Description of a new species of Pseudobornella Baba, 1932 (Gastropoda, Nudibranchia, Dendronotidae) from the Yellow Sea

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
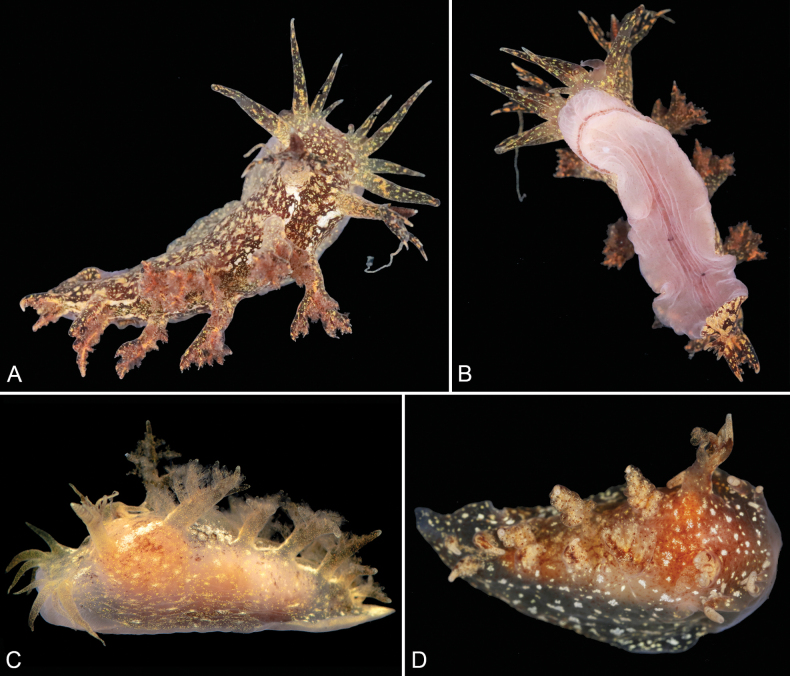 Figure 1
Figure 1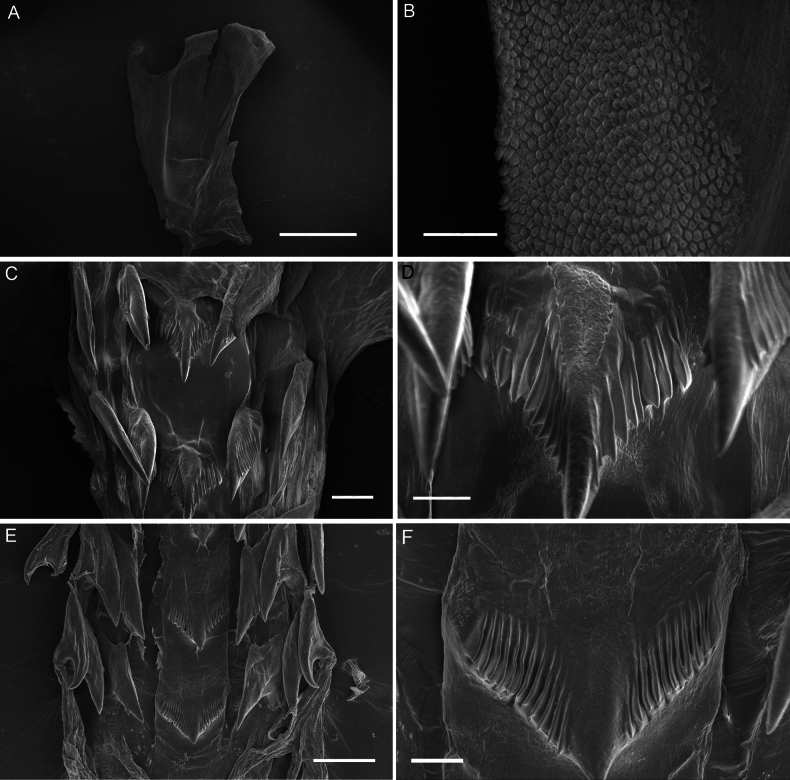 Figure 2
Figure 2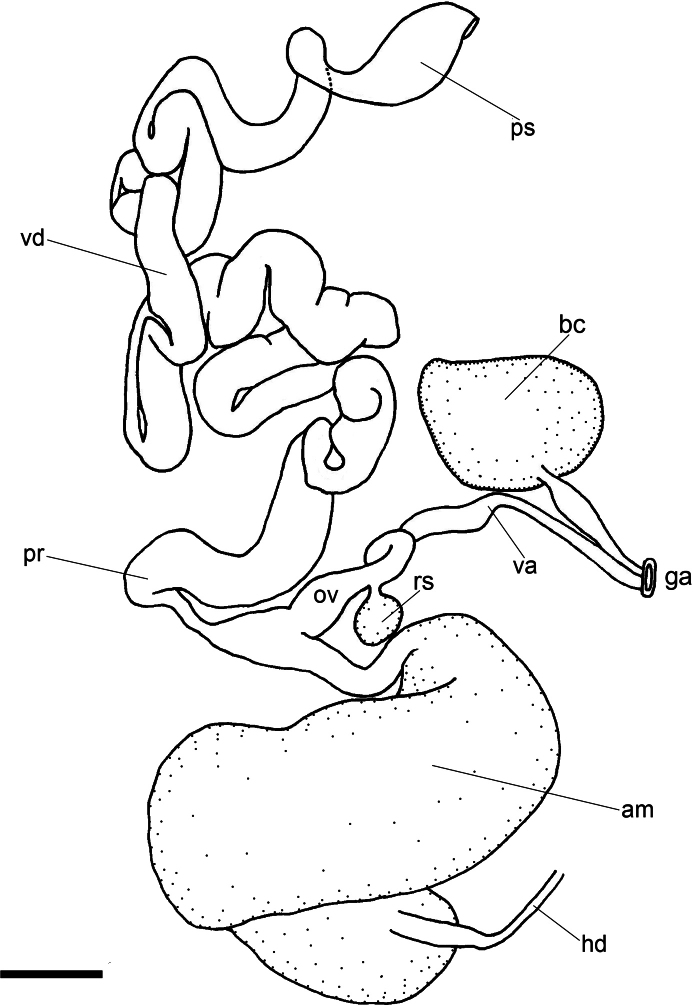 Figure 3
Figure 3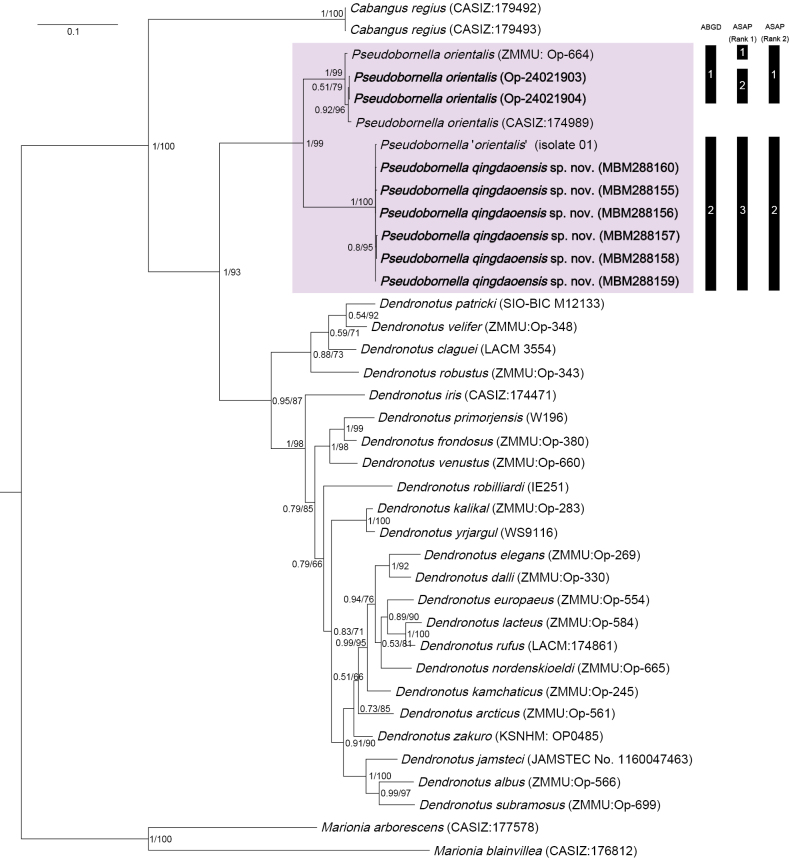 Figure 4
Figure 4| Species name | Voucher | Locality |
| 16S |
| Reference |
|---|---|---|---|---|---|---|
|
| ZMMU:Op-566 | USA: Washington |
|
| — |
|
|
| ZMMU:Op-561 | Russia: Laptev Sea |
|
| — |
|
|
| LACM 3554 | Mexico | — | — |
|
|
|
| ZMMU:Op-330 | Russia: Kamchatka |
|
|
|
|
|
| ZMMU:Op-380 | Norway |
|
|
|
|
|
| ZMMU:Op-269 | Russia: White sea |
|
|
|
|
|
| ZMMU:Op-554 | Norway |
|
| — |
|
|
| CASIZ:174471 | USA: Washington |
|
|
|
|
|
| JAMSTEC No. 1160047463 | Japan |
|
| — |
|
|
| ZMMU:Op-283 | Russia: Kamchatka |
|
|
|
|
|
| ZMMU:Op-245 | Russia: Kamchatka |
|
|
|
|
|
| ZMMU:Op-584 | Norway |
|
| — |
|
|
| ZMMU:Op-665 | Russia: Laptev sea |
|
| — |
|
|
| SIO-BIC M12133 | USA: California |
|
| — |
|
|
| W196 | — |
|
|
|
|
|
| IE251 | Russia: Kamchatka |
|
|
|
|
|
| ZMMU:Op-343 | Russia: Barents Sea |
|
|
|
|
|
| LACM:174861 | USA: Alaska |
|
|
|
|
|
| ZMMU:Op-699 | USA: Washington |
|
| — |
|
|
| ZMMU:Op-348 | Russia: Kara Sea |
