Cytosporalongdensis sp. nov. and C.sinensis (Diaporthales, Valsaceae) associated with Populusalba subsp. pyramidalis canker and dieback in China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
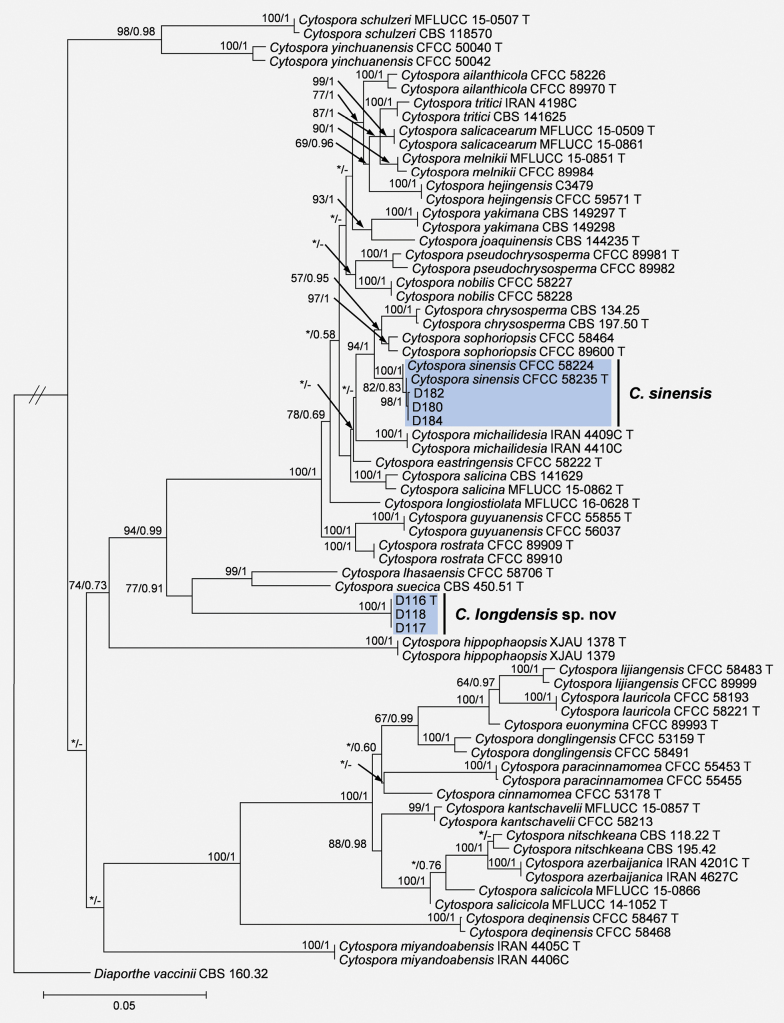 Figure 1
Figure 1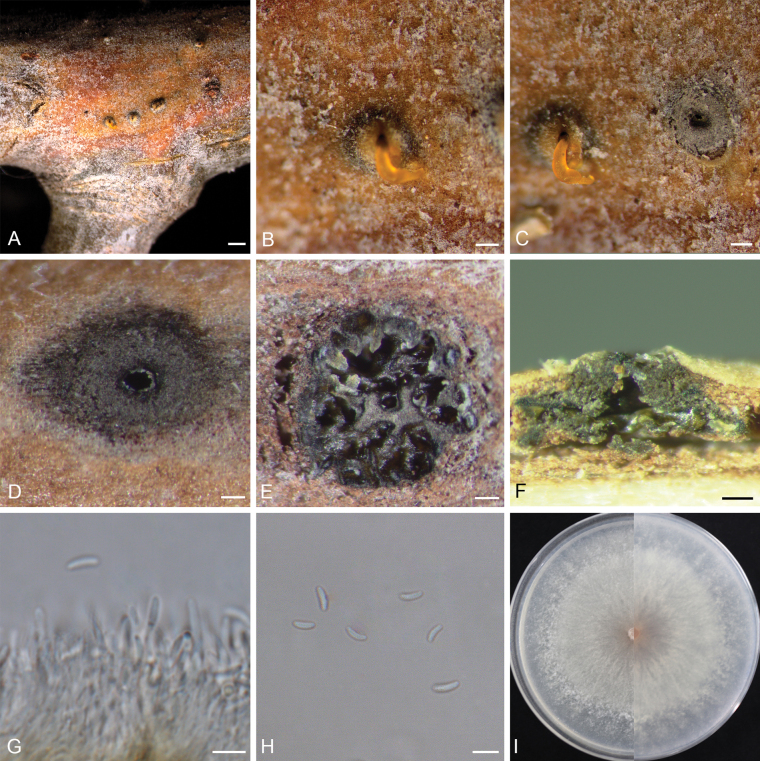 Figure 2
Figure 2 Figure 3
Figure 3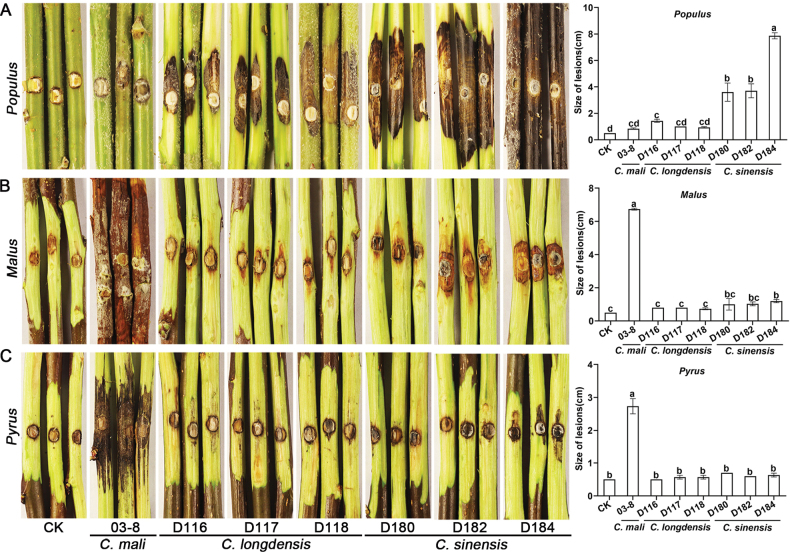 Figure 4
Figure 4| Name | Strain | Host | Origin | GenBank accession numbers | ||||
|---|---|---|---|---|---|---|---|---|
|
|
|
|
|
| ||||
|
| CFCC 58226 |
| Yunnan, China |
|
|
|
|
|
|
| CFCC 89970T |
| Ningxia, China |
|
|
|
|
|
|
| IRAN 4201CT |
| Urmia, Iran |
|
|
|
| NA |
|
| IRAN 4627C |
| Miandoab, Iran |
| NA | NA |
| NA |
|
| CBS 134.25 |
| NA |
|
|
|
|
|
|
| CBS 197.50T |
| UK |
|
|
|
|
|
|
| CFCC 53178T |
| Xinjiang, China |
|
| NA | NA |
|
|
| CFCC 58467T |
| Yunnan, China |
|
|
|
|
|
|
| CFCC 58468 |
| Yunnan, China |
|
|
|
|
|
|
| CFCC 53159T |
| Beijing, China |
|
|
|
|
|
|
| CFCC 58491 |
| Yunnan, China |
|
|
|
|
|
|
| CFCC 58222T |
| Yunnan, China |
| NA |
|
| NA |
|
| CFCC 89993T |
| Shanxi, China |
|
|
|
|
|
|
| CFCC 55855T | Ningxia, China |
| NA |
|
|
| |
|
| CFCC 56037T | Ningxia, China |
| NA |
|
|
| |
|
| C3479 | Xinjiang, China |
|
|
|
|
| |
|
| CFCC 59571T | Xinjiang, China |
|
|
|
|
| |
|
| XJAU 1378T |
| Xinjiang, China |
|
|
|
|
|
|
| XJAU 1379 |
| Xinjiang, China |
|
|
|
|
|
|
| CBS 144235T |
| California, USA |
|
| NA |
|
|
|
| CFCC 58213 |
| Yunnan, China |
|
|
|
|
|
|
| MFLUCC 15-0857T |
| Russia |
|
|
| NA | NA |
|
| CFCC 58193T |
| Yunnan, China |
|
|
|
|
|
|
| CFCC 58221T |
| Yunnan, China |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| CFCC 58706T | Tibet, China |
|
|
|
|
| |
|
| CFCC 58483T |
| Yunnan, China |
|
|
|
| NA |
|
| CFCC 89999 |
| Shanxi, China |
|
|
|
|
|
|
| MFLUCC 16-0628 |
| Russia |
|
|
| NA | NA |
|
| CFCC 89984 |
| Xinjiang, China |
|
|
|
|
|
|
| MFLUCC 15-0851T |
| Russia |
|
|
| NA | NA |
|
| IRAN 4409CT |
| Oshnavieh, Iran |
|
|
| NA | NA |
|
| IRAN 4410C |
| Oshnavieh, Iran |
|
|
| NA | NA |
|
| IRAN 4405CT |
| Miyandoab, Iran |
|
|
| NA | NA |
|
