Amphicercidus (Hemiptera, Aphidinae, Macrosiphini) species in China: two synonyms and two new records

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1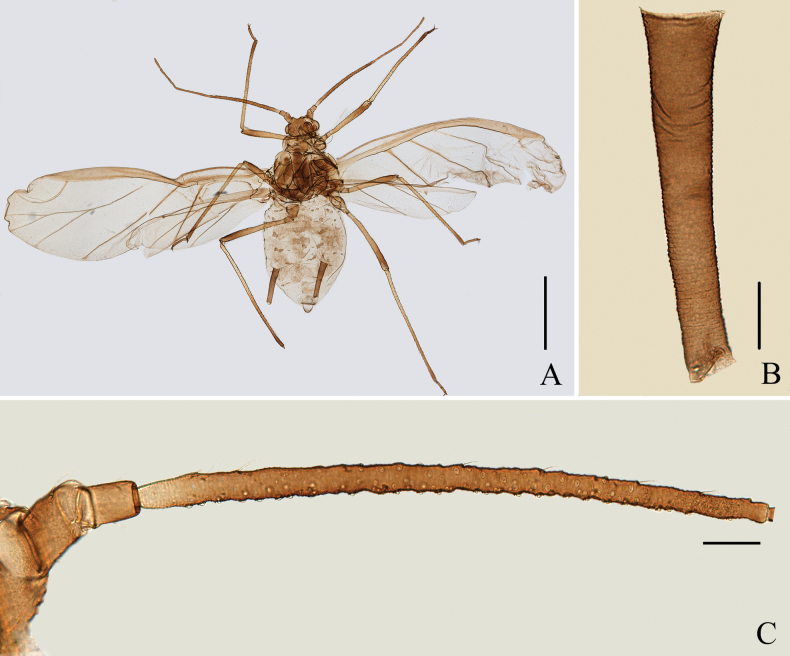 Figure 2
Figure 2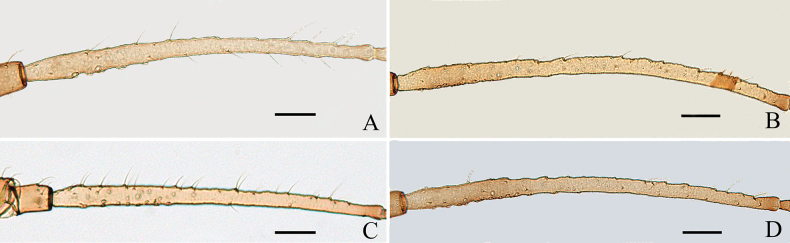 Figure 3
Figure 3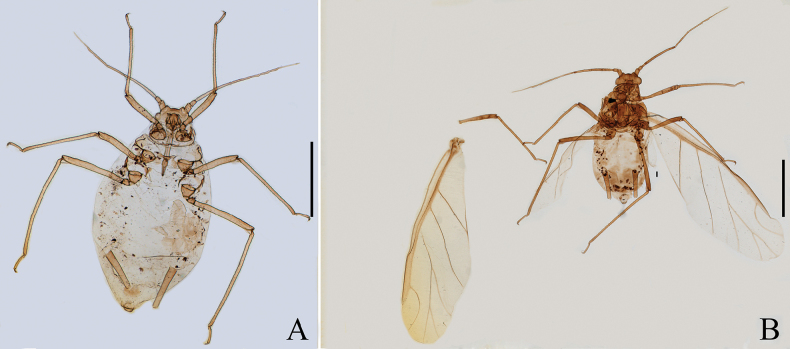 Figure 4
Figure 4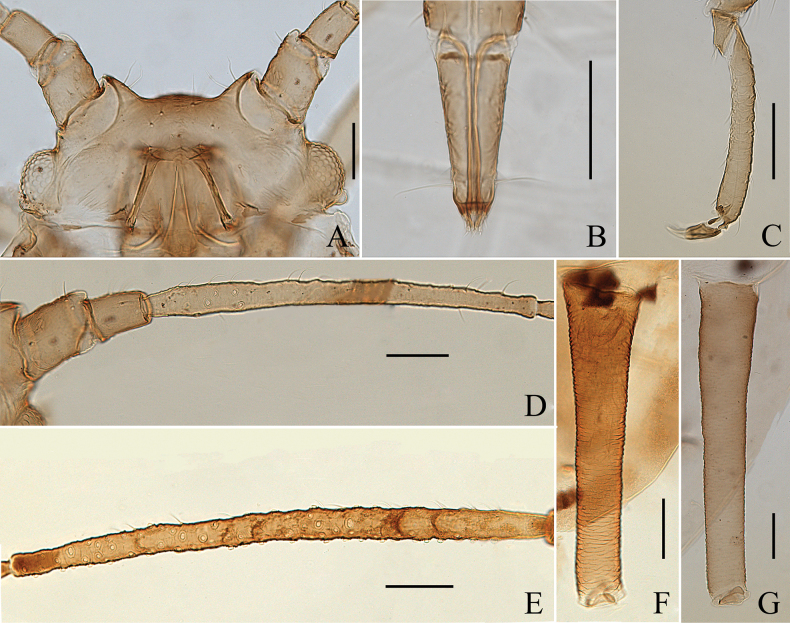 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7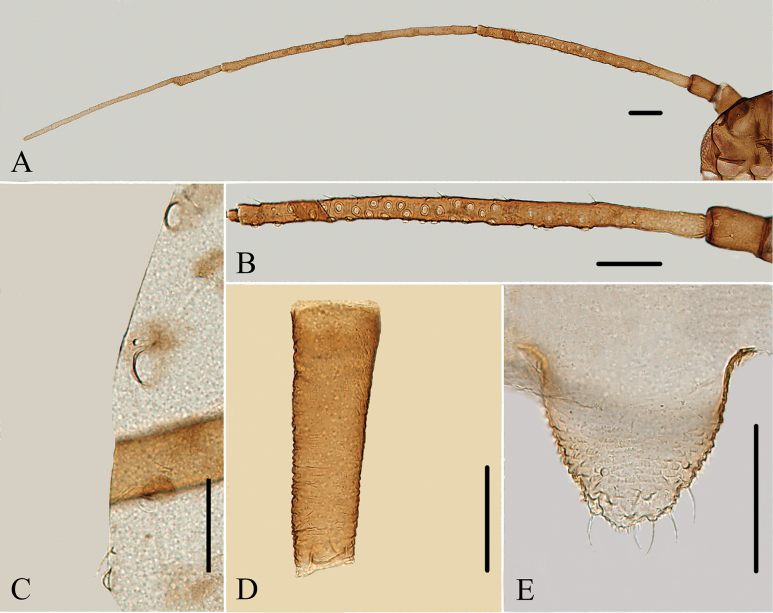 Figure 8
Figure 8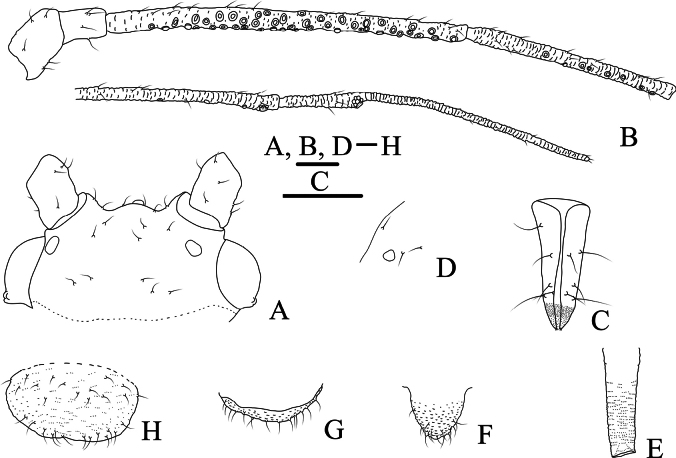 Figure 9
Figure 9 Figure 10
Figure 10 Figure 11
Figure 11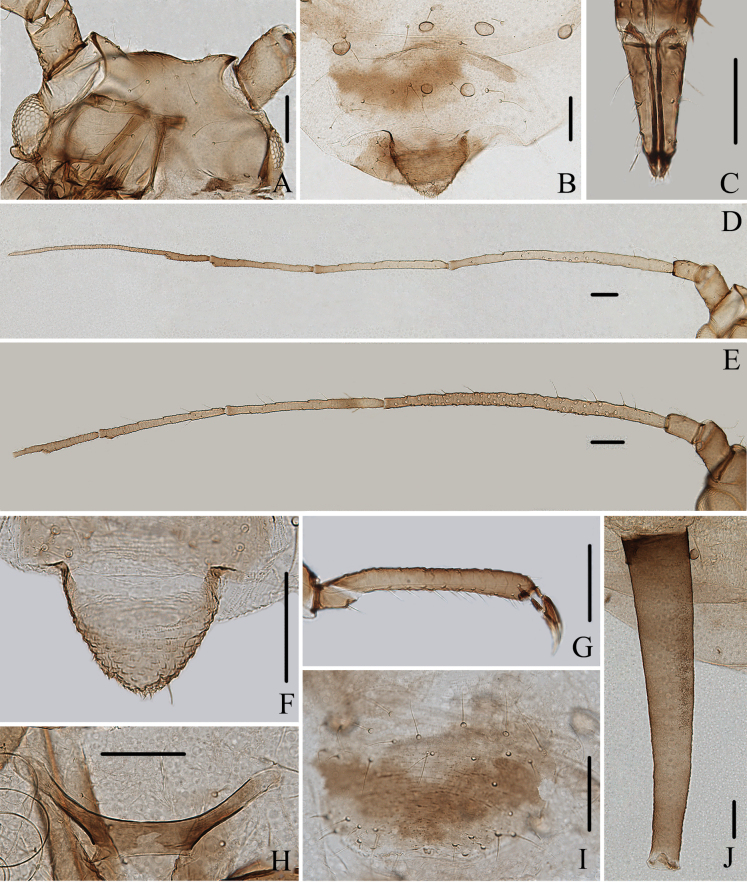 Figure 12
Figure 12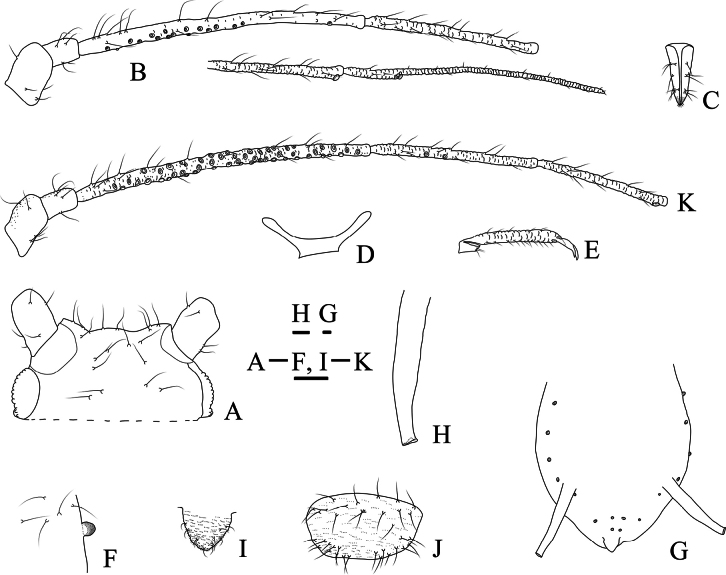 Figure 13
Figure 13 Figure 14
Figure 14| Species | Voucher number | Collection locality | Host plant | COI |
|---|---|---|---|---|
|
| 29606 | China: Xizang |
|
|
|
| 32720 | China: Xizang |
|
|
|
| 32721 | China: Xizang |
|
|
|
| 44055 | China: Beijing |
|
|
|
| 45445 | China: Beijing |
|
|
|
| 48119 | China: Beijing |
|
|
|
| 50282 | China: Yunnan |
|
|
|
| 030603J01 | Korea |
|
| Parts |
|
|
| ||||||
|---|---|---|---|---|---|---|---|---|---|
| Apterous viviparous females ( | Alate viviparous females ( | Apterous viviparous females ( | Alate viviparous females ( | Alate viviparous females ( | Alate viviparous females ( | Apterous viviparous females ( | Alate viviparous females ( | ||
| Length (mm) | Body length | 2.06–3.51 | 2.92–3.02 | 2.87–3.23 | 2.38–2.61 | 2.66–3.36 | 2.81–3.01 | 2.61–3.39 | 2.81–3.17 |
| Body width | 0.89–1.81 | 1.24–1.32 | 1.62–1.80 | 1.06–1.10 | 1.23–1.51 | 1.12–1.14 | 1.42–1.78 | 1.24–1.30 | |
| Antennae | 1.64–2.74 | – | 1.89–2.03 | 2.34–2.57 | – | 2.51–2.81 | 2.10–2.81 | – | |
| Ant. I | 0.11–0.15 | 0.15–0.15 | 0.13–0.13 | 0.12–0.14 | 0.12 | 0.14–0.17 | 0.13–0.18 | 0.14–0.15 | |
| Ant. II | 0.08–0.11 | 0.09–0.11 | 0.09–0.10 | 0.10 | 0.12 | 0.11–0.12 | 0.11–0.13 | 0.11 | |
| Ant. III | 0.44–1.07 | 0.98–1.11 | 0.59–0.65 | 0.81–0.88 | 1.18 | 0.72–0.83 | 0.57–0.84 | 0.86 | |
| Ant. IV | 0.26–0.54 | 0.50–0.51 | 0.31–0.36 | 0.40–0.43 | – | 0.44–0.50 | 0.35–0.50 | 0.44 | |
| Ant. V | 0.26–0.43 | 0.50–0.51 | 0.27–0.30 | 0.33–0.38 | – | 0.41–0.48 | 0.31–0.39 | 0.36 | |
| Ant. VIb | 0.14–0.16 | 0.13–0.13 | 0.15–0.15 | 0.16–0.17 | – | 0.17–0.19 | 0.15–0.17 | 0.16 | |
|
| 0.36–0.51 | – | 0.34–0.35 | 0.40–0.49 | – | 0.53–0.54 | 0.49–0.59 | – | |
|
| 0.15–0.18 | 0.16–0.17 | 0.16–0.17 | 0.14–0.16 | 0.17–0.18 | 0.15–0.17 | 0.15–0.17 | 0.15–0.16 | |
| Hind femur | 0.70–1.24 | 1.04–1.13 | 1.00–1.04 | 0.95–0.96 | 1.15–1.20 | 0.99–1.00 | 0.84–1.15 | 1.04 | |
| Hind tibia | 1.14–1.88 | 1.85–1.93 | 1.55–1.65 | 1.58–1.64 | 1.94–2.10 | 1.55–1.66 | 1.42–1.93 | 1.82 | |
|
| 0.23–0.28 | 0.24–0.27 | 0.26–0.26 | 0.25 | 0.29–0.30 | 0.25–0.27 | 0.24–0.28 | 0.26–0.27 | |
|
| 0.43–0.82 | 0.56–0.62 | 0.67–0.71 | 0.54–0.55 | 0.65–0.67 | 0.25–0.33 | 0.64–0.89 | 0.63 | |
|
| 0.12–0.16 | 0.09–0.12 | 0.14–0.15 | 0.12–0.13 | 0.16 | 0.08–0.09 | 0.14–0.18 | 0.10–0.11 | |
|
| 0.10–0.12 | 0.08–0.09 | 0.10 | 0.08–0.09 | 0.13 | 0.07–0.08 | 0.13–0.14 | 0.09 | |
|