|
| — |
|
|
| ZMMU:Op-660 | USA: Washington |
|
| — |
|
|
| WS9116 | Russia: Kara Sea |
|
|
|
|
|
| KSNHM: OP0485 | Japan |
|
|
|
|
|
| ZMMU: Op-664 | Russia: Sea of Japan |
|
| — |
|
|
| CASIZ:174989 | China: Daisong Bay | — |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| isolate 01 | China: Qingdao |
| — | — |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| CASIZ:179492 | Philippines |
|
|
|
|
|
| CASIZ:179493 | Philippines |
|
|
|
|
|
| CASIZ:177578 | Philippines |
|
|
|
|
|
| CASIZ:176812 | Portugal |
|
|
|
|
- —National Natural Science Foundation of China 501100001809 https://ror.org/01h0zpd94 http://doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine Biology and Ecology Research · Marine Bivalve and Aquaculture Studies · Ocean Acidification Effects and Responses
Introduction
Pseudobornella was originally proposed by Baba (1932) to include Pseudobornellaorientalis Baba, 1932, a species of central Japan. The genus was originally placed in the family Bornellidae Bergh, 1874 (Baba 1932, 1933; Pola et al. 2009). Based on molecular evidence, Pola and Gosliner (2010) transferred P.orientalis to Dendronotus Alder & Hancock, 1845, a genus of the family Dendronotidae. This familial placement was accepted by subsequent studies, but Martynov et al. (2015) argued that P.orientalis could be distinguished from Dendronotus species by the presence of remarkably elongated papillae of the rhinophoral sheath and the absence of distinct tertiary branches of the dorsolateral appendages. Korshunova et al. (2020) presented a review of the Dendronotidae using morphological and molecular data and reinstated Pseudobornella as a valid genus. To date, P.orientalis remains the only recognized species within the genus and has a distribution spanning Japan (Baba 1932; Pola et al. 2009), China (Song et al. 2009), and the Far East of Russia (Korshunova et al. 2020).
In recent years, several nudibranch sea slugs were collected from the intertidal zone of Shandong Province, China. Through morphological study and genetic analyses, these specimens were identified as representing an undescribed species belonging to the genus Pseudobornella. In the present study, we describe the species and compare it with P.orientalis.
Materials and methods
Sampling and preservation
Ten specimens of the new species were collected from the intertidal zones of Rushan and Qingdao in Shandong Province, China. The live animals were photographed and preserved in 99% ethanol for molecular study and 4% formalin for morphological dissections. Type specimens were deposited at the Marine Biological Museum, Chinese Academy of Sciences (MBMCAS).