| IRAN 4406C |
| Miyandoab, Iran |
|
|
| NA | NA |
|
| CBS 118.22T |
| Netherlands |
|
|
|
|
|
|
| CBS 195.42 | NA | Switzerland |
|
|
|
|
|
|
| CFCC 58227 |
| Yunnan, China |
|
|
|
|
|
|
| CFCC 58228 |
| Yunnan, China |
|
|
|
|
|
|
| CFCC 55453T |
| Gansu, China |
|
|
|
|
|
|
| CFCC 55455T |
| Gansu, China |
|
|
|
|
|
|
| CFCC 89981T |
| Gansu, China |
|
|
|
|
|
|
| CFCC 89982 |
| Tibet, China |
|
|
|
|
|
|
| CFCC 89909T |
| Gansu, China |
|
|
| NA | NA |
|
| CFCC 89910 |
| Gansu, China |
|
|
|
| NA |
|
| MFLUCC 15-0509T |
| Russia |
|
|
| NA | NA |
|
| MFLUCC 15-0861 |
| Russia |
|
|
| NA | NA |
|
| MFLUCC 14-1052T |
| Russia |
|
| NA | NA | NA |
|
| MFLUCC 15-0866 | Thailand |
|
|
| NA | NA | |
|
| CBS 141629 |
| Iran |
|
|
|
|
|
|
| MFLUCC 15-0862T |
| Russia |
|
|
| NA | NA |
|
| CBS 118570 |
| Michigan, USA |
|
|
|
|
|
|
| MFLUCC 15-0507T |
| Russia |
|
|
| NA | NA |
|
| CFCC 58224T |
| Gansu, China |
|
|
|
|
|
|
| CFCC 58235T |
| Gansu, China |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| CFCC 58464 |
| Yunnan, China |
|
|
|
|
|
|
| CFCC 89600T |
| Gansu, China |
|
|
|
|
|
|
| CBS 450.51T |
| Sweden |
|
|
|
|
|
|
| CBS 141625 | Iran |
|
|
|
|
| |
|
| IRAN 4198C |
| Iran |
|
|
|
| NA |
|
| CBS 149297T |
| USA |
|
|
|
|
|
|
| CBS 149298 |
| USA |
|
|
|
|
|
|
| CFCC 50040T |
| Ningxia, China |
|
|
|
|
|
|
| CFCC 50042T |
| Gansu, China |
|
|
|
|
|
|
| CBS 160.32 |
| USA |
|
| NA |
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Yeasts and Rust Fungi Studies · Mycorrhizal Fungi and Plant Interactions
Introduction
Poplars (Populus spp.) are among the most widely used afforestation and greening species in China due to their strong ecological adaptability, rapid growth, and high-yield timber. Their total plantation area in China exceeds 7.57 million hectares, of which 4.45 million hectares are timber forest (Forest Resources Management Division 2018; Lin et al. 2023). However, canker and dieback caused by some Cytospora species are prevalent in Populus stands, particularly in northern China, resulting in significant ecological and economic losses (Zhuang 2005; Fan et al. 2020; Lin et al. 2023).
