Morphology and multigene phylogeny reveals five new species of Hydnaceae (Cantharellales, Basidiomycota) from China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
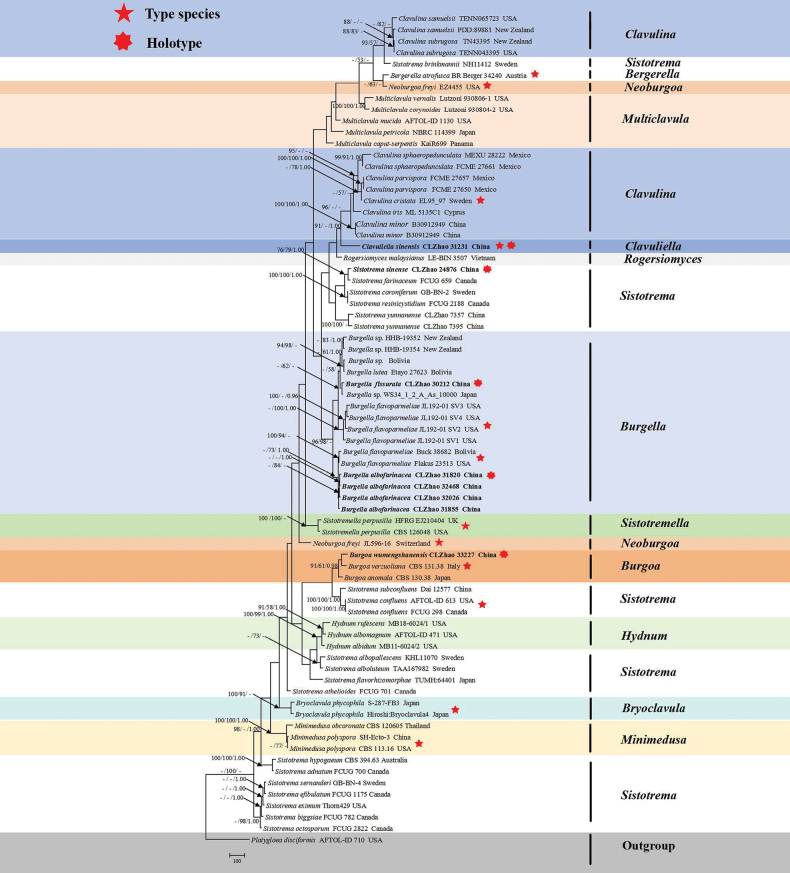 Figure 1
Figure 1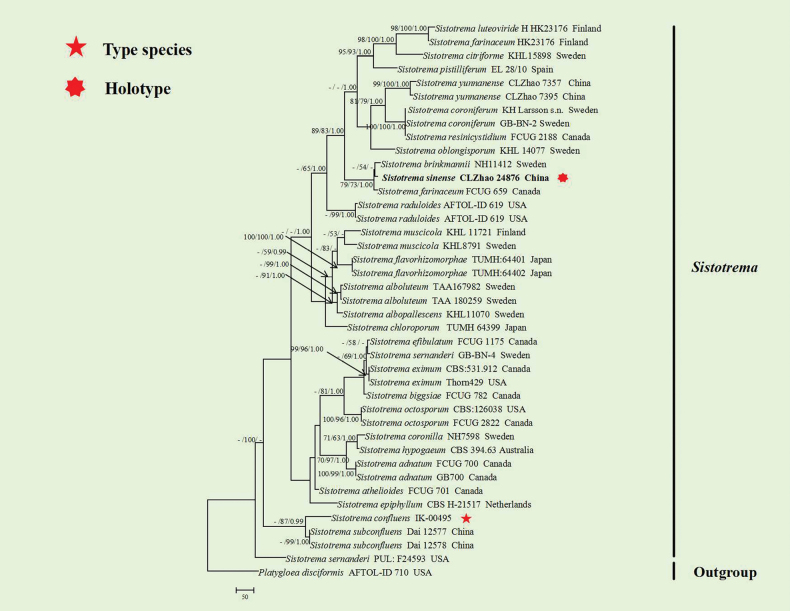 Figure 2
Figure 2 Figure 3
Figure 3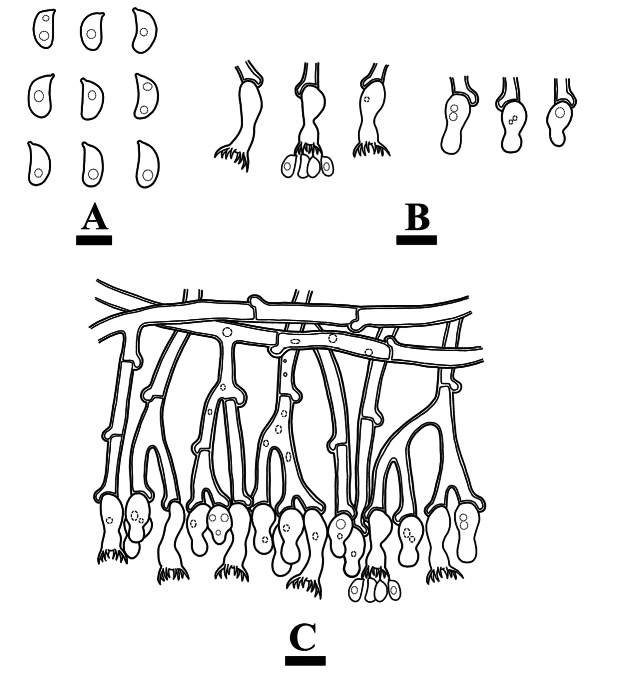 Figure 4
Figure 4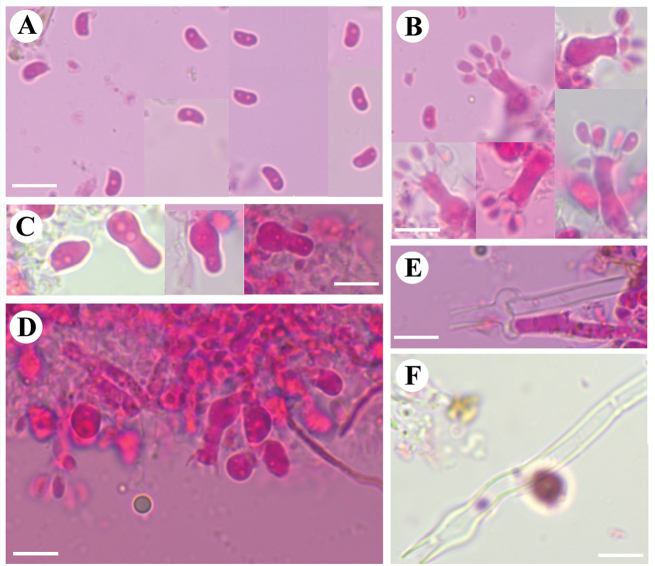 Figure 5
Figure 5 Figure 6
Figure 6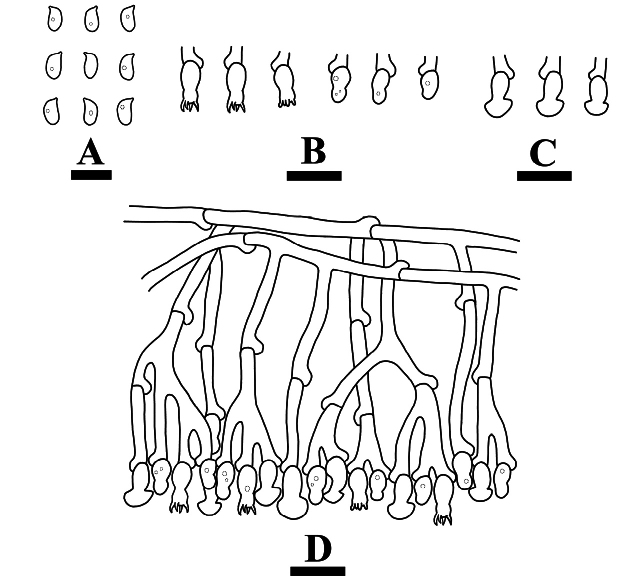 Figure 7
Figure 7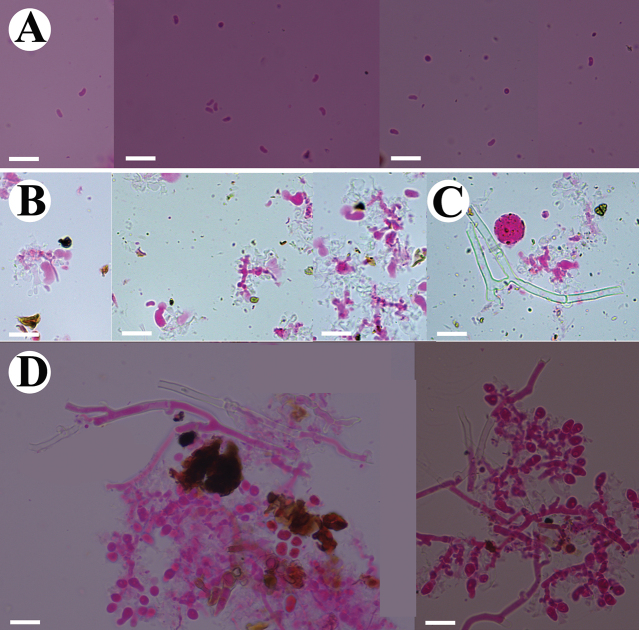 Figure 8
Figure 8 Figure 9
Figure 9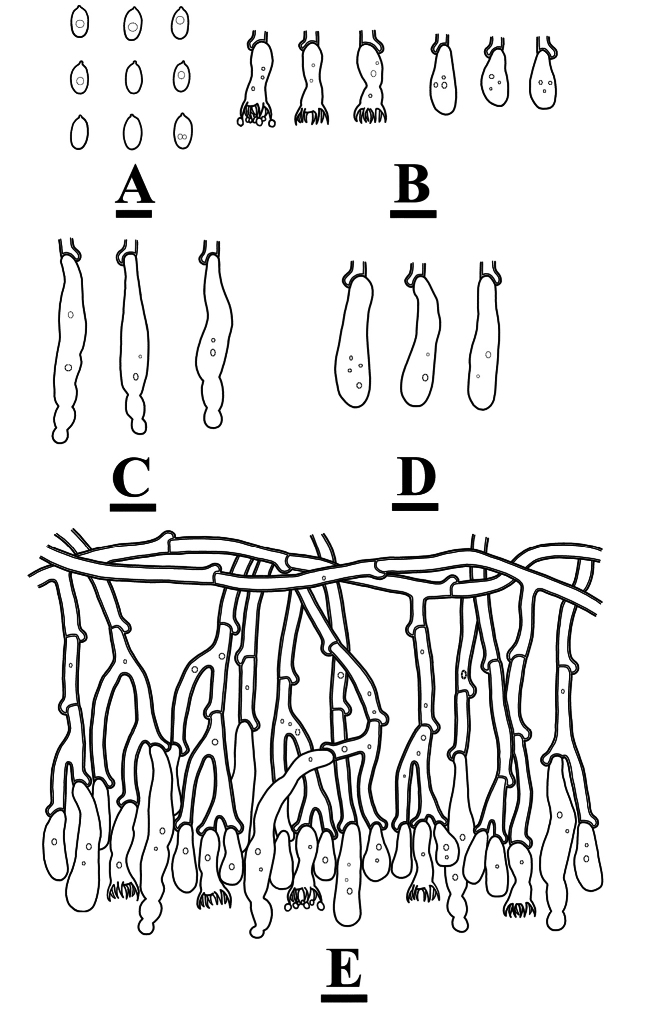 Figure 10
Figure 10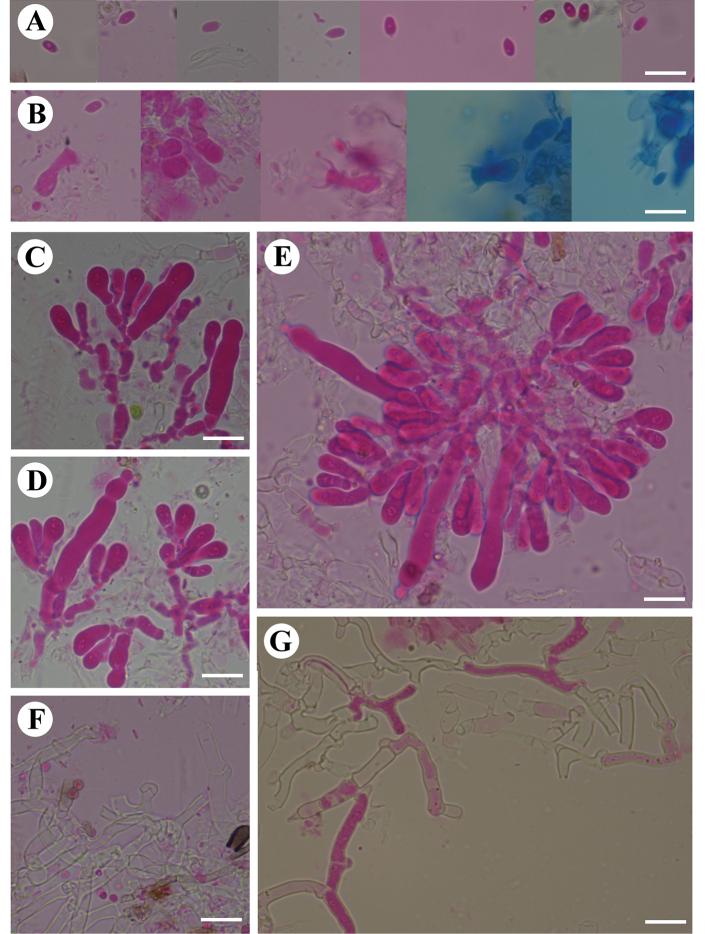 Figure 11
Figure 11 Figure 12
Figure 12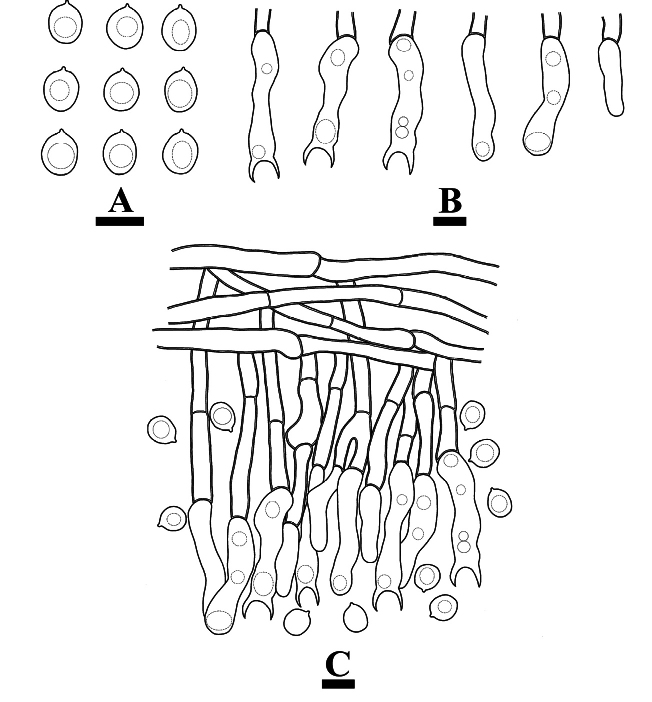 Figure 13
Figure 13 Figure 14
Figure 14 Figure 15
Figure 15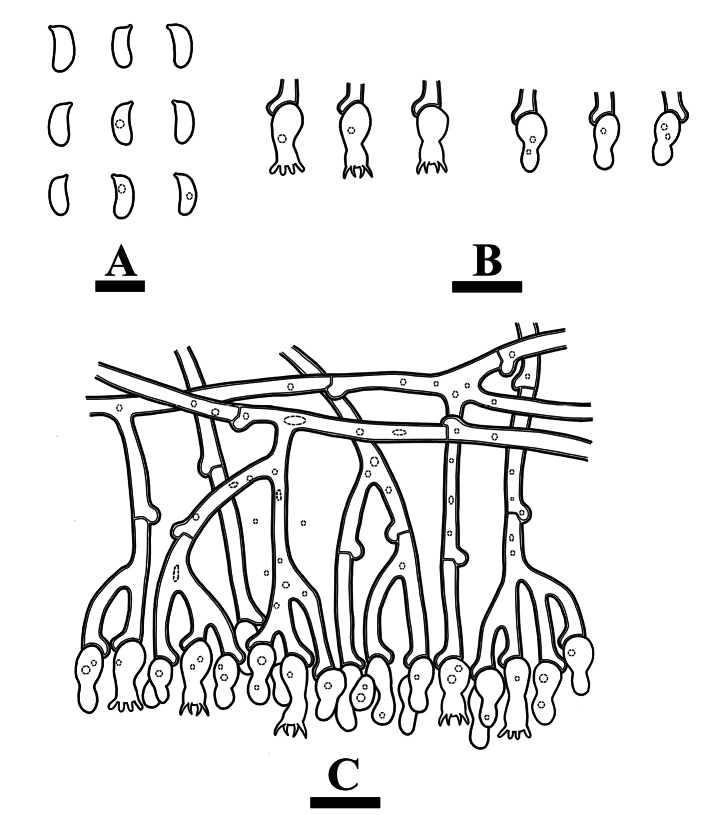 Figure 16
Figure 16 Figure 17
Figure 17| Species name | Specimen No. | GenBank accession No. | Country | References | |
|---|---|---|---|---|---|
| ITS | nLSU | ||||
|
| BR Berger 34240 |
|
| Austria |
|
|
| Hiroshi:Bryoclavula4 |
|
| Japan | NCBI Database |
|
| S-287-FB3 |
| — | Japan |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Flakus 23513 | — |
| USA |
|
|
| Buck 38682 | — |
| Bolivia |
|
|
| JL192-01 SV1 |
| — | USA |
|
|
| JL192-01 SV2 |
| — | USA |
|
|
| JL192-01 SV3 |
| — | USA |
|
|
| JL192-01 SV4 |
| — | USA |
|
|
| Etayo 27623 |
|
| Bolivia |
|
| WS34_1_2_A_As_10000 |
| — | Japan | Unpublished | |
| HHB-19354 |
| — | New Zealand | Unpublished | |
| HHB-19352 |
| — | New Zealand | Unpublished | |
|
| CBS 130.38 |
| — | Japan |
|
|
|
|
| — |
|
|
|
| CBS 131.38 |
| — | Italy |
|
|
|
|
|
|
|
|