Light and scanning electron microscopy
External morphology and internal anatomy were examined, dissected, and photographed using a dissecting microscope (ZEISS Discovery V20). For scanning electron microscopy (SEM) analysis, jaws and radulae were dissected from the buccal bulb. They were then cleaned with 10% NaOH, rinsed in distilled water, air-dried, coated with gold, and examined under SEM at an accelerating voltage of 5 kV.
Molecular analyses
Six individuals of the new species and two specimens of Pseudobornellaorientalis were subjected to molecular analysis. Genomic DNA from each individual was extracted using the Column Genomic DNA Isolation Kit (Beijing TIANGEN, China) following to the manufacturer’s instructions. Polymerase chain reactions (PCRs) were conducted in a total volume of 25 μl, including 2 μl DNA template, 0.5 μl of each 10 mM primers, 0.5 μl of 10 mM dNTPs, 2.5 μl of 10× buffer, and 0.5 U Taq DNA polymerase. Thermal cycling was performed under the following conditions: 94 °C for 2 min (initial denaturation); followed by 30 cycles of 94 °C for 30 s (denaturation), 42 °C for 45 s (annealing), and 72 °C for 60 s (extension); and a final extension at 72 °C for 5 min. The cytochrome c oxidase subunit I gene (COI) was amplified by polymerase chain reaction (PCR) using the primers LCO1490 (forward: 5′-GGTCAACAAATCATAAAGA TATTGG-3′) and HCO2198 (reverse: 5′-TTAACTTCAGGGTGACCAAAAAATCA-3′) (Folmer et al. 1994), the 16S ribosomal RNA (16S rRNA) was amplified by the primers 16Sar (forward: 5’-CGCCTGTTTATCAAA AACAT-3’) and 16Sbr (reverse: 5’-CTCCGGTTTGAACTCAGATCA-3’) (Palumbi 1996), and the histone 3 gene (H3) was amplified by the primers HexAF (forward: 5’-ATGGCTCGTACCAAGCAGACGGC-3’) and HexAR (reverse: 5’-ATATCCTTGGGCATGATGGTGAC-3’) (Colgan et al. 1998). PCR products were verified on a GelGreen-stained 1.5% agarose gel and sequenced using the BigDye Terminator Cycle Sequencing Kit (v. 3.1 Applied Biosystems, USA) and an AB PRISM 3730 (Applied Biosystems, USA) automatic sequencer.
Phylogenetic analyses
Three partial gene sequences (COI, 16S, and H3) were obtained from each specimen and deposited in the GenBank. Additionally, sequences of other dendronotid species were retrieved from GenBank and used for phylogenetic analyses (Table 1). Two tritoniid species, Marioniablainvillea (Risso, 1818) and Marioniopsisarborescens (Bergh, 1890), were used to root the phylogenetic tree.
Sequences were aligned with MAFFT (Katoh and Standley 2013) using ‘G-INS-i (accurate)’ strategy and normal alignment mode. Separate analyses were conducted for COI (634 bp), 16S (401 bp), H3 (302 bp), and concatenated data (1328 bp). Ambiguously aligned fragments of 16S alignment were removed using Gblocks (Talavera and Castresana 2007). Maximum-likelihood phylogenies (ML) were inferred using IQ-TREE (Nguyen et al. 2015) under Edge-unlinked partition model for 10,000 ultrafast bootstraps (Minh et al. 2013), as well as the Shimodaira–Hasegawa-like approximate likelihood ratio test (Guindon et al. 2010). Bayesian-inference (BI) phylogenies were inferred using MrBayes v. 3.2.6 (Ronquist et al. 2012) under partition model (2 parallel runs, 5 million generations), in which the initial 25% of sampled data were discarded as burn-in. The best-fit models of evolution (GTR+F+I+G4 for COI and 16S, GTR+F+I for H3) were determined by ModelFinder (Kalyaanamoorthy et al. 2017) using AIC criterion. Results were visualized using FigTree v. 1.4.3. The p-distances within and among each species grouping were estimated with MEGA 6 (Tamura et al. 2013) based on the Kimura 2-parameter (K2P) model (Kimura 1980).
Species delimitation
The Automatic Barcode gap Discovery (ABGD) (Puillandre et al. 2012) and the Assemble Species by Automatic Partitioning (ASAP) (Puillandre et al. 2021) methods were used to assess the number of Pseudobornella species. The alignment from the fast-evolving COI gene was uploaded to the online servers of ABGD (https://bioinfo.mnhn.fr/abi/public/abgd/abgdweb.html) and ASAP (https://bioinfo.mnhn.fr/abi/public/asap), respectively. The analyses were performed with the model of Jukes-Cantor (JC69) with default settings.