The genus Cytospora (Cytosporaceae, Diaporthales) comprises many important plant pathogens, such as Cytosporachrysosperma, C.leucosperma, and C.mali (=Valsamali), etc. These pathogens are associated with canker and dieback disease in herbaceous and woody plants worldwide (Adams et al. 2005; Feng et al. 2023; Lin et al. 2024). The pathogens usually invade the host tissues through small cracks, wounds, and natural openings on the bark surface. Later, they can colonize and decompose the cambium and penetrate extensively into the phloem and xylem, causing cortical tissue rot, slight depressions, cracks, and discoloration (Feng et al. 2023; Azizi et al. 2024). Finally, cankers and diebacks were formed on the infected plants, which can lead to host plant mortality in severe cases (Pan et al. 2021; Feng et al. 2023; Azizi et al. 2024).
Species of Cytospora were often known by their asexual morph: pycnidia stromata submerged in bark, with single or labyrinthiform cavities visible in tangential section, with or without black conceptacles, filamentous conidiophores, phialidic conidiogenous cells, and allantoid hyaline spores (Adams et al. 2005; Lawrence et al. 2018; Fan et al. 2020; Lin et al. 2024). This is largely due to the fact that the asexual state is the one most commonly encountered on diseased plants in nature. In the sexual state, the ascomata are immersed in bark tissues with erumpent pseudostromata; ellipsoid to clavate asci contain four, eight, or more elongate-allantoid hyaline ascospores (Spielman 1985; Adams et al. 2005; Lin et al. 2024).
Traditionally, species of Cytospora were identified based on their morphological characteristics and host associations (Donk 1964; Spielman 1985; Adams et al. 2005). However, the morphological variation between some species was complicated. The spore size of the same species could vary depending on the host plant and climatic conditions (Fan et al. 2020; Lin et al. 2023, 2024). This led to incorrect species identification and the fact that cryptic species were often overlooked (Norphanphoun et al. 2017; Fan et al. 2020).