| 0.08–0.09 | 0.08–0.09 | 0.09–0.10 | 0.08–0.09 | 0.11–0.12 | 0.06–0.07 | 0.09–0.09 | 0.09 | |
| Cauda | 0.09–0.14 | 0.13–0.14 | 0.12 | 0.09–0.13 | 0.10–0.13 | 0.13 | 0.13–0.16 | 0.16 | |
|
| 0.15–0.22 | 0.18 | 0.18 | 0.14–0.16 | 0.14–0.16 | 0.17 | 0.16–0.20 | 0.18 | |
|
| 0.03–0.04 | 0.04 | 0.03–0.04 | 0.03–0.04 | 0.05–0.06 | 0.03–0.04 | 0.03–0.04 | 0.03 | |
| Hind tibia MW | 0.04–0.06 | 0.04–0.05 | 0.06 | 0.04–0.10 | – | 0.05 | 0.05–0.06 | 0.05 | |
| Cephalic setae | 0.05–0.07 | 0.05 | 0.06 | 0.05 | 0.05 | 0.04 | 0.08 | 0.08 | |
| Setae on Tergite I | 0.04–0.07 | 0.04–0.06 | 0.05 | 0.03 | 0.05–0.08 | 0.04 | 0.06–0.08 | 0.07 | |
| Setae on Tergite VIII | 0.06–0.08 | 0.06–0.07 | 0.06 | 0.06–0.07 | – | 0.05 | 0.09 | 0.09 | |
| Setae on Ant. III | 0.03–0.06 | – | 0.05 | 0.04 | 0.04 | 0.03 | 0.06–0.09 | 0.07 | |
| Setae on Hind tibia | 0.06–0.09 | 0.07 | 0.08–0.09 | 0.05–0.07 | 0.07–0.10 | 0.07 | 0.07–0.10 | 0.07–0.09 | |
| Number | Number of secondary rhinaria on Ant. III | 3–39 | 101–103 | 4–6 | 90–112 | 120–123 | 43–46 | 3–17 | 58–59 |
| Number of secondary rhinaria on Ant. IV | 0 | 0 | 0 | 1–4 | 4 | 0–8 | 0 | 0 | |
| Number of secondary rhinaria on Ant. V | 0 | 0 | 0 | 0 | 0 | 0–1 | 0 | 0 | |
| Ratio (times) | Body length / Body width | 1.74–2.31 | 2.28–2.35 | 1.78–1.80 | 2.25–2.37 | 2.16–2.42 | 2.51–2.63 | 1.84–1.90 | 2.27–2.44 |
| Whole antennae / Body | 0.80–0.87 | – | 0.58–0.71 | 0.90–1.08 | – | 0.89–0.93 | 0.81–0.83 | – | |
| Hind femur / Ant. III | 1.18–1.69 | 0.94–1.15 | 1.54–1.75 | 1.08–1.19 | 0.97–1.01 | 1.2 | 1.37–1.48 | 1.20–1.22 | |
| Hind tibia / Body | 0.56–0.59 | 0.63–0.64 | 0.51–0.54 | 0.61–0.69 | 0.58–0.65 | 0.55 | 0.54–0.57 | 0.57–0.65 | |
| Ant. I / Ant. III | 0.15–0.24 | 0.13–0.15 | 0.21–0.22 | 0.14–0.17 | 0.10 | 0.19–0.20 | 0.21–0.23 | 0.16–0.17 | |
| Ant. II / Ant. III | 0.11–0.19 | 0.08–0.11 | 0.15–0.16 | 0.11–0.13 | 0.10 | 0.14 | 0.16–0.19 | 0.13 | |
| Ant. IV / Ant. III | 0.45–0.61 | 0.45–0.52 | 0.52–0.56 | 0.49–0.50 | – | 0.60 | 0.59–0.63 | 0.51 | |
| Ant. V / Ant. III | 0.42–0.59 | 0.34–0.40 | 0.45–0.46 | 0.41–0.44 | – | 0.57 | 0.47–0.55 | 0.42 | |
| Ant. VIb / Ant. III | 0.17–0.33 | 0.14 | 0.24–0.26 | 0.19–0.20 | – | 0.23 | 0.20–0.27 | 0.18 | |
| 0.53–0.81 | – | 0.54–0.58 | 0.49–0.56 | – | 0.65 | 0.70–0.86 | – | ||
| 2.29–3.28 | – | 2.25–2.26 | 2.44–2.90 | – | 2.87 | 3.21–3.43 | – | ||
| 2.50–2.68 | 2.45–3.00 | 2.23–2.69 | 2.30–2.59 | 1.57–1.66 | 2.34–2.62 | 2.22–2.38 | 2.03–2.19 | ||
| 0.61–0.66 | 0.62–0.68 | 0.62–0.64 | 0.55–0.63 | 0.57–0.62 | 0.58–0.62 | 0.61–0.63 | 0.57–0.60 | ||
| Cauda / | 0.58–0.69 | 0.73–0.77 | 0.67 | 0.63–0.80 | 0.68–0.83 | 0.78 | 0.78–0.80 | 0.86–0.89 | |
| Cephalic setae / | 1.36–2.14 | 1.28 | 1.59–1.97 | 1.37 | 0.90 | 1.22 | 2.11–2.70 | 2.52–2.55 | |
| Setae on Tergite I / | 1.25–2.10 | 1.13–1.57 | 1.30–1.30 | 0.94 | 0.92–1.45 | 1.16 | 2.03–2.18 | 2.12–2.15 | |
| Setae on Tergite VIII / | 2.05–2.62 | 1.44–2.03 | 1.57 | 1.88–1.94 | – | 1.13 | 2.32–2.90 | 2.73–2.82 | |
| Setae on ANT. III / ANT. III BD | 0.96–1.76 | – | 1.38–1.45 | 1.00–1.03 | 0.74–0.77 | 0.80 | 2.07–2.29 | 2.09–2.12 | |