Systematics
Order Nudibranchia Cuvier, 1817
Superfamily Dendronotoidea Allman, 1845
Family Dendronotidae Allman, 1845
Pseudobornella
Taxon classificationAnimaliaNudibranchiaDendronotidae
Genus
Baba, 1932
89D7A121-500F-51C8-847A-0CC4EA4313B1
Type species.
Pseudobornellaorientalis Baba, 1932, by original designation.
Type locality.
Kanagawa Prefecture, Japan.
Pseudobornella
qingdaoensis
sp. nov.
Taxon classificationAnimaliaNudibranchiaDendronotidae
B7FCCAAF-DFD7-5A81-ADA0-B2EF6C9CAA80
https://zoobank.org/3213D127-1786-4FA9-9BE7-9FEE128E1538
Type material.
Holotype (Fig. 1A, B): China • 1 specimen, length alive 40 mm; dissected; Shandong Province, Qingdao, Golden Beach Park, intertidal zone; 35°57'03"N, 120°14'26"W; 18 Jan. 2025; Shu-Qian Zhang leg.; MBM288155. Paratypes 1–4: China • 4 specimens, lengths 35–40 mm; complete; complete; Shandong Province, Qingdao, Golden Beach Park, intertidal zone; 35°57'03"N, 120°14'26"W; 20 Jan. 2025; Shu-Qian Zhang leg.; MBM288156–MBM288159. Paratype 5 (Fig. 1C): China • 1 specimen, preserved length 10 mm; dissected; Shandong Province, Qingdao, Taiping Bay, intertidal zone; 11 Apr. 2024; Ju-Hao Wang leg.; MBM288160. Paratype 6 (Fig. 1D): China • 1 specimen, preserved lengths 20 mm; dissected; Shandong Province, Rushan, intertidal zone; 19 Apr. 2019; Ju-Hao Wang leg.; MBM288161. Paratypes 7–10: China • 4 specimens, preserved lengths 20 mm; complete; same collection data as paratypes 6; MBM288162–MBM288165.
Pseudobornellaqingdaoensis sp. nov. A, B holotype, MBM288155, length 40 mm C paratype 5, MBM288160, preserved length 1.0 cm D paratype 6, MBM288161, preserved length 20 mm.
Diagnosis.
A large Pseudobornella reaching 40 mm in body length. Background color generally translucent yellow to dark brown, with numerous scattered orange to brown spots and white blotches. Radula formula 11 × 3.0.3.
Description.
External morphology (Fig. 1). Living animals up to 40 mm in length. Foot wide and tapering relatively abruptly towards the tail. Background color generally translucent yellow to dark brown, with numerous scattered orange to brown spots and white blotches. In some individuals, the white blotches forming two longitudinal streaks running along both sides of the dorsum. Anterior margin of head rounded. Each side of the mouth equipped with four or five smooth, tapering oral tentacles of varying size. Rhinophore sheath very elongate. Upper edge of each rhinophore sheath bearing four lateral digitiform branched papillae and one posterior papilla that is remarkably longer. Rhinophores with ~12 lamellae. Dorsal margin with four pairs of dorsolateral papillae, decreasing in size towards the posterior end of the foot. Each papilla with numerous unbranched gills attached to its inner side. Anus small, located on the right side of the dorsum between the first and second pair of dorsolateral appendages. Genital opening located on the right side, midway between the rhinophore sheath and the first dorsolateral appendage. Foot pale pink, scattered with numerous very small, indistinct darker spots, separated from the head by a long, transversal groove.
Internal anatomy. Jaws (Fig. 2A, B) elongate, inner margin (masticatory margin) terminating distally in a small, pointed process. Masticatory margin equipped with rows of short, cone-shaped rodlets. Radular (Fig. 2C–F) formula 11 × 3.0.3. Rachidian teeth stout, with a strong and bluntly pointed cusp and ~9–12 denticles on both sides of the cusp. Innermost lateral teeth with a very strong elongate and sharp cusp and serrated sides. Outer lateral teeth simple and hamate, and outermost one smallest.