|
| EL95_97 | — |
| Sweden |
|
|
|
|
| Cyprus |
| |
|
| B30912949 |
|
| China |
|
|
| B30912949 |
|
| China |
|
|
| FCME 27650 |
|
| Mexico |
|
|
| FCME 27657 |
|
| Mexico |
|
|
| TENN065723 |
| — | USA |
|
|
| PDD:89881 |
| — | New Zealand |
|
|
| TENN043395 |
| — | USA |
|
|
| TN43395 |
|
| New Zealand |
|
|
| FCME 27661 |
|
| Mexico |
|
|
| MEXU 28222 |
|
| Mexico |
|
|
| MB11-6024/2 | — |
| Thailand |
|
|
| AFTOL-ID 471 |
|
| USA |
|
|
| MB18-6024/1 | — |
| Panama |
|
|
| CBS 120605 |
|
| USA |
|
|
| CBS:113.16 | — |
| USA |
|
|
| SH-Ecto-3 | — |
| China | NCBI Database |
|
| KaiR699 |
|
| Japan |
|
|
| Lutzoni 930804-2 |
|
| USA |
|
|
| AFTOL-ID 1130 |
|
| Switzerland |
|
|
| NBRC 114399 |
|
| USA |
|
|
| Lutzoni 930806-1 |
|
| USA |
|
|
| JL596-16 |
|
| Vietnam |
|
|
| EZ4455 |
|
| Canada |
|
|
| AFTOL-ID 710 |
|
| USA |
|
|
| LE-BIN 3507 |
| — | Poland |
|
|
| FCUG 298 | — |
| Canada |
|
|
| AFTOL-ID 613 |
|
| Canada |
|
|
| FCUG 700 | — |
| Sweden |
|
|
| GB700 |
|
| Sweden |
|
|
| TAA167982 | — |
| Canada |
|
|
| TAA180259 | — |
| Sweden |
|
|
| — |
| Canada |
| |
|
| FCUG 701 | — |
| Japan |
|
|
| NH11412 | — |
| Sweden |
|
|
| FCUG 782 | — |
| Sweden |
|
|
| TUMH 64399 |
|
| Sweden |
|
|
|
|
| Sweden |
| |
|
| GB-BN-2 | — |
| Canada |
|
|
| KH Larsson s.n. |
|
| Netherlands |
|
|
| NH7598 | — |
| USA |
|
|
| FCUG 1175 | — |
| Canada |
|
|
| CBS H-21517 |
| — | Canada | NCBI Database |
|
| Thorn429 | — |
| Finland |
|
|
| CBS:531.91 |
|
| Japan |
|
|
| FCUG 659 | — |
| Japan |
|
|
| HK23176 |
|
| Australia |
|
|
| TUMH:64401 |
|
| Finland |
|
|
| TUMH:64402 |
|
| Sweden |
|
|
| CBS 394.63 |
|
| Finland |
|
|
| H HK23176 |
| — | Sweden |