| Setae on hind tibia / Hind tibia MW | 1.11–1.70 | 1.48–1.55 | 1.32–1.52 | 0.52–1.66 | 0.96–1.44 | 1.50 | 1.17–2.00 | 1.58–2.00 | |
| 0.21–0.24 | 0.19–0.20 | 0.22–0.23 | 0.21–0.23 | 0.19–0.21 | 0.09–0.11 | 0.25–0.26 | 0.20–0.22 | ||
| 5.22–6.21 | 4.03–4.78 | – | 4.30–6.30 | 5.17–6.21 | 1.97 | 5.13–5.45 | 3.97–3.99 | ||
| 0.73–0.98 | 0.51–0.63 | 1.04–1.20 | 0.62–0.68 | 0.54–0.57 | 0.35–0.39 | 1.06–1.13 | 0.73 | ||
| 3.49–6.23 | 4.59–6.71 | 4.49–4.94 | 4.18–4.59 | 3.70–4.28 | 3.02–3.48 | 4.45–5.08 | 5.85–6.04 | ||
| 4.44–7.91 | 6.78–7.23 | 6.61–7.12 | 5.83–6.85 | 4.35–5.29 | 3.79–4.19 | 4.58–7.11 | 7.14–7.20 | ||
| 5.20–10.04 | 6.56–7.43 | 6.88–8.38 | 5.96–6.85 | 4.98–5.55 | 4.62–5.03 | 7.37–9.99 | 6.75–7.03 | ||
| 1 | Abdominal tergites lacking distinct tubercles; antennal segment III without secondary rhinaria in apterae and with 2–5 secondary rhinaria in alatae; on |
|
| – | Abdominal tergites with distinct rounded tubercles; antennal segment III with at least 6 secondary rhinaria in apterae and alatae; on |
|
| 2 | Siphunculi as short cylinder, 0.09–0.11× body length, 2.00× cauda |
|
| – | Siphunculi as long and thick cylinder, at least 0.20× body length, at least 4.00× cauda |
|
| 3 | Abdominal tergites II-VI each with one pair of developed and round marginal tubercles and tergites VII-VIII each with one pair of spinal tubercles; processus terminalis 2.90–3.70× as long as the basal part of the last antennal segment; antennal segment III with 6–11 secondary rhinaria in apterae; on |
|
| – | Abdominal tergites II-VIII with irregularly arranged round tubercles; processus terminalis 1.60–2.80× as long as the basal part of the last antennal segment; antennal segment III with 3–39 secondary rhinaria in apterae; on |
|
| 53 | (Fund.) Dorsal and antennal hairs short and inconspicuous. | |
| – | Dorsal and antennal hairs long and fine-pointed. |
|
| 54 |
| |
| – |
| |
| 55 | ANT III with only 3–4 rhinaria (except in alatiform individuals). Cauda with 6 hairs |
|
| – | ANT III with 6–49 rhinaria. Cauda with 6–19 hairs |
|
| 1 | Siphunculi pore-shaped or short conic; ultimate rostral segment acute, stiletto-shaped |
|
| – | Siphunculi cylindrical; ultimate rostral segment wedge-shape |
|
| 2 | Cauda with semicircular basal part and pointed finger-shaped distal part, and the hind margin of anal plate indented to two conical tubercles in apterae |
|
| – | Cauda semicircular, helmet- or triangular-shaped, and anal plate semicircular in apterae |
|
| 3 | Antennal segment III with secondary rhinaria in apterae |
|
| Antennal segment III without secondary rhinaria in apterae |
| |
| 4 | Abdominal tergites without sclerotized markings; the second hind tarsal segment at least 1.50× ultimate rostral segment; siphunculi as stout cylinder, without flange |
|
| – | Abdominal tergites with sclerotized markings; the second hind tarsal segment shorter than ultimate rostral segment; siphunculi as long cylinder, with flange |
|
| 5 | Antennal segments III and IV with secondary rhinaria in alatae; cauda semicircular |
|
| – | Antennal segments III-V with secondary rhinaria in alatae; cauda escutcheon-shaped |