Jaws, radulae of Pseudobornellaqingdaoensis sp. nov. A jaw, MBM288155 B jaw masticatory border, MBM288155 C, D radula, MBM288155 E, F radula, MBM288161. Scale bars: 1 mm (A); 50 μm (B, C); 20 μm (D, F); 100 μm (E).
Reproductive system (Fig. 3) located in the front right corner of the body. Ampulla very large, sausage-shaped, one end connecting with ovotestis via a thin hermaphroditic duct, the other end divided into two different, thin ducts: the short oviduct entering the female gland mass and the second duct branching into a wide, short prostate which extends into a long, folded vas deferens, narrowly opening to a wide and stout penial sac. Two ducts departing from the genital aperture: vagina relatively long and simply folded, connecting with a small, rounded receptaculum seminis; second duct wider and shorter, connecting with a very large pyriform bursa copulatrix.
Reproductive system of Pseudobornellaqingdaoensis sp. nov., with ovotestis and female gland mass removed, MBM288155. Abbreviations: am–ampulla; bc–bursa copulatrix; ga–genital aperture; hd–hermaphrodite duct; ov–oviduct, pr–prostate, ps–penial sac; rs–receptaculum seminalis; va–vagina; vd–vas deferens. Scale bar: 1 mm.
Etymology.
The new species is named after its type locality, Qingdao.
Distribution and habitat.
Known from Yellow Sea, China, and possibly also distributed in Japan (Kinoshita 2002; Nobuhiko 2017). It was found in the rocky intertidal zone, feeding on a hydroid (Kinoshita 2002).
Molecular support
The phylogenetic trees inferred using BI and ML criteria were generally congruent (Fig. 4). The genus Pseudobornella was recovered as monophyletic with high support (PP = 1, BS = 99). Within the genus, P.qingdaoensis sp. nov. formed an independent clade (PP = 1, BS = 100) from P.orientalis. This result supports the systematic placement of the new species in the genus Pseudobornella and its separation from P.orientalis.
Phylogenetic tree inferred by Bayesian analysis (BI) and maximum likelihood (ML) based on concatenated dataset of COI, 16S and H3 genes. Numbers adjacent to nodes refer to BI posterior probability (PP > 0.5) and ML bootstrap scores. Numbers in the vertical black bars indicate the results of ABGD and ASAP species delimitations.
The ABGD analysis resulted in the delimitation of two species. The prior maximal distance (p-distance) ranged between 0.0017 and 0.1. However, the ASAP analysis recovered three partitions with the lowest score (generally considered to be the most supported partition scheme) (Fig. 4; Suppl. material 1). The partition with the second highest score was chosen here as it avoided the splitting of two populations of P.orientalis with a pairwise genetic distance of only 0.6%. In both cases, P.qingdaoensis sp. nov. was separated from P.orientalis.
Based on the available molecular data, the analysis of a 634-bp fragment of the COI gene yielded a pairwise distance of 11.7–12.5% between P.qingdaoensis sp. nov. and P.orientalis, a divergence much higher than the known intraspecific variation (0–0.6%) (Suppl. material 2), further supporting the separation of P.qingdaoensis sp. nov. from P.orientalis.
Discussion
In this study, we incorporated the recently obtained molecular data for Pseudobornellaorientalis and the new species described here into the phylogenetic analyses. The results show that the family Dendronotidae was divided into three distinct clades with high support, corresponding to the three genera currently recognized: Dendronotus Alder & Hancock, 1845; Pseudobornella Baba, 1932; and Cabangus Korshunova, Bakken, Grøtan, K.B. Johnson, Lundin & Martynov, 2020. This topology is consistent with that of a previous study by Korshunova et al. (2020). Our results once again confirmed the validity of Pseudobornella. During the molecular analysis, we found a COI sequence of P.orientalis (accession number: OQ573562) derived from a specimen collected in Qingdao. However, this is a misidentification, as the external morphology of this specimen is clearly different from P.orientalis but consistent with P.qingdaoensis sp. nov. (photograph, Wei pers. comm., 7 Jun. 2024). The COI sequence of this specimen is almost identical to that of the new species, with only 0.2–0.5% divergence, further proving that they belong to the same species.
