Park, located in La-ngu District, and from Ao Talo Wao, Ko Tarutao, in Tarutao National Park, Mueang Satun District, Satun Province, Peninsular Thailand (Fig. 6).
Habitat and ecology.
Rhizophorastylosa typically occurs along coastal areas with muddy-sandy, sandy or sandy-rocky substrates. This species is found at the seaward edge of mangrove forests and is well adapted to grow in open coastal environments, particularly along fairly exposed shores.
The populations of R.stylosa observed in Thailand are also located in low intertidal zones characterized by rocky and sandy substrates, similar to the observations reported from the Andaman and Nicobar Islands, India by Ragavan (2015) and Ragavan et al. (2021).
In the Andaman and Nicobar Islands, the species is uncommon and is mostly found in mangroves fringing the open coast, towards the seaward side. It is often found in mid to low intertidal zones and along downstream tidal creeks. The species grows in a variety of habitats, including disturbed mangrove areas. One distinctive niche is its ability to grow along the edges of small coral islands, where it establishes on coral substrates (Ragavan 2015; Ragavan et al. 2021).
Phenology.
Flowering, fruiting and viviparous germination more than once, nearly throughout the year.
Conservation status.
Rhizophorastylosa is widely distributed, ranging from India (Andaman and Nicobar Islands) and China through Indo-China to the Pacific Islands. It is known from numerous localities, with a very large EOO of 52,723,897.41 km^2^ and a relatively large AOO of 1,908 km^2^. In Thailand, however, the species is restricted to Satun Province in the peninsular region, where it has a small EOO of 251.22 km^2^ and a relatively small AOO of 12 km^2^. Despite its limited national distribution, its broad global range, presence in numerous localities, and the absence of significant threats to its survival, we recommend a conservation assessment of Least Concern (LC), in agreement with Ellison et al. (2010).
Etymology.
The specific epithet of Rhizophora stylosa is a Latin word that refers to its characteristic long style.
Vernacular name.
Kongkang phetra (โกงกางเภตรา) (suggested here); Red mangrove, Small stilted mangrove, Spotted mangrove, Stilted mangrove (English).
Uses.
To date, there has been no record of its use in Thailand. On Japan’s Iriomote Islands, the traditional dyeing of cotton fabric using tannins extracted from the bark of Rhizophorastylosa (Yaeyama hirugi), a technique known as kusaki-zome, remains an important cottage industry. The dye from the outer bark is brownish, while that from the inner bark is reddish (Baba 2004; Baba et al. 2013). On Iriomote and Ishigaki Islands, seedlings of R.stylosa are sold in souvenir shops to tourists as ornamentals (Baba et al. 2013). This species is utilized for timber, firewood, and the production of charcoal in the Pacific Island countries (Giesen et al. 2007; Kainuma et al. 2015). In Australia, Aboriginal people use its wood to make boomerangs, spears, and ceremonial items (Giesen et al. 2007).
Lectotypification.
Rhizophorastylosa was named by Griffith (1854: 665), who cited only the type locality “Hab. Malacca, in littoribus limosis, Pulo Bissar”. However, he did not designate a holotype, nor did he provide a collector number or indicate the herbarium where the material was housed. We have located the specimen W. Griffth s.n. from Malacca at K [K005752944] and BR [BR0000031831338], which following Art. 9.6 of the ICN (Turland et al. 2018), must be considered syntypes. William Griffith (1810–1845) was a British colonial physician and botanist who made substantial contributions to the herbarium of the East India Company. His herbarium was originally deposited at LINN, but was later transferred to Kew (K), with significant duplicate specimens housed at BR and CAL (Stafleu and Cowan 1976). Accordingly, we designate the specimen at K [K005752944] as the lectotype, with the specimen at BR [BR0000031831338] being an isolectotype, in accordance with Arts. 9.3 and 9.12 of the ICN (Turland et al. 2018).
Notes.