|
| 6 | Dorsal setae thick and long; dorsum covered with densely papillate tubercles; siphunculi as long cylinder, without flange |
|
| – | Dorsal setae short and pointed, or long and pointed; dorsum smooth or with small spicules; siphunculi as short cylinder, with flange |
|
| 7 | Siphunculi swollen at middle part, shorter than triangular cauda |
|
| – | Siphunculi cylindrical, not swollen, longer than cauda |
|
| 8 | Abdominal tergites I-V with paired and large rounded marginal and spinal tubercles; siphunculi cylinder |
|
| – | Dorsum without marginal tubercles or only with small marginal tubercles; siphunculi truncate conical or thick cylinder |
|
| 9 | Siphunculi truncate conical or cylindrical, with a subapical annular incision |
|
| – | Siphunculi cylindrical, without subapical annular incision |
|
| 10 | Ultimate rostral segment long and thin, wedge-shaped, > 2.00× of the second hind tarsal segment |
|
| – | Ultimate rostral segment wedge-shaped, less than 1.50× the second hind tarsal segment |
|
| 11 | Siphunculi long cylindrical, with long and pointed setae; first tarsal chaetotaxy: 4, 4, 2; abdominal tergites with a large central sclerite in alatae |
|
| – | Siphunculi short cylindrical, without setae; first tarsal chaetotaxy: 3, 3, 3; abdominal tergites with transverse bands in alatae |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhytoplasmas and Hemiptera pathogens · Orthoptera Research and Taxonomy · Hemiptera Insect Studies
Introduction
Amphicercidus was erected by Oestlund (1923) who specified Aphispulverulens Gillette, 1911 as the type species. The main morphological characteristics of the genus are: body heavily covered with wax powder in life, antennal segment III rather long and bearing numerous secondary rhinaria, the siphunculi are stout, cylindrical and without flange (Oestlund 1923). The characters of Amphicercidus are similar to those of Anuraphis, so some species of the genus were initially described in Anuraphis. Hori (1927) described Anuraphisjaponicus feeding on Lonicera in Japan, Takahashi (1927) described Anuraphislanigera feeding on Stauntonia in China (Taiwan), Gillette and Palmer (1929) described Anuraphisflocculosa feeding on Lonicera in USA. They have since been transferred to Amphicercidus (Gillette and Palmer 1932; Hori 1938; Eastop and Hille Ris Lambers 1976). Ferganaphisalticola Narzikulov & Mukhamediev, 1965 was described feeding on Lonicera in Tajikistan, and Ferganaphislonicericola Blackman & Eastop, 2024 feeding on Lonicera was also described in Central Asia. Later, the genus Ferganaphis was regarded as a synonym of Amphicercidus (Remaudière and Remaudière 1997). Subsequently, two species feeding on Lonicera, Amphicercidustuberculatus David, Narayanan & Rajasingh, 1971 and Amphicerciduslonicerae Maity & Chakrabarti, 1982, were found in India and a further two species feeding on Lonicera, Amphicercidussinilonicericola Zhang, 1980 and Amphicercidusforsythiae Zhang, Zhong & Zhang, 1992 were recorded in China. Until now, ten valid species of Amphicercidus have been recorded worldwide (Favret 2024).