|
|
| KHL8791 | — |
| Canada |
|
|
| KHL 11721 | — |
| USA |
|
|
| KHL 14077 | — |
| Spain |
|
|
| FCUG 2822 | — |
| USA |
|
|
| CBS:126038 |
|
| Finland |
|
|
| EL 28/10 |
|
| Canada |
|
|
| AFTOL-ID 619 | — |
| Sweden |
|
|
| LR 44004 |
|
| USA |
|
|
| FCUG 2188 | — |
| China |
|
|
| GB-BN-4 | — |
| China |
|
|
| PUL: |
| — | China | NCBI Database |
|
|
|
|
|
|
|
|
| Dai 12577 |
|
| China |
|
|
| Dai 12578 | — |
| Sweden |
|
|
| CLZhao 7357 |
|
| USA |
|
|
| CLZhao 7395 |
|
| UK |
|
|
| NH11412 | — |
| Thailand |
|
|
| CBS 126048 |
|
| USA |
|
|
| HFRG |
| — | Panama | NCBI Database |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Plant Pathogens and Fungal Diseases · Forest Ecology and Biodiversity Studies
Introduction
Fungi constitute an integral and valuable part of our natural ecosystem and play an essential ecological role in driving carbon cycling in forest soils, mediating mineral nutrition of plants, and alleviating carbon limitations (Chen et al. 2023a, b; Niego et al. 2023; Dong et al. 2024a; Yang et al. 2025). The wood-inhabiting fungal family Hydnaceae Chevall includes many variations of the fruiting body types within the order Cantharellales J. Schröt (Cai and Zhao 2023; Gao et al. 2024), in which it comprises many representative wood-inhabiting fungi taxa, such as bulbil-shaped, hypochnoid, corticioid, odontoid, poroid, clavarioid, ramarioid, mucronelloid, cantharelloid, and hydnoid basidiomes with diverse hymenophoral and cystidial morphology (Uehling et al. 2012; Diederich et al. 2014; Gruhn et al. 2017; Masumoto and Degawa (2020); Lawrey et al. 2020; Bondartseva and Zmitrovich 2023; Cai and Zhao 2023; Gao et al. 2024).