The colors and sizes of the stipules of Rhizophorastylosa are pale green or green, turning whitish pale green or creamish white before falling off. They are 3.5–7.5 cm long, 2.5–9 mm diam. at the base. In contrast, the stipules of R.mucronata are pale green, reddish pale green, or pale greenish red when young, turning pale yellow or reddish pale yellow before falling off, and are larger, 4.5–12.5 cm in length and 0.5–1.7 cm in diameter at the base. The petioles, peduncles, rachises of inflorescences, and pedicels of R.stylosa are usually terete, whereas those of R.mucronata are slightly to somewhat flattened.
Morphologically, R.stylosa is closely related to R.mucronata (Fig. 7) in its inflorescences and flowers (including flower buds, pedicelled flowers, bracteoles, sepals, and stamens); in the length of the stiff, pointed leaf tips; in the shape and basal diameter of the fruits (when the hypocotyls nearly come off); in the shape and color of the hypocotyls; and in the colors of the cotyledonous cylindrical tubes and colleters. The differences between these two Thai species of Rhizophora are presented in Table 2.
Rhizophoramucronata. A. Branchlets, leaves and terminal interpetiolar stipule; B. Leaves showing adaxial and abaxial surfaces; C. Flowering branchlet showing inflorescences with flower buds and fully open flower; D. Petal with densely and shorter villous along margins; E. Flower bud in longitudinal section showing very short style; F. Fruiting branchlet showing leaves and mature fruit with persistent sepals; G, H. Fruiting branchlets showing leaves and successive stages in development of hypocotyl. Photos: Chatchai Ngernsaengsaruay (A–C, F–H), Pichet Chonton (D, E).
Two distinct populations of R.stylosa were found in Mu Ko Phetra National Park, La-ngu District, Satun Province, Peninsular Thailand. Population A is distributed along coastal areas characterized by muddy-sandy substrates, predominantly occupying the seaward edge of the mangrove forest at Ko Lidi, La-ngu Subdistrict. In contrast, Population B occurs along coastal areas with sandy to sandy-rocky substrates, particularly along relatively exposed shorelines at Ko Bulon Le, Pak Nam Subdistrict. Morphological comparisons between the two populations reveal that individuals in Population A (3–8 m) tend to be taller than those in Population B (2.5–5 m). Furthermore, vegetative and reproductive structures, including stipules, leaves, petioles, inflorescences, peduncles, and fruits, are generally larger in Population A. These morphological differences are likely influenced by environmental conditions, suggesting that substrate type and coastal exposure play significant roles in shaping the growth and reproductive traits of R.stylosa in this region (Table 3).
Additional specimens examined.
Thailand. Peninsular: • Satun [Ko Lidi, Mu Ko Phetra National Park, La-ngu Subdistrict, La-ngu District, fl., fr. & vivipary, 22 Apr 2025, C. Ngernsaengsaruay et al. Rs01-22042025 (BKF); • Ao Phangka, Ko Bulon Le, Mu Ko Phetra National Park, Pak Nam Subdistrict, La-ngu District, fl., fr. & vivipary, 22 Apr 2025, C. Ngernsaengsaruay et al. Rs02-22042025 (BKF); • ibid., fl., fr. & vivipary, 22 Apr 2025, C. Ngernsaengsaruay et al. Rs03-22042025 (BKF); Ao Talo Wao, Ko Tarutao, Tarutao National Park, Mueang Satun District, fl., 30 May 2025, N. Mianmit & N. Kaveethanathum personal observation with photos; Note. C. Ngernsaengsaruay et al. = C. Ngernsaengsaruay, N. Mianmit, R. Pothitan, V. Jintana, N. Jintana & S. Kamsanor].
A key to the species of Rhizophora in Thailand
(Modified from Ngernsaengsaruay et al. 2024)
**: **
Anatomical study
Leaf anatomy of Rhizophorastylosa in Thailand
The leaf anatomical structure of Rhizophorastylosa is composed of five principal tissue layers: adaxial epidermis, hypodermis, palisade mesophyll, spongy mesophyll, and abaxial epidermis. Leaf thickness ranges from 574.11–881.72 μm. A thick cuticular wax layer (cuticle) covers both epidermal surfaces, with the adaxial surface bearing a slightly thicker cuticle than the abaxial surface (Fig. 9).