A systematic revision of the genus in China has not been conducted, and the validity of A.lonicerae and A.sinilonicericola has been questioned (Blackman and Eastop 2024). Through a systematic investigation of Amphicercidus specimens in China, two species are here confirmed as synonyms and two are newly recorded.
Materials and methods
Aphid terminology used in this paper follows that of Miyazaki (1971) and Blackman and Eastop (2024), with body length measured from the frons to the end of the cauda. The specimens were examined using a Leica DM 2500 light microscope and photographed with a Leica MC 5400 camera. The unit of measurement is millimeter (mm) in Table 2. The following abbreviations are used in Table 2: Ant. I, II, III, IV, V, and VIb: antennal segments I, II, III, IV, V, and the base of antennal segment VI, respectively; PT: processus terminalis; Ant. III BD: basal diameter of antennal segment III; URS: ultimate rostral segment; BW URS: basal width of ultimate rostral segment; hind tibia MW: mid-width of hind tibia; 2HT: second hind tarsal segment; SIPH: siphunculus; DW SIPH: distal width of siphunculus; BW SIPH: basal width of siphunculus; MW SIPH: mid-width of siphunculus; BW Cauda: basal width of cauda; cephalic setae: the longest seta on vertex; setae on Ant. III: the longest seta on antennal segment III; setae on hind tibia: the longest seta on hind tibia; setae on tergite I: the longest marginal seta on abdominal tergite I; setae on tergite VIII: the longest spinal seta on abdominal tergite VIII.
DNA barcodes of COI were obtained from Chinese specimens and the voucher details are recorded (Table 1). Total genomic DNA was extracted from a single aphid nymph, preserved in 95% or 100% ethanol, using DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany). The standard DNA barcode gene of aphids was used and amplified with primers LepF and LepR (Foottit et al. 2008). The polymerase chain reaction (PCR) thermal regime was as follows: 5 min initial denaturation at 95 °C followed by 35 cycles of 95 °C for 30s, 52 °C for 30s, 72 °C for 1 min, and a 10 min final extension at 72 °C. The PCR products were sequenced in both directions with BigDye Terminator v. 3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA) and run on an ABI 3730 automated sequencer (Applied Biosystems). Sequences were assembled by SeqMan II (DNASTAR, Inc., Madison, WI, USA) with inspection and manual editing, and then were examined using BLAST to confirm the sequences were highly similar to other aphid sequences. All sequences were deposited in GenBank (Table 1). Multiple alignments were performed with Clustal W (Thompson et al. 1994) and then verified manually. Pairwise genetic distances for the COІ gene were estimated using MEGX (Kumar et al. 2018) under Kimura’s two-parameter (K2P) model (Kimura 1980).
The specimens examined are deposited in the National Animal Collection Resource Center, Institute of Zoology, Chinese Academy of Sciences, Beijing, China.
Taxonomic results
Amphicercidus
Taxon classificationAnimalia
Oestlund, 1923
2E37C9D0-5830-5412-B3DD-0DD7F4BE1F61
Amphicercidus Oestlund, 1923: 126.
Type species.
Aphispulverulens Gillette, 1911 by original designation.
Diagnosis.
Body elliptical and heavily covered with white wax powder in life; body dorsum smooth, abdominal tergites often with large and round marginal or spinal tubercles; setae of body dorsum and appendages long and pointed; head with undeveloped diverging antennal tubercles; median frontal tubercle flat or slightly swollen; antennal segment III with 3–49 secondary rhinaria in apterae and 2–152 secondary rhinaria in alatae; the second hind tarsal segment at least 1.50× of ultimate rostral segment; siphunculi as long stout cylinder, without flange, smooth or with fine transverse wrinkles; cauda broad, about as long as or slightly longer than its basal width.
The genus resembles Macchiatiella Del Guercio, 1909 in antennal segment III with secondary rhinaria and broad cauda, but it can be distinguished by the following characteristics: (1) antennal tubercles undeveloped (in Macchiatiella: antennal tubercles well developed); (2) body dorsum without sclerotized markings (in Macchiatiella: body dorsum with sclerotized markings); (3) second hind tarsal segment at least 1.50× of ultimate rostral segment (in Macchiatiella: second hind tarsal segment shorter than ultimate rostral segment); (4) siphunculi without flange (in Macchiatiella: siphunculi with flange). The genus also resembles Allocotaphis Börner, 1950 in having broad cauda, but it can be distinguished as follows: (1) abdominal tergites smooth, without sclerotized markings (in Allocotaphis: abdominal tergites often with sclerotized markings); (2) setae of body dorsum and appendages long and pointed (in Allocotaphis: setae of body dorsum and appendages short and pointed or capitate); (3) antennal segment III with numerous round secondary rhinaria in alatae (in Allocotaphis: antennal segment III with protuberant secondary rhinaria); (4) second hind tarsal segment at least 1.50× of ultimate rostral segment (in Allocotaphis: second hind tarsal segment shorter than ultimate rostral segment); (5) siphunculi without flange (in Allocotaphis: siphunculi with developed flange). The genus also similar to Anuraphis Del Guercio, 1907 in having broad cauda, but it can be distinguished as follows: (1) most species feeding on Lonicera (Caprifoliaceae) (in Anuraphis: the species transfer between Prunus (Rosaceae) and Asteraceae or Apiaceae); (2) body dorsum without sclerotized markings (in Anuraphis: body dorsum with sclerotized markings) (3) siphunculi long stout cylinder, smooth or with fine transverse wrinkles, without flange (in Anuraphis: siphunculi short cylinder and tapering, with close-set rows of minute fine spinules, with distinct flange); (4) second hind tarsal segment at least 1.50× of ultimate rostral segment (in Anuraphis: second hind tarsal segment shorter than ultimate rostral segment).