The genus Burgella Diederich & Lawrey (Hydnaceae, Cantharellales), typified by B.flavoparmeliae Diederich & Lawrey, is characterized by the following features: conidia cylindrical, conidiophores short, hyphae hyaline, septate, straight, rarely branched or anastomosed. Agglomerations of bulbils gelatinous in appearance, almost coralloid, composed of irregularly shaped bulbils; bulbils externally and internally composed of irregular, roundish or elongate cells with clamped septa (Diederich and Lawrey 2007). Based on the MycoBank database (http://www.mycobank.org, accessed on 30 March 2025) and the Index Fungorum (http://www.indexfungorum.org, accessed on 30 March 2025), the genus Burgella has 2 registered species and intraspecies names (Diederich and Lawrey 2007). Previous studies have shown that B.flavoparmeliae and Sistotremaoblongisporum M.P. Christ. & Hauerslev were the sister group of the genus Multiclavula (Diederich and Lawrey 2007). The species B.flavoparmeliae was only distantly related to the type species of the genus Burgoa Goid., which appeared in a different place in the order Cantharellales, and the research revealed that B.flavoparmeliae should not be included in Burgoa, but instead placed in the new genus Burgella (Diederich and Lawrey 2007).
The genus Burgoa, typified by B.verzuoliana Goid. (Hydnaceae, Cantharellales, Agaricomycotina), was established by Goidŕnich to accommodate microfungi producing multicellular spore-like structures with differentiated peridial and internal cells, i.e. bulbils. Apart from the production of the bulbils, members of this genus were distinguished by the formation of clamp connections on their mycelium. This feature showed their affinity to members of Agaricomycotina, but their position within the order Cantharellales was recognised only recently (Diederich and Lawrey 2007; Koukol and Kubátová 2015; Kiyuna et al. 2015). The genus Burgoa is a peculiar microscopic basidiomycete not forming any basidiocarps in its life cycle. So far, this saprotroph has sporadically been found mainly on different woody substrates but the overall knowledge of its ecology and distribution remains sparse due to its rarity (Koukol and Kubátová 2015). Based on the MycoBank database (http://www.mycobank.org, accessed on 30 March 2025) and the Index Fungorum (http://www.indexfungorum.org, accessed on 30 March 2025), the genus Burgoa has 10 registered species and intraspecies names (Diederich and Lawrey 2007; Koukol and Kubátová 2015; Kiyuna et al. 2015).
Clavulina J. Schröt. (Hydnaceae, Cantharellales), with Clavulinacristata (Holmsk.) J. Schröt. as its type species, was established in 1888 (Schröter 1888; He et al. 2019). In China, 14 Clavulina species have been reported on the basis of morphological and molecular analyses, most of which are found in subtropical regions (Gao et al. 2024). In the present study, the new genus Clavuliella falls within Hydnaceae (Cantharellales) and is closely related to Clavulina.
The genus Sistotrema Fr. (Hydnaceae, Cantharellales, Agaricomycetes, Agaricomycotina, Basidiomycota), typified by S.confluens Pers., is a comparatively large genus belonging to the phylum Basidiomycota and is morphologically characterized by resupinate or pileate-stipitate, soft basidiomes, smooth, grandinioid, hydnoid, or poroid hymenophore with various characteristic textures (pellicular, membranaceous, or ceraceous), a monomitic hyphal system with oily inclusions, urniform basidia, and smooth, thin-walled, basidiospores containing cytoplasmic oil droplets (Eriksson et al. 1984; Bernicchia and Gorjón 2010; Cai and Zhao 2023). Based on the MycoBank database (http://www.mycobank.org, accessed on 30 March 2025) and the Index Fungorum (http://www.indexfungorum.org, accessed 30 March 2025), the genus Sistotrema has 224 registered species and intraspecies names, however the actual number of recognized species is 111 (Eriksson et al. 1984; Bernicchia and Gorjón 2010; Sugawara et al. 2022; Cai and Zhao 2023).
In the present study, extensive morphological examinations, combined with analyses of multi-gene sequences data, support the introduction of a new genus and 4 new species of wood-inhabiting fungi. Descriptions and illustrations based on morphological characteristics are presented along with evidence from phylogenetic analyses.
Materials and methods
Morphology
Fresh basidiomata of the fungi growing on angiosperm branches were collected from the Dali, Dehong, and Zhaotong of Yunnan Province, and Guiyang of Guizhou Province, P.R. China. Specimens were dried in an electric food dehydrator at 40 °C (Dong et al. 2024b) then sealed and stored in an envelope and deposited in the Herbarium of the Southwest Forestry University (SWFC), Kunming, Yunnan Province, P.R. China. Macromorphological descriptions were based on field notes and photos captured in the field and lab. Colour terminology followed Petersen (1996). Micromorphological data were obtained from the dried specimens when observed under a light microscope following the previous study (Cai and Zhao 2023). The following abbreviations are used for the micro characteristic description: KOH = 5% potassium hydroxide water solution, CB = Cotton Blue, CB– = acyanophilous, IKI = Melzer’s Reagent, IKI– = both inamyloid and indextrinoid, L = mean spore length (arithmetic average for all spores), W = mean spore width (arithmetic average for all spores), Q = variation in the L/W ratios between the specimens studied and n = a/b (number of spores (a) measured from given number (b) of specimens).