Epidermal cells are polygonal, occasionally trapezoidal with straight anticlinal walls, and are arranged in a single layer on both adaxial and abaxial surfaces. The adaxial epidermis is slightly thicker than the abaxial epidermis (Fig. 8).
Leaf anatomy of Rhizophorastylosa. A. Adaxial epidermis (under LM); B, C. Abaxial epidermis (under LM); D. Abaxial epidermis (under SEM) (Cw = cork wart, EpC = epidermal cells, St = stoma, and SubC = subsidiary cells). Photos: Pichet Chanton & Chatchai Ngernsaengsaruay.
Leaf anatomy of Rhizophorastylosa. A–E. Transverse section (Abe = abaxial epidermis, Ade = adaxial epidermis, Cr = druse crystals, Ct = cuticle, Cw = cork wart, EpC = epidermal cells, Hp = hypodermis, P = phloem, Pa = parenchyma, Pl = palisade mesophyll, Sp = spongy mesophyll, St = stoma, Vb = vascular bundle, X = xylem). Photos: Pichet Chanton & Chatchai Ngernsaengsaruay.
The stomata of R.stylosa are confined to the abaxial leaf surface, indicating that the leaves are hypostomatic.
The stomata are of the sunken cyclocytic type, surrounded by 6–8 subsidiary cells arranged in a concentric ring around each stoma (Fig. 8). Stomatal size ranges from 18.31–30.48 × 9.56–15.21 μm, with a density of 77–153 stomata/mm^2^.
According to previous studies, stomatal density is categorized into three groups: low (<300 stomata/mm^2^), medium (300–500 stomata/mm^2^), and high (>500 stomata/mm^2^). Based on this classification, the stomatal density of R.stylosa is considered low. Cork warts are present on the abaxial surface, with a density ranging from 15–52 cork warts percm^2^ and a diameter of 41.57–94.58 μm. These structures consist of densely packed cells, morphologically distinct from other epidermal cells and easily distinguishable from stoma (Fig. 8). Rhizophorastylosa has the largest cork wart size among the three species in Thailand. Rhizophoramucronata exhibits cork wart sizes ranging from 39.86–82.36 μm, with an average of 61.76 ± 12.83 μm, while R.apiculata has the smallest cork wart sizes, ranging from 35.69–75.48 μm, with an average of 54.99 ± 10.64 μm. The data for R.apiculata and R.mucronata are based on Ngernsaengsaruay et al. (2024).
Rhizophorastylosa possesses bifacial leaves. The mesophyll is composed three main tissues: hypodermis, palisade parenchyma, and spongy parenchyma (Fig. 9). The hypodermis consists of relatively large cells arranged in multiple layers beneath the epidermis. These cells are often filled with tannins, which cause them to appear darkly stained. The adaxial surface exhibits a greater average hypodermal thickness compared to the abaxial surface.
Hypodermal cells on the adaxial side are typically subcircular, elliptic, oblong, or rounded polygonal in shape, while those on the abaxial side tend to be broadly elliptic, circular, or rounded polygonal. The average cell size of the hypodermis is also larger on the adaxial surface than on the abaxial surface (Fig. 9).
Druse crystals are present in the hypodermal cells on both the adaxial and abaxial surfaces. The diameter of the druse crystals ranges from 14.84–30.06 μm (Fig. 9).
The palisade parenchyma consists of one to two layers of tightly packed, elongated cells located directly beneath the hypodermis. The spongy parenchyma is composed of five to seven layers of loosely arranged, irregularly shaped cells situated below the palisade parenchyma and above the abaxial epidermis. This tissue exhibits prominent intercellular spaces, forming a characteristic net-like structure (Fig. 9).
Observations of the vascular bundle arrangement in the midrib of R.stylosa reveal a complex structure, divided into three distinct regions: abaxial, medullary (central), and adaxial. Each vascular bundle comprises phloem located on the outer side and xylem on the inner side. These vascular bundles are incompletely enclosed by a layer of sclerenchyma cells (Fig. 9).