The emergence of DNA sequencing technology has greatly enhanced our scientific capacity to recognize cryptic species and identify unknown species. A combination of five gene regions, including the nuclear rDNA internal transcribed spacer region (ITS), actin (act), RNA polymerase II subunit (rpb2), translation elongation factor 1-alpha (tef1-α), and β-tubulin (tub2), has been found to provide stable and reliable resolution for the identification of Cytospora species (Norphanphoun et al. 2017, 2018; Lawrence et al. 2018; Fan et al. 2020; Lin et al. 2024). Based on the DNA sequence analysis of these five gene regions, Lin et al. (2024) recently identified 399 Cytospora isolates from 32 countries and proposed 44 novel species.
To date, about 36 species of Cytospora spp. have been reported on poplar trees worldwide, of which 24 are found in China, including C.ailanthicola, C.alba, C.atrocirrhata, C.beijingensis, C.chrysosperma, C.davidiana, C.deqinensis, C.diqingensis, C.donglingensis, C.eastringensis, C.fugax, C.hoffmannii (a synonym of C.nivea and C.paratranslucens), C.kantschavelii, C.lijiangensis, C.palmoides, C.populi, C.pseudochrysosperma, C.sanbaensis, C.shangrilaensis, C.sinensis, C.sophoriopsis, C.translucens, C.tritici, and C.yuduensis (Zhuang 2005; Zhang et al. 2012, 2013; Wang et al. 2013, 2015; Fan et al. 2020; Lin et al. 2023, 2024).
During our recent investigations in the Ningxia Hui Autonomous Region, China, a disease exhibiting symptoms of trunk, crown, and collar canker, as well as branch dieback, was observed in a stand of approximately fifty 10-year-old Populusalbasubsp.pyramidalis trees. The aim of this study was to (1) identify the isolates associated with the canker and dieback disease and (2) evaluate the pathogenicity of the isolates on Populusalbasubsp.pyramidalis (Bunge) W. Wettst, Malusdomestica (Suckow) Borkh. cv. ‘Fuji’ and Pyrusbretschneideri Rehd.
Sample collection and fungal isolation
During 2022–2023, a disease survey was conducted in a 700-m^2^ stand of approximately fifty 10-year-old Populusalbasubsp.pyramidalis trees in Longde County, Ningxia Hui Autonomous Region, China. Three trees exhibiting canker and dieback symptoms were examined. Twigs, branches, and trunk cortex segments with obvious fruiting bodies were sampled, packed in clean paper bags, and transferred to the laboratory for further research.
Under a dissecting microscope (Olympus SZX16, Tokyo, Japan), conidiomata and ascomata formed under the bark on the specimens were sectioned using a sterile scalpel. Mucoid spore masses were transferred directly to 2% malt extract agar (MEA: 20 g malt extract powder, 20 g agar powder, distilled water to complete 1000 mL) using sterile needles. After one day of incubation at 25 °C, a single hyphal tip from each culture was transferred to new MEA plates and incubated at 25 °C for seven days. The herbarium specimens and cultures were deposited in the culture collection of State Key Laboratory for Crop Stress Resistance and High-Efficiency Production, Northwest A&F University (NWAFU).
DNA extraction, PCR amplification, and sequencing
The total genomic DNA was extracted from one-week-old cultures using Prepman™ Ultra Sample Preparation Reagent (Thermo Fisher Scientific, Waltham, MA, USA) following the protocol described by Duong et al. (2012). Five gene regions, including ITS, act, rpb2, tef1-α, and tub2, were amplified using the protocol described by Fan et al. (2020). Amplicons were sequenced in both directions by Sangon Biotech Company Limited, Shanghai, China. The sequences obtained were assembled using Geneious v. 7.0 (Kearse et al. 2012) and subjected to BLASTn searches to preliminarily identify their classification. Genotypes of the isolates were determined based on the sequences of the ITS, act, rpb2, tef1-α, and tub2 gene regions. Isolates from three distinct trees, showing different morphological characteristics and genotypes, were selected for phylogenetic analyses and pathogenicity tests. All sequences generated in this study were deposited in GenBank (Table 1).
Phylogenetic analyses
The referenced sequences of phylogenetically related Cytospora species were downloaded from NCBI (http://www.ncbi.nlm.nih.gov). Diaporthevaccinii (CBS 160.32) was selected as the outgroup taxon (Lin et al. 2024). Sequences generated from this study and those from NCBI were aligned using the MAFFT v. 7 online version (https://mafft.cbrc.jp/alignment/server/) and manually edited in MEGA v. 6.0 (Tamura et al. 2013).