Despite the close resemblance between P.orientalis and P.qingdaoensis sp. nov., a comparison of their external morphology and internal anatomy allows for a clear differentiation between the two species. In terms of external coloration, the new species has prominent white spots scattered over the body surface and lacks the yellow diagonal stripes on the dorsal body surface that are characteristic of P.orientalis. In its internal anatomy, P.qingdaoensis sp. nov. differs from P.orientalis in having a distinct radula with three instead of two lateral teeth. Additionally, the vas deferens of P.qingdaoensis sp. nov. is significantly longer than that of P.orientalis. In Japan, some recorded individuals identified as P.orientalis are very similar to P.qingdaoensis sp. nov. in color pattern and number of oral tentacles (Kinoshita 2002; Nobuhiko 2017) and thus may be conspecific with the new species. If this is the case, the distribution of P.qingdaoensis sp. nov. would extend to Japan.
Pseudobornellaorientalis was first described in 1932 from Japan. Since then, no additional species have been described within the genus. A review of the literature and online photographic records suggests that the biodiversity of this group may be largely underestimated. As previously noted by Pola et al. (2009), some specimens identified as P.orientalis on the Sea Slug Forum (Rudman 2002) exhibit some differences, such as longer lateral papillae on the rhinophore sheath, indicating that these specimens may represent different species. Some naturalists have documented P.orientalis in the Northeastern Pacific (e.g. Johnson 2016; Pomeroy 2016; Young 2016; Agarwal 2017), but those specimens differ in the number of oral tentacles (five pairs) compared to those from the Western Pacific specimens (three pairs). The specimens are also similar to the new species by having five oral tentacles, but they differ in having a reticulated pattern of yellow stripes on the body surface and potentially represent another distinct species. Huang and Huang (2021) reported an undescribed species of Pseudobornella from Kinmen, China, but this species is easily distinguishable by its translucent whitish body with scattered purple-brown spots and oral tentacles with alternating purple-brown and white bands. Further integrative taxonomic work is needed to accurately determine the biodiversity of this group.
Supplementary Material
XML Treatment for Pseudobornella
XML Treatment for Pseudobornella qingdaoensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agarwal M (2017) First record of Dendronotusorientalis (Baba, 1932) (Nudibranchia: Dendronotidae) in the temperate Eastern Pacific.Bio Invasions Records 6(2): 135–138. 10.3391/bir.2017.6.2.08 · doi ↗
- 2Baba K (1932) Pseudobornellaorientalis, nov. gen. et sp. from Japan.Annotationes Zoologicae Japonenses 13(4): 369–376.
- 3Baba K (1933) Supplementary note on the nudibranchia collected in the vicinity of the Amakusa Marine Biological Laboratry.Annotationes Zoologicae Japonenses 14: 273–283.
- 4Colgan DJ Mc Lauchlan A Wilson GDF Livingston SP Edgecombe GD Macaranas J Cassis G Gray MR (1998) Histone H 3 and U 2 sn RNA DNA sequences and arthropod molecular evolution.Australian Journal of Zoology 46(5): 419–437. 10.1071/ZO 98048 · doi ↗
- 5Ekimova I Korshunova T Schepetov D Neretina T Sanamyan N Martynov A (2015) Integrative systematics of northern and arctic nudibranchs of the genus Dendronotus (Mollusca, Gastropoda), with descriptions of three new species.Zoological Journal of the Linnean Society 173(4): 841–886. 10.1111/zoj.12214 · doi ↗
- 6Ekimova I ValdésÁSchepetov D Chichvarkhin A (2016 a) Was Gordon Robilliard right? Integrative systematics suggest that Dendronotusdiversicolor (multicolor frond-aeolis) is a valid species.Canadian Journal of Zoology 94(11): 793–799. 10.1139/cjz-2016-0096 · doi ↗
- 7Ekimova I Schepetov D Chichvarkhina O Chichvarkhin AY (2016 b) Nudibranch molluscs of the genus Dendronotus Alder et Hancock, 1845 (Heterobranchia: Dendronotina) from northwestern Sea of Japan with description of a new species.Zoologia Bespozvonocnyh 13(1): 15–42. 10.15298/invertzool.13.1.02 · doi ↗
- 8Ekimova I ValdésÁChichvarkhin A Antokhina T Lindsay T Schepetov D (2019) Diet-driven ecological radiation and allopatric speciation result in high species diversity in a temperate-cold water marine genus Dendronotus (Gastropoda: Nudibranchia). Molecular Phylogenetics and Evolution 141: 106609. 10.1016/j.ympev.2019.10660931494182 · doi ↗ · pubmed ↗