Biology.
Most species feed on the shoots and leaves of Lonicera (Caprifoliaceae), but Amphicercidusflocculosus (Gillette & Palmer, 1929) and Amphicerciduspulverulens (Gillette, 1911) feed on Symphoricarpus (Caprifoliaceae), and Amphicerciduslaniger (Takahashi, 1927) feeds on the young stems of Stauntonia (Lardizabalaceae) (Blackman and Eastop 2024).
Distribution.
China, Japan, Korea, India, Central Asia, Russia, USA, Canada, Mexico (Blackman and Eastop 2024).
Amphicercidus
japonicus
Taxon classificationAnimalia
(Hori, 1927)
F549A924-713E-5B37-9313-969668CE2511
Figs 1 , 2 , 3 , 4 , 5 , 6 , 14A–C ; Table 2
Anuraphis japonicus Hori, 1927: 193. Amphicercidus forsythiae Zhang, Zhong & Zhang, 1992: 196, syn. nov. Amphicercidus sinilonicericola Zhang, 1980: 53, syn. nov.
Types examined.
Holotype and paratypes of Amphicercidusforsythiae Zhang, Zhong & Zhang, 1992: two apterous females and two alate viviparous females, China: Yunnan (Lijiang City), 24.V.1980, No. 7150-1-1, on Lonicera, coll. T. S. Zhong and L. Y. Wang. Holotype and paratypes of Amphicercidussinilonicericola Zhang, 1980: two apterous females and three alate viviparous females, China: Yunnan (Kunming City), 24.II.1960, No. 4108, on Lonicera, coll. Y. F. Han.
Additional material examined.
Other specimens of Amphicercidusjaponicus (Hori, 1927): two apterous females, • China: Beijing (Baihua Mountain; alt. 1077 m), 3.VII.2018, No. 44055-1-1, on Lonicera, coll. H. Long; one apterous female (COI: PQ611221), • China: Beijing (Baihua Mountain), 3.VII.2018, No. 44055-1-1, on Lonicera, coll. H. Long; two apterous females, • China: Beijing (Baihua Mountain; alt. 731 m), 19.V.2019, No. 45445-1-1, on Lonicera, coll. H. Long; one apterous females (PQ611222), • China: Beijing (Baihua Mountain), 19.V.2019, No. 45445-1-1, on Lonicera, coll. H. Long; one alate viviparous female, • China: Beijing (Baihua Mountain: alt. 731 m), 19.V.2019, No. 45446-1-1, on Lonicera, coll. H. Long; one apterous female, • China: Yunnan (Lijiang City; alt. 2416.78 m), 26.V.2021, No. 50282-1-1. on Lonicera, coll. T. Y. Liu and S. Xu; one apterous female (COI: PQ611224), • China: Yunnan (Lijiang City), 26.V.2021, No. 50282-1-1. on Lonicera, coll. T. Y. Liu and S. Xu; one alate viviparous female and two apterous females, • China: Beijing (alt. 43.67 m), 11.V.2021, No. 48119-1-1, on Lonicera, coll. Y. Xu; one apterous female (COI: PQ611223), • China: Beijing, 11.V.2021, No. 48119-1-1, on Lonicera, coll. Y. Xu; one apterous female, • China: Xizang (Motuo county; 1757.5 m), 25.VI.2022, No. 51330-1-1, on Lonicera, coll. Z. X. Li; one nymph, • China: Sichuan (Aba City; alt. 3259.5 m); 22.VI.2021, No. 51598-1-1, on Lonicera, coll. T. Y. Liu and S. Xu.
Diagnosis.
Dorsal tubercles small and round, irregularly distributed on abdominal tergites II–V; antennal segment III with 3–39 secondary rhinaria in apterae and with more than 90 secondary rhinaria in alatae; processus terminalis 1.60–2.80× as long as the base of this segment (Figs 1–3).
Specimen No. 45445-1-1-2: apterous viviparous female of Amphicercidusjaponicus (Hori). A. Habitus of body; B. Ultimate rostral segment; C. Second hind tarsal segment; D. Siphunculus; E. Antennal segment III. Scale bars: 1.00 mm (A); 0.10 mm (B–E).
Specimen No. 45445-1-1-1: alate viviparous female of Amphicercidusjaponicus (Hori): A. Habitus of body; B. Siphunculus; C. Antennal segment III. Scale bars: 1.00 mm (A); 0.10 mm (B, C).
The secondary rhinaria on the antennal segment III of Amphicercidusjaponicus (Hori) in apterae shown: A. Antennal segment III with 5 secondary rhinaria in specimen No. 51330; B. Antennal segment III with 29 secondary rhinaria in specimen No. 48119; C. Antennal segment III with 20 secondary rhinaria in specimen No. 44055; D. Antennal segment III with 35 secondary rhinaria in specimen No. 45445. Scale bars: 0.10 mm.
Comment.
The validity of Amphicercidusforsythiae Zhang, Zhong & Zhang, 1992 and A.sinilonicericola Zhang, 1980 has been in doubt (Blackman and Eastop 2024). In the original descriptions, the primary differences between A.japonicus, A.forsythiae, and A.sinilonicericola are the number of secondary sensoria (Fig. 3), the length of setae on antennal segment III, the number of accessory hairs on the ultimate rostral segment, and the ratio of siphunculi length to head width across the eyes (Zhang et Zhong 1980; Zhang et al. 1992). However, after examining approximately twenty specimens collected in China, the number of accessory hairs on the ultimate rostral segment are 6–8, siphunculi are longer than head width, and other characteristics are overlapped. Therefore, the evidence is insufficient to support the validity of Amphicercidusforsythiae and A.sinilonicericola.
Amphicercidusforsythiae (Figs 4, 5) was distinguished from A.japonicus by the mesosternal furca with a long stem, antennal segment III with 4–6 circular secondary rhinaria in apterae and the longest setae on antennal segment III 0.79× of the basal width (Zhang et al. 1992). After examining the holotype and paratypes of A.forsythiae, these features are also present in A.japonicus and its morphological characteristics are extremely similar to those of A.japonicus. The only differences are following: A.forsythiae has slightly protuberant antennal tubercles (Fig. 5A) and siphunculi 1.04–1.20× of antennal segment III, while A.japonicus has undeveloped antennal tubercles and siphunculi 0.73–0.98× of antennal segment III. However, when the specimens are well-prepared, A.japonicus also has slightly protuberant antennal tubercles (Fig. 1A). The quantitative characteristics are a not sufficient to support the validity of A.forsythiae. Therefore, Amphicercidusforsythiae Zhang, Zhong & Zhang, 1992 is regarded as the junior synonym of Amphicercidusjaponicus (Hori, 1927).