Maximum likelihood (ML) and Bayesian inference (BI) methods were used for phylogenetic analyses on the CIPRES Science Gateway platform (Miller et al. 2010). The sequence datasets for the five individual gene regions were initially analyzed separately to determine the phylogenetic relatedness of all isolates. Subsequently, a multi-gene phylogenetic analysis was conducted for the combined dataset of the five gene regions. The GTR+G+I substitution model and 1000 bootstrap replicates were set for ML analysis in RAxML v. 8.2.12 (Stamatakis 2014). For BI phylogenetic analysis, the appropriate substitution model was generated for each partitioned locus using jModeltest v. 2.1.5 (Guindon and Gascuel 2003; Darriba et al. 2012). Four Markov Chain Monte Carlo (MCMC) chains for 1,000,000 generations were employed for BI analysis in MrBayes v. 3.2.7 (Ronquist et al. 2012). The resultant trees were visualized in FigTree v. 1.4.4 and edited using Adobe Illustrator 2020.
Morphology
To induce sporulation, cultures were grown on 2% water agar (WA) plates containing one-year-old, twice-autoclaved poplar shoots on the surface and incubated at room temperature under near-ultraviolet light (12-h photoperiod). The fungal was examined weekly. The fruiting bodies were sectioned (tangentially and vertically) using sterile razor blades and photographed under an Olympus SZX16 stereomicroscope (Tokyo, Japan). The micro-morphological structures were mounted in a drop of 85% lactic acid (Liu et al. 2020) on microscope slides and examined under an Olympus BX53 microscope (Tokyo, Japan). At least 30 replicate measurements were made for each taxonomic character by cellSens imaging software (Olympus, Tokyo, Japan). Colony characteristics were examined on PDA plates after growth at 25 °C in the dark for seven days. Colony colors were described according to the color charts of Rayner (1970).
Pathogenicity tests
Healthy one-year-old branches (approximately 2 cm in diameter) of Populusalbasubsp.pyramidalis, Malusdomestica cv. ‘Fuji’ and Pyrusbretschneideri were collected from the cultivar conservation nursery of Northwest A&F University (34°15'41.7"N, 108°3'33.3"E). The collected branches were surface-disinfected in a 3% sodium hypochlorite solution for 90 s, rinsed three times with sterile distilled water, and dried at room temperature. The cut ends of the branches (approximately 50 cm long) were sealed with paraffin to minimize desiccation.
A sterilized cork borer (5 mm in diameter) was used to create a 2 mm deep wound in the center of each branch. Five mm in diameter mycelial plugs were excised from a 3-day-old colony of each selected isolate of Cytospora species identified in this study and placed onto the branch wound. The inoculated sites were covered with sterile, moistened cotton balls and wrapped with masking tape to maintain humidity. Sterile MEA served as the negative control, and C.mali 03-8, a pathogen causing Apple Valsa Canker from our laboratory culture collection, was designated as the positive control. Three replicates were conducted for each isolate and control, respectively. All of them were incubated in humid chambers at room temperature (25 ± 2 °C) under natural light conditions. Lesion sizes around the inoculation sites were measured on the fifth and tenth days post-inoculation. To fulfill Koch’s postulates, the fungi were re-isolated from the lesion margins and identified based on colony morphology. The experiment was replicated three times following the same protocol. The data were analyzed using one-way analysis of variance (ANOVA) and Tukey’s multiple comparison test (p < 0.05) in IBM SPSS Statistics v. 22 (IBM Corp., USA).
Results
Isolates and sequencing
A total of nine isolates with typical morphological characteristics of Cytospora species were obtained. BLASTn searches and sequence alignments for the ITS, act, rpb2, tef1-α, and tub2 gene regions confirmed that the nine isolates belong to the genus Cytospora and represent three genotypes. Based on the origin details (specific tree and tissue, collection date), morphological characteristics, and genotypes of the isolates, six isolates were selected for further study (Table 1).
Phylogenetic analyses
Based on the comparisons of phylogenetic tree topologies for each of the five gene regions, sequences of 63 strains representing 35 Cytospora species (Table 1), closely related to the species collected in this study, were selected for multi-gene phylogenetic analyses. The dataset of combined ITS, act, rpb2, tef1-α, and tub2 sequence alignments comprised 2478 characters, including 475 characters for ITS, 252 characters for act, 644 characters for rpb2, 619 characters for tef1-α, and 488 characters for tub2.
For ML analysis, the matrix had 1024 distinct alignment patterns, with 25.07% gaps or undetermined characters. The final ML optimization likelihood value of the best-scoring tree was -14539.91. The estimated base frequencies were as follows: A = 0.250442, C = 0.289329, G = 0.239478, T = 0.220750; substitution rates: AC = 1.420694, AG = 3.784817, AT = 1.797716, CG = 0.974888, CT = 7.890924, GT = 1.000000; and the gamma distribution shape parameter α = 0.304660. For the BI analysis, the best-fit model of nucleotide substitution for each partitioned locus was as follows: ITS = K80+I+G, act = HKY+G, rpb2 = TrNef+G, tef1-α = TrN+G, and tub2 = HKY+I+G.