Molecular phylogeny
The EZNA HP Fungal DNA Kit (Omega Biotechnologies Co., Ltd., Kunming, China) was used to extract DNA with some modifications from the dried specimens. The nuclear ribosomal ITS region was amplified with primers ITS5 and ITS4 (White et al. 1990). The PCR procedure for ITS was as follows: initial denaturation at 95 °C for 3 min, followed by 35 cycles at 94 °C for 40 s, 58 °C for 45 s and 72 °C for 1 min, with a final extension of 72 °C for 10 mins. The nuclear nLSU region was amplified with primer pair LR0R and LR7 (Rehner and Samuels 1994). The PCR procedure for nLSU was as follows: initial denaturation at 94 °C for 1 min, followed by 35 cycles at 94 °C for 30 s, 48 °C for 1 min and 72 °C for 1.5 mins with a final extension of 72 °C for 10 mins. The PCR procedure for ITS and nLSU followed a previous study (Zhao and Wu 2017). All newly generated sequences were deposited in NCBI GenBank (https://www.ncbi.nlm.nih.gov/genbank/) (Table 1).
The sequences were aligned in MAFFT version 7 (Katoh et al. 2019) using the G-INS-i strategy. The alignment was adjusted manually using AliView version 1.27 (Larsson 2014). The sequence alignments were deposited in TreeBase (https://treebase.org/treebase-web/home.html;jsessionid=4359D218F4D60336C2A9F7EB7D135CCD) (ID 32177 (Fig. 1)). The sequence alignments were deposited in TreeBase (https://treebase.org/treebase-web/home.html;jsessionid=4359D218F4D60336C2A9F7EB7D135CCD) (ID 32178 (Fig. 2)). Sequences of Platygloeadisciformis (Fr.) Neuhoff retrieved from GenBank were used as the outgroup in the ITS+nLSU analysis (Figs 1, 2; Sugawara et al. 2022).
Maximum Parsimony strict consensus tree illustrating the phylogeny of four new species and a new genus within Hydnaceae, based on ITS+nLSU sequences. Branches are labelled with Maximum Likelihood bootstrap values ≥ 70%, parsimony bootstrap values ≥ 50% and Bayesian posterior probabilities ≥ 0.95, respectively.
Maximum parsimony strict consensus tree illustrating the phylogeny of the one new species and related species in Sistotrema, based on ITS+nLSU sequences. Branches are labelled with Maximum Likelihood bootstrap values > 70%, parsimony bootstrap values > 50% and Bayesian posterior probabilities > 0.95, respectively.
Maximum Parsimony (MP), Maximum Likelihood (ML), and Bayesian Inference (BI) analyses were applied to the combined three datasets following a previous study (Zhao and Wu 2017). All characters were equally weighted and gaps were treated as missing data. Trees were inferred using the heuristic search option with TBR branch swapping and 1,000 random sequence additions. Max trees were set to 5,000, branches of zero length were collapsed, and all parsimonious trees were saved. Clade robustness was assessed using bootstrap (BT) analysis with 1,000 pseudo replicates (Felsenstein 1985). Descriptive tree statistics - tree length (TL), composite consistency index (CI), composite retention index (RI), composite rescaled consistency index (RC) and composite homoplasy index (HI) – were calculated for each maximum parsimonious tree generated. The combined dataset was also analysed using Maximum Likelihood (ML) in RAxML-HPC2 through the CIPRES Science Gateway (Miller et al. 2012). Branch support (BS) for the ML analysis was determined by 1,000 bootstrap pseudo replicates.
MrModelTest 2.3 (Nylander 2004) was used to determine the best-fit evolution model for each dataset for the purposes of Bayesian Inference (BI) which was performed using MrBayes 3.2.7a with a GTR+I+G model of DNA substitution and a gamma distribution rate variation across sites (Ronquist et al. 2012). A total of four Markov chains were run for two runs from random starting trees for 2 million generations for ITS+nLSU (Fig. 1) and 2 million generations for ITS+nLSU (Fig. 2) with trees and parameters sampled every 1,000 generations. The first quarter of all the generations were discarded as burn-in. A majority rule consensus tree was computed from the remaining trees. Branches were considered as significantly supported if they received a Maximum Likelihood bootstrap support value (BS) of ≥ 70%, a maximum parsimony bootstrap support value (BT) of ≥ 70%, or a Bayesian posterior probability (BPP) of ≥ 0.95.
Results
Molecular phylogeny
Burgellaalbofarinacea BLASTN homology search using the ITS nucleotide sequence indicated that the sequence had 87% identity with the sequence as OR471304, named Burgellaflavoparmeliae from the NCBI culture collection (551/635 bp); the nLSU sequence had 98% identity with the sequence as DQ915469, named B.flavoparmeliae from the NCBI culture collection (1294/1323 bp). Burgellafissurata BLASTN homology search using the ITS nucleotide sequence indicated that the sequence had 88% identity with the sequence as OR471304, named B.flavoparmeliae from the NCBI culture collection (553/627 bp); the nLSU sequence had 98% identity with the sequence as DQ915469, named B.flavoparmeliae from the NCBI culture collection (1290/1319 bp). Burgoawumengshanensis BLASTN homology search using the ITS nucleotide sequence indicated that the sequence had 83% identity with the sequence as AB972780, named Burgellaflavoparmeliae from the CBS culture collection (532/643 bp). Clavuliellasinensis BLASTN homology search using the ITS nucleotide sequence indicated that the sequence had 88% identity with the sequence as MT196962, named Clavulinacastaneipes (G.F. Atk.) Corner. from the NCBI culture collection (602/688 bp); the nLSU sequence had 96% identity with the sequence as OM942737, named C.iris from the NCBI culture collection (1281/1338 bp). Sistotremasinense BLASTN homology search using the ITS nucleotide sequence indicated that the sequence had 81% identity with the sequence as OM100765, named S.coroniferum (Höhn. & Litsch.) Donk from the NCBI culture collection (541/669 bp); the nLSU sequence had 99% identity with the sequence as OR460882, named S.brinkmannii (Bres.) J. Erikss. from the NCBI culture collection (1324/1340 bp).
The ITS+nLSU dataset (Fig. 1) comprised sequences from 55 fungal specimens representing 73 taxa. The dataset had an aligned length of 3499 characters, of which 2000 characters were constant, 546 were variable and parsimony-uninformative, and 953 were parsimony-informative. Maximum parsimony analysis yielded 6 equally parsimonious trees (TL = 5747, CI = 0.4047, HI = 0.5953, RI = 0.5604 and RC = 0.2268). The best model of nucleotide evolution for the ITS+nLSU dataset estimated and applied in the Bayesian analysis was found to be GTR+I+G. Bayesian analysis and ML analysis resulted in a similar topology as in the MP analysis. The Bayesian analysis had an average standard deviation of split frequencies = 0.207165 (BI) and the effective sample size (ESS) across the two runs is double the average ESS (avg. ESS) = 362.5. The phylogram, based on the ITS+nLSU rDNA gene regions (Fig. 1), included five genera, viz. Burgella, Burgoa, Clavuliella and Sistotrema, within the family Hydnaceae (Cantharellales, Agaricomycetes). The phylogenetic tree (Fig. 1) inferred from the ITS and nLSU sequences highlighted that the two new species were grouped into the genus Burgella, in which B.albofarinacea was closely related to B.flavoparmeliae, and B.fissurata was grouped with B.lutea Diederich, Capdet, A.I. Romero & Etayo. The phylogram based on the ITS and nLSU data (Fig. 1) showed that Burgoawumengshanensis was clustered into the genus Burgoa, in which it was closely related to B.anomala (Hotson) Goid. and B.verzuoliana Goid. The phylogram based on the ITS+nLSU rDNA gene regions (Fig. 1), included one new species, viz. Sistotremasinense, in which it was grouped into the genus Sistotrema.