A comparison of the leaf anatomical characteristics of R.stylosa in Thailand with findings from previous studies is presented in Table 4.
Palynological study
Pollen morphology of Rhizophorastylosa in Thailand
The pollen grains of Rhizophorastylosa are monads, isopolar, radially symmetrical, and tricolporate. They exhibit either a prolate spheroidal or oblate spheroidal shape, with a P/E ratio ranging from 0.88–1.15. In polar view (amb), the outline is subcircular or rounded triangular.
The pollen grains are small in size, with the polar axis measuring 17.93–22.36 μm and the equatorial axis 16.79–22.94 μm. The colpus length ranges from 12.92–20.40 μm, and the colpus width from 2.35–3.85 μm. The exine thickness ranges from 0.83–2.43 μm., and the exine sculpturing is reticulate (Fig. 10). Rhizophorastylosa has the largest pollen size among the three species in Thailand. Rhizophoramucronata exhibits pollen sizes along the polar axis ranging from 12.37–18.49 μm, with an average of 15.77 ± 1.58 μm, while R.apiculata has the smallest pollen size along the polar axis, ranging from 11.16–13.50 μm, with an average of 12.36 ± 0.78 μm. The data for R.apiculata and R.mucronata are based on Ngernsaengsaruay et al. (2024).
LM and SEM micrographs of pollen grains of Rhizophorastylosa. A. Tricolporate aperture and rounded-triangular shape in polar view; B. Colporate aperture in equatorial view; C. Tricolporate aperture and rounded-triangular shape in polar view; D. Colporate aperture in equatorial view; E. Exine sculpturing, under LM (A, B), under SEM (C–E). Photos: Pichet Chanton & Chatchai Ngernsaengsaruay.
A comparison of the pollen morphology of Rhizophorastylosa in Thailand with previous studies is presented in Table 5.
Supplementary Material
XML Treatment for Rhizophora stylosa
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baba S (2004) Keynote Presentation: What we can do for mangroves. In: Vannucci M (Ed.) Mangrove Management and Conservation: Present and Future.United Nations University Press, Tokyo, Japan, 8–36.
- 2Baba S Chan HT Aksornkoae S (2013) Useful Products from Mangrove and Other Coastal Plants. ISME Mangrove Educational Book Series No. 3.International Society for Mangrove Ecosystems (ISME), Okinawa, Japan, and International Tropical Timber Organization (ITTO), Yokohama, Japan, 98 pp.
- 3Bachman S Moat J Hill A Wde la Torre J Scott B (2011) Supporting red list threat assessments with Geo CAT: Geospatial conservation assessment tool. In: Smith V Penev L (Eds) e-Infrastructures for data publishing in biodiversity Science.Zoo Keys 150: 117–126. 10.3897/zookeys.150.2109 PMC 323443422207809 · doi ↗ · pubmed ↗
- 4Backer CA Bakhuizen van den Brink RC (1963) Rhizophoraceae. In: Flora of Java Vol 1. N. V. P. Noordhoff, Groningen, The Netherlands, 378–381.
- 5Duke NC (2010) Overlap of eastern and western mangroves in the south-western Pacific: Hybridization of all three Rhizophora (Rhizophoraceae) combinations in New Caledonia.Blumea 55(2): 171–188. 10.3767/000651910 X 527293 · doi ↗
- 6Duke NC Bunt JS (1979) The genus Rhizophora (Rhizophoraceae) in North-eastern Australia.Australian Journal of Botany 27(5): 657–678. 10.1071/BT 9790657 · doi ↗
- 7Ellison J Duke N Kathiresan K Salmo IIISG Fernando ES Peras JR Sukardjo S Miyagi T (2010) Rhizophorastylosa. The IUCN Red List of Threatened Species 2010. 10.2305/IUCN.UK.2010-2.RLTS.T 178850 A 7626520.en [Accessed on 24 April 2025] · doi ↗
- 8Erdtman G (1945) Pollen morphology and plant taxonomy V. on the occurrence of tetrads and dyads.Svensk Botanisk Tidskrift Utgifven af Svenska Botaniska Foreningen 39: 286–297.