The tree topology generated from the ML analysis was essentially similar to that produced from the BI analysis. Three isolates (D116, D117, and D118) representing one genotype (genotype I) formed an independent clade distinct from previously known species, supported by high values (ML/BI = 100/1), and were named as Cytosporalongdensis sp. nov. (Fig. 1). Isolates D180 (genotype II), D184 (genotype II), and D182 (genotype III), representing two genotypes with only one SNP difference across the five gene regions between genotype II and genotype III, were grouped in the same clade with the ex-type isolate of C.sinensis CFCC 58235 (ML/BI = 82/0.83) and were identified as Cytosporasinensis (Fig. 1).
Phylogenetic tree of Cytospora based on maximum likelihood analysis of a combined DNA sequence dataset of ITS, act, rpb2, tef1-α, and tub2 genes. Bootstrap values ≥ 50% (maximum likelihood) and posterior probabilities ≥ 0.5 (Bayesian) are presented at the branches. Values lower than these thresholds are marked with “”, and absent are marked with “-”. Type materials are marked with “T”. Isolates from this study are highlighted in blue. The tree was rooted to Diaporthevaccinii (CBS 160.32).*
Taxonomy
Cytospora
longdensis
Taxon classificationFungiDiaporthalesValsaceae
J.Q. Li & L.L. Huang sp. nov.
21291787-CDC5-5936-B0E5-6330464927C5
857641
Etymology.
Named after the locality, Longde County, Guyuan City, where the fungus was first collected.
Cytosporalongdensis (D 116). A–C. View of conidiomata on branch; D, E. Transverse section through a conidioma; F. Longitudinal section through a conidioma; G. Conidiophores, conidiogenous cells, and conidia; H. Conidia; I. Culture on PDA after five days, front side (left) and back side (right). Scale bars: 500 μm (A); 100 μm (B–F); 5 μm (G, H).
Typification.
China • Ningxia Hui Autonomous Region, Guyuan City, Longde County, Shatang Town, 35°35'12.5"N, 106°7'7.4"E, on cankered and diebacked branches of Populusalbasubsp.pyramidalis, 30 July 2022, J.Q. Li & L.L. Huang (holotype: NWAFU H116, ex-holotype culture: D116).
Description.
Sexual morph: not observed. Asexual morph: Conidiomata immersed or semi-immersed in bark, discoid, with multiple locules. Conceptacle absent. Ectostromatic disc greenish black to black, circular to ovoid, (296–)351–508(–603) μm in diameter, with one ostiole per disc. Ostiole black, circular to ovoid, (42–)52–87(–105) μm in diameter. Locules numerous, subdivided by invaginations with common walls. Conidiophores hyaline, thin-walled, unbranched, or occasionally branched at the bases, 15.1–26.3 × 1.0–2.1 μm (av. = 18.9 ± 2.9 × 1.4 ± 0.3 μm, n = 30). Conidiogenous cells enteroblastic, phialidic, subcylindrical to cylindrical. Conidia hyaline, smooth, allantoid, aseptate, 4.2–5.7 × 1.6–2.2 μm (av. = 5.0 ± 0.4 × 1.8 ± 0.2 μm, n = 50).
Culture characteristics.
Colonies at 25 °C on PDA are initially white, flat, reaching 4.7 cm in diameter after three days, becoming buff, and fully covering the 9.0 cm Petri dish after seven days.
Additional materials examined.
China • Ningxia Hui Autonomous Region, Guyuan City, Longde County, Shatang Town, 35°35'12.5"N, 106°7'7.4"E, on cankered branches of Populusalbasubsp.pyramidalis, 30 July 2022, J.Q. Li & L.L. Huang (NWAFU H117, culture: D117; NWAFU H118, culture: D118).
Notes.
Cytosporalongdensis is phylogenetically closely related to C.suecica and C.lhasaensis. However, it is distinct from them by producing larger conidia, i.e., 4.2–5.7 × 1.6–2.2 μm in C.longdensis vs. 0.5–2 × 0.5–1 μm in C.suecica and 3.5–5 × 1–1.5 μm in C.lhasaensis (Lin et al. 2024). Additionally, it differs from C.suecica (CBS 450.51) and C.lhasaensis (CFCC 58706) in SNPs for the five gene region sequences (C.longdensis vs. C.suecica: 23 out of 473 bp in ITS, 28 out of 203 bp in act, 26 out of 644 bp in rpb2, 70 out of 500 bp in tef1-α, and 55 out of 388 bp in tub2; C.longdensis vs. C.lhasaensis: 17 out of 473 bp in ITS, 19 out of 203 bp in act, 25 out of 644 bp in rpb2, 75 out of 500 bp in tef1-α, and 61 out of 388 bp in tub2). Additionally, the multi-gene (ITS, act, rpb2, tef1-α, and tub2) phylogram reveals that C.longdensis represents an independent clade with high support (ML/BI = 100/1, Fig. 1).