The ITS+nLSU dataset (Fig. 2) comprised sequences from 30 fungal specimens representing 40 taxa. The dataset had an aligned length of 2031 characters, of which 1085 characters were constant, 365 were variable and parsimony-uninformative, and 581 were parsimony-informative. Maximum parsimony analysis yielded 12 equally parsimonious trees (TL = 2690, CI = 0.5487, HI = 0.4513, RI = 0.5550 and RC = 0.3045). The best model of nucleotide evolution for the ITS+nLSU dataset estimated and applied in the Bayesian analysis was found to be GTR+I+G. Bayesian analysis and ML analysis resulted in a similar topology as in the MP analysis. The Bayesian analysis had an average standard deviation of split frequencies = 0.005023 (BI) and the effective sample size (ESS) across the two runs is double the average ESS (avg. ESS) = 813.5. The phylogenetic tree (Fig. 2), inferred from the ITS+nLSU sequences, highlighted that Sistotremasinense was grouped closely with S.brinkmannii (Bres.) J. Erikss. and S.farinaceum Hallenb.
Taxonomy
Burgella
albofarinacea
Taxon classificationFungiCantharellalesHydnaceae
Q. Zhou & C.L. Zhao sp. nov.
6A972891-96CF-54B2-8095-7E6FA2D87CF4
857296
Holotype.
China • Yunnan Province, Zhaotong, Yiliang County, Longhai Town, Jianfeng mountain, GPS coordinates: 27°76′N, 104°37′E, altitude: 1777 m asl., on the fallen branch of angiosperm, leg. C.L. Zhao, 26 August 2023, CL Zhao 31820 (SWFC).
Etymology.
albofarinacea (Lat.): refers to the albicans and farinaceous hymenophore of the type specimens.
Description.
Basidiomata annual, resupinate, adnate, pellicular, coriaceous, without odor or taste when fresh, up to 11.5 cm long, 2 cm wide, 50–100 µm thick. Hymenial surface smooth, farinaceous, white when fresh and drying, cracked. Sterile margin thin, white, thinning out, up to 1 mm wide.
Basidiomata of Burgellaalbofarinacea (holotype). Scale bars: 1 cm (A); 1 mm (B).
Hyphal system monomitic, generative hyphae with clamp connections, sometimes with oily contents, colorless, slightly thick-walled, frequently branched, interwoven, 3.5–5.5 μm in diameter; IKI–, CB–, tissues unchanged in KOH.
Microscopic structures of Burgellaalbofarinacea (holotype): basidiospores (A), basidia and basidioles (B), a section of hymenium (C). Scale bars: 5 µm (A); 10 µm (B, C).
Cystidia and cystidioles absent. Basidia suburniform to urniform, slightly thick-walled, with 8 sterigmata and a basal clamp connection, 10.5–22 × 3.5–7 μm; basidioles abundant, in shape similar to basidia, but slightly smaller.
Microscopic structures of Burgellaalbofarinacea (holotype): basidiospores (A), basidia (B), basidioles (C), a section of hymenium (D), generative hyphae (E, F). Scale bars: 10 µm (A–F).
Basidiospores ellipsoid to allantoid, colorless, thin-walled, smooth, with oil drop, IKI–, CB–, (4–)4.5–6.5(–7) × 2–4 μm, L = 5.34 µm, W = 2.83 µm, Q = 1.79–1.97 μm (n = 120/4).
Additional specimens examined (paratypes).
China • Yunnan Province, Zhaotong, Yiliang County, Longan Town, GPS coordinates: 27°73′N, 104°16′E, altitude: 1550 m asl., on the fallen branch of angiosperm, leg. C.L. Zhao, 27 August 2023, CLZhao 32026, CLZhao 31855 (SWFC) • Zhaotong, Yiliang County, Xiaocaoba Town, GPS coordinates: 27°26′N, 104°26′E, altitude: 2225 m asl., on the fallen branch of angiosperm, leg. C.L. Zhao, 28 August 2023, CLZhao 32468 (SWFC).
Burgella
fissurata
Taxon classificationFungiCantharellalesHydnaceae
Q. Zhou & C.L. Zhao sp. nov.
F2CDDF76-4168-598C-B5CB-B26A636EE51A
857297
Holotype.
China • Yunnan Province: Dehong, Yingjiang County, Tongbiguan provincial nature reserve, GPS coordinates: 24°30′N, 097°30′E, altitude: 1300 m asl., on the fallen branch of angiosperm, leg. C.L. Zhao, 19 July 2023, CLZhao 30212 (SWFC).
Etymology.
fissurata (Lat.): refers to the cracking hymenial surface of the type specimens.
Description.
Basidiomata annual, resupinate, adnate, pruinose, hypochnoid, without odor or taste when fresh, up to 10.2 cm long, 1 cm wide, 50–100 µm thick. Hymenial surface smooth, cracked, white when fresh, turning to pale cream upon drying. Sterile margin thin, white, thinning out, up to 1 mm wide.
Basidiomata of Burgellafissurata (holotype). Scale bars: 1 cm (A); 1 mm (B).
Hyphal system monomitic, generative hyphae with clamp connections, colorless, thin-walled, frequently branched, interwoven, 2–4 μm in diameter; IKI–, CB–, tissues unchanged in KOH.
Microscopic structures of Burgellafissurata (holotype): basidiospores (A), basidia and basidioles (B), umbrella-shaped cystidia (C), a section of hymenium (D). Scale bars: 5 µm (A); 10 µm (B, C).
Cystidia umbrella-shaped, colorless, thin-walled, smooth, 5.5–10 × 4–6 µm; cystidioles absent. Basidia urniform, with a median constriction, thin-walled, with 4 sterigmata and a basal clamp connection, 6–11.5 × 2–4.5 μm; basidioles abundant, in shape similar to basidia, but slightly smaller.
Microscopic structures of Burgellafissurata (holotype): basidiospores (A), basidia and basidioles; umbrella-shaped cystidia (B), generative hyphae (C), a section of hymenium (D). Scale bars: 10 µm (A–D).
Basidiospores narrowly ellipsoid, colorless, thin-walled, smooth, IKI–, CB–, (2.5–)3–4 × 1–2 μm, L = 3.36 µm, W = 1.63 µm, Q = 2.06 μm (n = 30/1).
Burgoa
wumengshanensis
Taxon classificationFungiCantharellalesHydnaceae
Q. Zhou & C.L. Zhao sp. nov.
A66E80E0-33C3-56B0-B936-F0A92C534D8C
857298
Holotype.
China • Yunnan Province: Zhaotong, Yiliang County, Luozehe Town, Lijiaping Village, Wumengshan National Nature Reserve, GPS coordinates: 27°29′N, 103°55′E, altitude: 1900 m asl., on the fallen branch of angiosperm, leg. C.L. Zhao, 19 September 2023, CLZhao 33227 (SWFC).
Etymology.
wumengshanensis (Lat.): refers to the locality, Wumengshan National Natural Reserve, of the type specimens.