Specimen No. 7150: Amphicercidusjaponicus (Hori) (Amphicercidusforsythiae Zhang, Zhong & Zhang, 1992 syn. nov.). A. Habitus of apterous viviparous female; B. Habitus of alate viviparous female. Scale bars: 1.00 mm.
Specimen No. 7150-1-1-2: Amphicercidusjaponicus (Hori) (Amphicercidusforsythiae Zhang, Zhong & Zhang, 1992 syn. nov.). Apterous viviparous female: A. Dorsal view of head; B. Ultimate rostral segment; C. Second hind tarsal segment; D. Antennal segments I-III; G. Siphunculus. Alate viviparous female; E. Antennal segment III; F. Siphunculus. Scale bars: 0.10 mm.
Amphicercidussinilonicericola (Fig. 6) was distinguished from A.japonicus by dorsal seta of head 0.93× basal width of antennal segment III, antennal segment III with 113–163 circular secondary rhinaria in alate, ultimate rostral segment with four secondary setae, mesosternal furca with a long stem, and siphunculi longer than head width across the eyes (Zhang et Zhong 1980; Blackman and Eastop 2024). After examining the holotype and paratypes of A.sinilonicericola Zhang, 1980 (Fig. 6), its morphological characteristics are extremely similar to those of A.japonicus, with differences only in the following characteristics: ultimate rostral segment wide wedge-shaped, 1.57–1.66× its basal width in alatae (in A.japonicus: ultimate rostral segment wedge-shaped, 2.45–3.00× its basal width in alatae); antennal segment III with 120–123 secondary rhinaria in alatae (Fig. 6B) (in A.japonicus: antennal segment III with 101–103 secondary rhinaria (Fig. 2C) in alatae). Considering that these quantitative characters can be affected by the specimen preparation process, it is impossible to accurately classify them as two species based on these characters. So, Amphicercidussinilonicericola Zhang, 1980 is regarded as a junior synonym of Amphicercidusjaponicus (Hori, 1927).
Specimen No. 4108-1-1-1: Amphicercidusjaponicus (Hori) (Amphicercidussinilonicericola Zhang, 1980 syn. nov.). Alate viviparous female: A. Habitus of body; B. Antennal segment III; C. Siphunculus. Apterous viviparous female; D. Antennal segment III; E. Siphunculus. Scale bars: 1.00 mm (A); 0.10 mm (B–E).
Biology.
The species feed on the shoots and leaves of Lonicera (Caprifoliaceae) (Hori 1927; Zhang and Zhong 1980; Zhang et al. 1992; Blackman and Eastop 2024).
Distribution.
China (Beijing, Liaoning, Shaanxi, Sichuan, Xizang, Yunnan); Japan; Korea; India; Russia.
Amphicercidus
laniger
Taxon classificationAnimalia
(Takahashi, 1927)
32B234CE-F833-5BE0-9881-D20C3381D0ED
Anuraphis lanigera Takahashi, 1927: 7.
Diagnosis.
Abdominal tergites lacking distinct tubercles; antennal segment III without secondary rhinaria in apterae and with 2–5 secondary rhinaria in alatae; processus terminalis 2.20× of the basal part of last antennal segment; feed on Stauntonia (Lardizabalaceae) (Takahashi 1927).
Biology.
The species feeds on the young stems of Stauntonia (Lardizabalaceae) (Takahashi 1927).
Distribution.
China (Taiwan).
New records for China
Amphicercidus
pulverulens
Taxon classificationAnimalia
(Gillette, 1911)
ABB8DA85-E26B-536A-A2D6-9C50564459EF
Figs 7 , 8 , 9 , 14D , Table 2
Aphis pulverulens Gillette, 1911: 324.
Specimens examined.
One alate viviparous female, China: Xizang (Linzhi City; alt. 2000 m), 4.VIII.2014, No. 29606-1-1, on Lonicera, coll. J. Chen and X. C. Zhu; one alate viviparous female (COI: PQ611218), China: Xizang (Linzhi City), 4.VIII.2014, No. 29606-1-1, on Lonicera, coll. J. Chen and X. C. Zhu; one alate viviparous female, China: Xizang (Linzhi City; alt. 3900 m), 5.VIII.2014, No. 29578-1-1, on Lonicera, coll. J. Chen and X. C. Zhu.
Specimen No. 25978-1-1: Amphicerciduspulverulens (Gillette): habitus of alate viviparous female. Scale bar: 1.00 mm.
Diagnosis.
Pronotum and abdominal tergites I-V each with one pair of marginal tubercles (Fig. 8C); siphunculi short stout cylinder, as long as second hind tarsal segment (Figs 8D, 9E), 0.09–0.11× body length; antennal segment III with 43–46 secondary rhinaria in alatae (Figs 8B, 9B); processus terminalis 2.87–3.14× the base of the last antennal segment.
Specimen No. 25978-1-1: Amphicerciduspulverulens (Gillette). Alate viviparous female: A. Antenna; B. Antennal segment III C. Marginal tubercles on abdominal tergites II-IV; D. Siphunculus; E. Cauda. Scale bars: 0.10 mm.
Comment.
The species has the shortest siphunculi in the genus and occurs on Symphoricarpos (Caprifoliaceae) in USA and Canada. After checking the syntype USNMENT00399393.001 of A.pulverulens from the USA (Smithsonian National Museum of Natural History, 2025), it is a new record for Chinese aphid fauna. This is the first record of A.pulverulens on Lonicera (Caprifoliaceae) in China. The species population found in China is very similar to the original description, with siphunculi being short and stout cylinder, as long as second hind tarsal segment, without flange; the Ant. III as long as Ant. III+IV; cauda short and broad. But, the characteristics of this species exhibit some differences between those found in China and the original records. In China, the species is orange in life, with straight siphunculi, whereas in USA, it is dusty green in life, and the siphunculi are weakly curved (Gillette 1911; Oestlund 1923).
Specimen No. 25978: Amphicerciduspulverulens (Gillette). Alate viviparous female: A. Dorsal view of head; B. Antenna; C. Ultimate rostral segment; D. Marginal tubercle on abdominal tergite II; E. Siphunculus; F. Cauda; G. Anal plate; H. Genital plate. Scale bars: 0.10 mm.
Biology.
The species feeds on the young stems of Symphoricarpos (Caprifoliaceae) in USA and Canada (Gillette 1911; Blackman and Eastop 2024), but this is the first record of it feeding on Lonicera (Caprifoliaceae) in China.