Cytospora
sinensis
Taxon classificationFungiDiaporthalesValsaceae
L. Lin & X.L. Fan, Studies in Mycology 109: 381 (2024)
04C7DF46-9D88-5063-9068-EC58B937141A
Description.
Sexual morph: not observed. Asexual morph: Conidiomata immersed in bark, erumpent when mature, discoid, with multiple locules. Conceptacle absent. Ectostromatic disc isabelline or greenish black, circular to ovoid, (285–)305–452(–512) μm in diameter, with single ostiole per disc. Ostiole black, circular to ovoid, (38–)45–84(–103) μm in diameter. Locules numerous, divided with shared walls. Conidiophores hyaline, thin-walled, unbranched, or branched at the bases, 12.5–23.2 × 1.2–2.2 μm (av. = 17.6 ± 2.7 × 1.6 ± 0.2 μm, n = 30). Conidiogenous cells enteroblastic, phialidic, subcylindrical to cylindrical. Conidia hyaline, smooth, allantoid, aseptate, 4.5–5.9 × 1.6–2.1 μm (av. = 5.2 ± 0.4 × 1.8 ± 0.1 μm, n = 50).
Culture characteristics.
Colonies at 25 °C on PDA are initially white, flat, growing up to 6.2 cm in diameter after three days, becoming olivaceous and completely covering the 9.0 cm Petri dish after five days.
Additional materials examined.
China • Ningxia Hui Autonomous Region, Guyuan City, Longde County, Shatang Town, 35°35'12.5"N, 106°7'7.4"E, on cankered and infected branches of Populusalbasubsp.pyramidalis, 30 July 2022, J.Q. Li & L.L. Huang (NWAFU H180, culture: D180; NWAFU H182, culture: D182; NWAFU H184, culture: D184).
Notes.
Cytosporasinensis has been reported from Populus species in Gansu and Yunnan provinces of China (Lin et al. 2024). In the present study, three isolates causing canker and dieback on Populusalbasubsp.pyramidalis in the Ningxia Hui Autonomous Region were identified as C.sinensis based on the phylogenetic topology (Fig. 1) and morphological characteristics consistent with previous descriptions (Fig. 3, Lin et al. 2024).
Cytosporasinensis (D 184). A View of conidiomata on branch; B, C Transverse section through a conidioma; D, E Longitudinal section through a conidioma; F Conidiophores and conidiogenous cells; G Conidia; H Cultures on PDA after five days, front side (left) and back side (right). Scale bars: 500 μm (A); 100 μm (B, C, E); 50 μm (D); 5 μm (F, G).
Pathogenicity tests
Inoculation of Populusalbasubsp.pyramidalis with conidial masses of C.longdensis and C.sinensis resulted in the persisting symptoms of canker after five days. No considerable disease symptoms were observed for the negative control (Fig. 4A). Moreover, the lesions on Populusalbasubsp.pyramidalis caused by C.sinensis were significantly larger than those caused by C.longdensis. However, both species were found to be essentially non-pathogenic to Malusdomestica and Pyrusbretschneideri, as no clear disease symptoms were observed at the inoculated sites (Fig. 4B, C).
The lesions caused by Cytospora isolates on branches of Populus, Malus, and Pyrus five days post-inoculation. A.Cytosporalongdensis (D116, D117, and D118), C.sinensis (D180, D182, and D184), and C.mali (03-8) on Populusalbasubsp.pyramidalis; B. on Malusdomestica cv. ‘Fuji’; C. and Pyrusbretschneideri. Vertical bars represent the standard error of the mean. Different letters above the bars indicate treatment mean values that are significantly different (P = 0.05).
Discussion
In this study, we identified two Cytospora species, C.sinensis and a novel species described as C.longdensis, associated with canker and dieback in a stand of approximately fifty 10-year-old Populusalbasubsp.pyramidalis trees in Ningxia, China. Applied pathogenicity assays confirmed that both species were pathogenic to Populusalbasubsp.pyramidalis through nine experimental replicates, with C.sinensis exhibiting significantly stronger virulence than C.longdensis. In contrast, both species showed negligible pathogenicity on Malusdomestica and Pyrusbretschneideri.