Basidiomata of Burgoawumengshanensis (holotype). Scale bars: 1 cm (A); 1 mm (B)
Description.
Basidiomata annual, resupinate, adnate, pellicular, pruinose upon drying, without odor or taste when fresh, up to 7.4 cm long, 2.1 cm wide, 40–90 µm thick. Hymenial surface smooth, white when fresh, turning to pale cream upon drying. Sterile margin thin, white, up to 1 mm wide.
Microscopic structures of Burgoawumengshanensis (holotype) basidiospores (A), basidia and basidioles (B), schizopapillate cystidia (C), clavate cystidioles (D), a section of hymenium (E). Scale bars: 5 µm (A), 10 µm (B, E).
Hyphal system monomitic, generative hyphae with clamp connections, sometimes with oily contents, colorless, slightly thick-walled, frequently branched, interwoven, 3.5–5 µm in diameter; IKI–, CB–, tissues unchanged in KOH.
Microscopic structures of Burgoawumengshanensis (holotype) basidiospores (A), basidia (B), clavate cystidioles (C), schizopapillate cystidia (D), a section of hymenium (E), generative hyphae (F, G). Scale bars: 10 µm (A–G).
Cystidia schizopapillate, colorless, thin-walled, smooth, 30.5–49 × 5.5–8 µm; cystidioles clavate, colorless, thin-walled, smooth, 22–29.5 × 5–8 µm. Basidia urniform, with a median constriction, slightly thick-walled, with 8 sterigmata and a basal clamp connection, 12–20 × 3.5–6 µm; basidioles abundant, in shape similar to basidia, but slightly smaller.
Basidiospores ellipsoid, colorless, thin-walled, smooth, with oil drop, IKI–, CB–, (3.5)–4–5.5(–6) × 2–3.5 µm, L = 4.5 µm, W = 2.78 µm, Q = 1.62 (n = 30/1).
Clavuliella
Taxon classificationFungiCantharellalesHydnaceae
Q. Zhou & C.L. Zhao gen. nov.
DDF9D9F6-E815-53E6-BDDA-1598E2418025
858532
Type species.
Clavuliellasinensis Q. Zhou & C.L. Zhao, sp. nov.
Etymology.
Clavuliella (Lat.): refers to the related genus Clavulina.
Description.
Basidiomata annual, coralloid, gregarious to caespitose clusters, greyish white to light grey when fresh, turning to dark grey upon drying; with sharply acuminate or cristate tips. Hyphal system monomitic, generative hyphae simple-septa, colorless, slightly thick-walled, frequently branched, interwoven. Cystidia and cystidioles absent. Basidia cylindrical, with a median constriction, with 2 sterigmata and a basal simple-septum, with oily contents. Basidiospores subglobose to broadly ellipsoid, colorless, thin-walled, smooth, with oil drop, IKI–, CB–.
Notes.
In our phylogenetical analyses (Fig. 1), Clavuliella was identified as a monophyletic group, typified by C.sinensis. The new genus Clavuliella falls within the family Hydnaceae (Cantharellales) and is closely related to Clavulina. Clavulina is distinguished from Clavuliella by its clavarioid to coralloid, simple or branched basidiomata with amphigenous hymenia, cylindrical to subclavate basidia with two or more cornuted sterigmata (Schröter 1888; Uehling et al. 2012; He et al. 2019; Gao et al. 2024).
Clavuliella resembles Clavulina in sharing coralloid basidiomata, subglobose, thin-walled basidiospores. However, Clavuliella differs from Clavulina by cylindrical basidia, with a median constriction, slightly thick-walled, with 2 sterigmata and a basal simple-septum, with oily contents and simple-septa generative hyphae. In this study, Clavuliella originating from the subtropical regions, suggests the possibility of discovering new corticioid taxa through further investigations and molecular analyses.
Clavuliella
sinensis
Taxon classificationFungiCantharellalesHydnaceae
Q. Zhou & C.L. Zhao sp. nov.
FF1BAA6A-D40A-5FF2-97BD-0F8FA8756525
858533
Holotype.
China • Guizhou Province: Guiyang, Qianlingshan Forest Park, GPS coordinates: 26°36′N, 106°41′E, altitude: 1396 m asl., on the ground, leg. C.L. Zhao, 21 August 2023, CLZhao 31231 (SWFC).
Etymology.
sinensis (Lat.): refers to the type locality (China).
Description.
Basidiomata annual, coralloid, gregarious to caespitose clusters, 0.6–1.5 cm tall, 0.62–1.6 cm wide, frequently branched 2–3 times, forming dichotomous branches at the apices, without odor or taste, soft when fresh, becoming brittle upon drying, usually lacking obvious basal mycelium; greyish white to light grey when fresh, turning to dark grey upon drying; with sharply acuminate or cristate tips.
Basidiomata of Clavuliellasinensis (holotype). Scale bars: 1 cm (A); 1 mm (B).
Hyphal system monomitic, generative hyphae simple-septa, colorless, slightly thick-walled, frequently branched, interwoven, 4–10.5 μm in diameter; IKI–, CB–, tissues unchanged in KOH.
Microscopic structures of Clavuliellasinensis (holotype): basidiospores (A), basidia and basidioles (B), a section of hymenium (C). Scale bars: 5 µm (A); 10 µm (B, C).
Cystidia and cystidioles absent. Basidia cylindrical, with a median constriction, slightly thick-walled, with 2 sterigmata and a basal simple-septum, with oily contents, 18.5–43 × 6–9 μm; basidioles abundant, in shape similar to basidia, but slightly smaller.
Microscopic structures of Clavuliellasinensis (holotype): basidiospores (A), basidia and basidioles (B), a section of hymenium (C), generative hyphae (D). Scale bars: 10 µm (A–D).
Basidiospores subglobose to broadly ellipsoid, colorless, thin-walled, smooth, with oil drop, IKI–, CB–, (7–)7.5–9.5(–10) × 6.5–8(–8.5) μm, L = 8.52 µm, W = 7.51 µm, Q = 1.13 μm (n = 30/1).
Sistotrema
sinense
Taxon classificationFungiCantharellalesHydnaceae
Q. Zhou & C.L. Zhao sp. nov.
D0864714-BA6E-5CEA-861F-C1A84E938B0A
857299
Holotype.
China • Yunnan Province: Dali, Weishan County, Qinghua Town, GPS coordinates: 25°01′N, 100°22′E, altitude: 2071.6 m asl., on the fallen branch of angiosperm, leg. C.L. Zhao, 18 October 2022, CLZhao 24876 (SWFC).
Basidiomata of Sistotremasinense (holotype). Scale bars: 1 cm (A); 1 mm (B).
Etymology.
sinense (Lat.): refers to the type locality (China).
Description.
Basidiomata annual, resupinate, adnate, soft coriaceous when fresh, becoming coriaceous upon drying, without odor or taste when fresh, up to 11 cm long, 2.5 cm wide, 50–100 µm thick. Hymenial surface smooth, white when fresh, turning to white to incanus upon drying. Sterile margin thin, white, thinning out, up to 1 mm wide.
Microscopic structures of Sistotremasinense (holotype): basidiospores (A), basidia and basidioles (B), a section of hymenium (C). Scale bars: 5 µm (A); 10 µm (B, C).