Distribution.
China (Xizang); USA; Canada.
Amphicercidus
tuberculatus
Taxon classificationAnimalia
David, Narayanan & Rajasingh, 1971
9DC130B2-44E4-5F14-B0CC-877BECB3429D
Figs 10 , 11 , 12 , 13 , 14E, F , Table 2
Amphicercidus tuberculatus David, Narayanan & Rajasingh, 1971: 419.
Specimens examined.
One alate viviparous female and one apterous female, 23.VII.2014, China: Xizang (Shigatse City; alt. 2657 m), No. 32720-1-1, on Lonicera, coll. J. Chen and X. C. Zhu; one apterous female (COI: PQ611219), 23.VII.2014, China: Xizang (Shigatse City), No. 32720-1-1, on Lonicera, coll. J. Chen and X. C. Zhu; one alate viviparous female and one apterous female, 23.VII.2014, China: Xizang (Shigatse City; alt. 2657 m), No. 32721-1-1, on Lonicera, coll. J. Chen and X. C. Zhu; one apterous female (COI: PQ611220), 23.VII.2014, China: Xizang (Shigatse City), No. 32721-1-1, on Lonicera, coll. J. Chen and X. C. Zhu.
Specimen No. 32721-1-1-2: Amphicercidustuberculatus David, Narayanan & Rajasingh: habitus of apterous viviparous female. Scale bar = 1.00 mm.
Specimen No. 32721-1-1-1: Amphicercidustuberculatus David, Narayanan & Rajasingh: habitus of alate viviparous female. Scale bar = 1.00 mm.
Diagnosis.
Dorsal tubercles on body are large and round; pronotum and abdominal tergites II–VII each with one pair of marginal tubercles, and abdominal tergites VII and VIII each with one pair of spinal tubercles (Figs 12B, 13G); antennal segment III with 3–17 secondary rhinaria in apterae (Figs 12D, 13B); processus terminalis 3.21–3.43× as long as the basal part of the last antennal segment.
Specimen No. 32721-1-1-2: Amphicercidustuberculatus David, Narayanan & Rajasingh. Apterous viviparous female: A. Dorsal view of head; B. Spinal tubercles of abdominal tergites VII-VIII C. Ultimate rostral segment; D. Antenna; F. Cauda; G. Second hind tarsal segment; H. Mesosternal furca siphunculus; I. Genital plate; J. Siphunculus. Alate viviparous female; E. Antennal segments I-VIb. Scale bars: 0.10 mm.
Specimen No. 32721-1-1-2: Amphicercidustuberculatus David, Narayanan & Rajasingh. Apterous viviparous female: A. Dorsal view of head; B. Antenna; C. Ultimate rostral segment; D. Mesosternal furca; E. Second hind tarsal segment; F. Marginal tubercle of abdominal tergite I; G. Marginal and spinal tubercles on abdomen; H. Siphunculus; I. Cauda; J. Genital plate. Alate viviparous female K. Antenna segments I-V. Scale bars: 0.10 mm.
Amphicercidus species in the field. A. Apterae of A.japonicus feeding on tender stem of Lonicera; B. Alatae of A.japonicus feeding on the underside of leaves of Lonicera; C. Apterae of A.japonicus feeding on tender stem of Lonicera; D. Alatae of A.pulverulens feeding on the underside of leaves of Lonicera; E. The apterae of A.tuberculatus feeding on tender stem of Lonicera; F. Apterae and alatae of A.tuberculatus feeding on tender stem of Lonicera.
Comment.
The species resembles Amphicercidusjaponicus (Hori), but the latter may be distinguished by having the following characters: body dorsum with small tubercles, irregularly distributed on the abdomen; antennal segment III with 16–39 secondary rhinaria in apterae; and the processus terminalis 2.41–3.11× as long as the basal part of the segment.
Biology.
The species feeds on feeds on the young stems and leaves of Lonicera (Caprifoliaceae) (David et al. 1971; Blackman and Eastop 2024).
Distribution.
China (Xizang); India.
DNA barcoding
The DNA barcodes of the three species of Amphicercidus (A.japonicus, A.pulverulens, and A.tuberculatus) from China were obtained, and the barcodes of A.pulverulens and A.tuberculatus were acquired for the first time. The final alignment sequences of COI consisted of 658 nucleotides, including 24 parsimony-informative sites. The interspecific genetic distances between A.japonicus and A.pulverulens is 2.80–3.00%, between A.japonicus and A.tuberculatus is 3.75–4.08%, and between A.pulverulens and A.tuberculatus is 4.56–4.57%. Based on DNA barcodes, the validity of these species is supported.
Key to species of Amphicercidus in China
**: **
Updated key couplets to aphids feeding on Lonicera in Blackman and Eastop, 2024
**: **
Discussion
The broad and rounded cauda is a generic distinctive morphological character in Macrosiphini, encompassing around thirty genera in the world (Shaposhnikov 1950). According to incomplete statistics, there are 12 genera with broad and rounded cauda belonging to the tribe Macrosiphini in China, which can be identified using the key below.
Key to Chinese Macrosiphini genera with broad cauda having a length shorter than its basal width
**: **
Supplementary Material
XML Treatment for Amphicercidus
XML Treatment for Amphicercidus japonicus
XML Treatment for Amphicercidus laniger
XML Treatment for Amphicercidus pulverulens
XML Treatment for Amphicercidus tuberculatus
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Blackman RL Eastop VF (2024) Aphids on the World’s Plants: An online identification and information guide. http://www.aphidsonworldsplants [accessed 20 Nov 2024]
- 2Börner C (1950) Neue europäische Blattlausarten.Privately published, Naumburg, 19 pp.
- 3David SK Narayanan K Rajasingh SG (1971) Five new species of aphids (Homotpera) from northwestern India.Oriental Insects 4(4): 431–425. 10.1080/00305316.1970.10433978 · doi ↗
- 4Del Guercio G (1909) Intorno a due nuovi generi e a tre specie nuove di afidi di California. Rivista di Patologia Vegetale. Serie 2 3: 328–332.
- 5Eastop VF Hille Ris Lambers D (1976) Survey of the World’s Aphids. Springer Dordrecht Publisher, 573 pp.
- 6Favret C (2024) Aphid Species File. http://Aphid.Species File [Accessed on: 2024-11-20]
- 7Foottit RG Maw HE Lvon Dohlen CD Hebert PDN (2008) Species identification of aphids (Insecta: Hemiptera: Aphididae) through DNA barcodes.Molecular Ecology Resources 8(6): 1189–1201. 10.1111/j.1755-0998.2008.02297.x 21586006 · doi ↗ · pubmed ↗
- 8Gillette CP (1911) Two Rhopalosiphum species and Aphispulverulens n. sp.Journal of Economic Entomology 4(3): 320–325. 10.1093/jee/4.3.320 · doi ↗