The co-infection of C.sinensis and C.longdensis was observed in a small stand of Populusalbasubsp.pyramidalis in Ningxia, China. This finding aligns with previous studies that have reported multiple Cytospora species infecting one plant host (Kepley et al. 2015; Lin et al. 2022). As a typical arid to semi-arid region in northwest China, Ningxia has an average annual temperature ranging from 6.9 °C to 11.5 °C (Zhang et al. 2025). The co-infection phenomenon may be linked to environmental stressors, such as drought, frost, and soil salinity, which can weaken host defenses and potentially facilitate co-infections (Adams et al. 2005; Lawrence et al. 2017, 2018; Lin et al. 2024). Understanding the diversity of these fungi is crucial for a possible clarification of their adaptive co-evolution and developing control strategies. Multi-gene phylogenetic analysis combined with morphological comparison provides reliable resolution for identifying cryptic species within the genus Cytospora on the same hosts (Norphanphoun et al. 2018; Lin et al. 2024).
Distinct virulence was observed between C.sinensis and C.longdensis. This suggests that the two species may employ different pathogenic mechanisms or display varying levels of host adaptation (Adams et al. 2005; Sun et al. 2023). Cytosporasinensis exhibited stronger virulence, indicating it may play a more dominant role in causing canker and dieback in some Populus stands (Lin et al. 2023). In contrast, C.longdensis showed weaker virulence, suggesting it may act as a secondary pathogen or require specific environmental conditions to cause significant damage (Adams et al. 2005; Lin et al. 2023). More extensive surveys and further studies are needed to confirm these findings and elucidate the genetic and molecular basis of these virulence differences.
Host preference was observed in this study. Phylogenetic analyses suggested that C.sinensis resides in the C.chrysosperma complex and is closely related to C.chrysosperma. Within the C.chrysosperma complex, several species, including C.chrysosperma and C.hoffmannii, have been identified as pathogens of Salicaceae and Rosaceae (Azizi et al. 2024; Lin et al. 2024). However, Cytosporasinensis isolated from Populus exhibited limited pathogenicity on Malus and Pyrus. Its highest virulence towards Populus indicates host preference in pathogenicity (Adams et al. 2005; Schulze-Lefert and Panstruga 2011).
Cytosporasinensis was recently reported on Populus spp. in Gansu and Yunnan provinces, but its pathogenic potential had not yet been confirmed (Lin et al. 2024). Our study fills this gap by confirming the virulence of C.sinensis towards Populusalbasubsp.pyramidalis and extending its known geographic distribution to Ningxia, China. The distribution of this species, spanning from northwestern to southwestern China, suggests its strong ecological adaptability. More extensive sampling is required to assess the host range and geographic distribution of C.sinensis. Targeted management strategies are needed to control its spread and mitigate associated damages.
This study contributed to the description of a new species, C.longdensis, and expanded the known geographical distribution and host-fungus interaction of C.sinensis on Populusalbasubsp.pyramidalis. Further studies are required to elucidate the genetic and molecular mechanisms driving their differences in pathogenicity. Moreover, the high virulence and wide distribution range of C.sinensis emphasize the urgency of developing targeted control strategies to manage these fungi effectively.
Supplementary Material
XML Treatment for Cytospora longdensis
XML Treatment for Cytospora sinensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adams GC Wingfield MJ Common R Roux J (2005) Phylogenetic relationships and morphology of Cytospora species and related teleomorphs (Ascomycota, Diaporthales, Valsaceae) from Eucalyptus. Studies in Mycology 52: 1–44.
- 2Azizi R Ghosta Y Ahmadpour A (2024) Cytospora species associated with apple canker and dieback disease in Iran with the description of three new species.Mycological Progress 23(1): 1–30. 10.1007/s 11557-024-02000-z · doi ↗
- 3Darriba D Taboada GL Doallo R Posada D (2012) j Model Test 2: More models, new heuristics and parallel computing. Nature Methods 9: 772. 10.1038/nmeth.2109 PMC 459475622847109 · doi ↗ · pubmed ↗
- 4Donk MA (1964) Nomina conservanda proposita I. Proposals in fungi: Deuteromycetes.Regnum Vegetabile 34: 7–15.
- 5Duong TA De Beer ZW Wingfield BD Wingfield MJ (2012) Phylogeny and taxonomy of species in the Grosmanniaserpens complex.Mycologia 104(3): 715–732. 10.3852/11-10922123658 · doi ↗ · pubmed ↗
- 6Fan XL Bezerra JD Tian CM Crous PW (2020) Cytospora (Diaporthales) in China.Persoonia 45(1): 1–45. 10.3767/persoonia.2020.45.0134456370 PMC 8375343 · doi ↗ · pubmed ↗
- 7Feng H Wang C He Y Tang L Han P Liang J Huang L (2023) Apple Valsa canker: Insights into pathogenesis and disease control.Phytopathology Research 5(1): 45. 10.1186/s 42483-023-00200-1 · doi ↗
- 8Forest Resources Management Division (2018) National forest resources statistics-main results of the ninth national forest resources inventory in China. Beijing, China.