Hyphal system monomitic, generative hyphae with clamp connections, often and characteristically with oil content, colorless, slightly thick-walled, frequently branched, interwoven, 2–4 μm in diameter; IKI–, CB–, tissues unchanged in KOH.
Microscopic structures of Sistotremasinense (holotype): basidiospores (A), a section of hymenium (B). Scale bars: 10 µm (A, B).
Cystidia and cystidioles absent. Basidia suburniform to urniform, slightly thick-walled, with 4 sterigmata and a basal clamp connection, 8–13.5 × 3–5 μm; basidioles abundant, in shape similar to basidia, but slightly smaller.
Basidiospores suballantoid to allantoid, colorless, thin-walled, smooth, IKI–, CB–, 3–4.5(–5) × (1–)1.5–2.5 μm L = 3.8 µm, W = 1.72 µm, Q = 2.21 μm (n = 30/1).
Discussion
In recent years, many taxa of wood-inhabiting fungi have been continuously reported and recorded all over the world, including in the genus Burgella, Burgoa, and Sistotrema (Diederich et al. 2014; Koukol and Kubátová 2015; Cai and Zhao 2023; Dong et al. 2024b; Yuan et al. 2024; Zhang et al. 2024; Yang et al. 2025). Several previous studies, based on ITS+nLSU sequence data, confirmed phylogenetic relationships, in which the genus Burgella, Burgoa, Clavuliella, and Sistotrema are nested in the order Cantharellales (Diederich et al. 2014; Koukol and Kubátová 2015).
Phylogenetically, based on the multiple loci in the family Hydnaceae, four genera; Burgella, Burgoa, Clavuliella, and Sistotrema were located in this family (Zhou and Qin 2013; Vu et al. 2019; Masumoto and Degawa 2020; Sugawara et al. 2022; Campo et al. 2023; Cai and Zhao 2023). In the present study, based on the phylogram inferred from the ITS+nLSU data (Fig. 1), two new species were grouped into the genus Burgella, in which B.albofarinacea was closely related to B.flavoparmeliae and B.fissurata was grouped with B.lutea. The phylogram based on inferences from the ITS and nLSU data (Fig. 1) showed that B.wumengshanensis clustered into the genus Burgoa, in which it was grouped closely with B.anomala and B.verzuoliana. The phylogram based on inferences from the ITS and nLSU data (Fig. 1) showed that Clavuliella was identified as a monophyletic group, typified by C.sinensis, in which it was grouped closely with C.minor X.X. Huang & L.H. Qiu, C.cristata, and C.iris Loizides, Bellanger & P.-A. Moreau. The phylogenetic tree (Fig. 2), inferred from the ITS+nLSU sequences, highlighted that Sistotremasinense was grouped closely with S.brinkmannii and S.farinaceum.
Morphologically, Burgellaflavoparmeliae, B.lutea and B.fissurata are similar to B.albofarinacea by having the generative hyphae with septa or with clamp connections. (Diederich and Lawrey 2007). However, B.flavoparmeliae differs from B.albofarinacea by the irregularly shaped, coralloid, orange agglomerations of bulbils and generative hyphae with septa or without clamp connections, with both morphologies present on the same hyphae at neighboring septa (Diederich and Lawrey 2007). B.lutea is distinguished from B.fissurata by the superficial, yellow to orange-yellow, roundish bulbils (Diederich et al. 2014). B.fissurata is distinguishable from B.albofarinacea by the pruinose hypochnoid hymenial surface, with umbrella-shaped cystidia, basidia with 4 sterigmata, and its smaller basidia (6–11.5 × 2–4.5 μm; Diederich and Lawrey 2007; Koukol and Kubátová 2015; Kiyuna et al. 2015).
Burgellaflavoparmeliae is separated from B.fissurata by the irregularly shaped, coralloid, orange agglomerations of bulbils and generative hyphae with septa or without clamp connections, both situations present on the same hyphae at neighbouring septa (Diederich and Lawrey 2007). B.lutea is distinguished from B.fissurata by the superficial, yellow to orange-yellow, roundish bulbils (Diederich et al. 2014). B.albofarinacea differs from B.fissurata by the pellicular, coriaceous hymenial surface, bigger basidia with 8 sterigmata and bigger basidiospores (Cai and Zhao 2023).
Morphologically, Burgoaanomala and B.verzuoliana are similar to B.wumengshanensis by having the generative hyphae with clamp connections (Koukol and Kubátová 2015). However, B.anomala is distinguishable from B.wumengshanensis by having spherical bulbils, and hyaline to pale brown generative hyphae, thin-walled, thinner (2–5 μm) in diameter (Koukol and Kubátová 2015); B.verzuoliana is distinguished from B.wumengshanensis by having spherical bulbils (Diederich and Lawrey 2007).
Morphologically, Clavulinacristata, C.griseoviolacea Yue Gao, Hao Zhou, & C.L. Hou, and C.pallida Yue Gao, Hao Zhou & C.L. Hou are similar to Clavuliellasinensis by having clavarioid to coralloid basidiomata and guttulate basidiospores (Uehling et al. 2012; Gao et al. 2024). However, Clavulinacristata is separated from Clavuliellasinensis by having cylindrical to subclavate basidia with two or more cornuted sterigmata (Gao et al. 2024); Clavulinagriseoviolacea differs from Clavuliellasinensis by having gray to dark grayish violet basidiomata with a white stipe, hyphae with clamp connections, and smaller basidiospores (6.5–8.0 × 6.2–7.2 μm; Crous et al. 2014); Clavulinapallida is distinguishable from Clavuliellasinensis by having solitary or scattered basidiomata, generative hyphae clamp connections, and longer basidia (34.2–48.5 × 4.8–6.3 μm; Gao et al. 2024).
Morphologically, Sistotremadiademiferum (Bourdot & Galzin) Donk, S.coroniferum (Höhn. & Litsch.) Donk and S.hispanicum M. Dueñas, Ryvarden & Tellería are similar to S.sinense by having the urniform basidia and basal hyphae with clamp connections (Bernicchia and Gorjón 2010). However, S.diademiferum is separated from S.sinense by the smooth, porulose hymenophore, larger basidia with 6 sterigmata (15–20 × 5–7 μm), and ovoid to subglobose basidiospores (Bernicchia and Gorjón 2010). S.coroniferum is distinguishable from S.sinense by the smooth hymenophore, with gloeocystidia, basidia with 6 sterigmata, and longer subcylindrical basidiospores (5–6 × 2–2.5 µm; Bernicchia and Gorjón 2010). S.hispanicum differs from S.sinense by the whitish to yellow hymenial surface and bigger narrowly ellipsoid to subreniform basidiospores (5.5–6 × 3–4 μm; Bernicchia and Gorjón 2010).
As wood-inhabiting fungi efficiently degrade lignocellulose in wood, they play a crucial ecological role in material recycling and energy flow in forest ecosystems, as well as playing a major economic role (Sugawara et al. 2022; Zhang et al. 2022; Bondartseva and Zmitrovich 2023; Campo et al. 2023; Cui et al. 2019; Gao et al. 2024; Liu et al. 2023a, b; Sun et al. 2020; Sun et al. 2022; Ji et al. 2022). Wood-inhabiting fungi are an extensively studied group of Basidiomycota, but their diversity is still not well known in China, and many recently described taxa in this ecological group have been discovered from China (Sugawara et al. 2022; Cai and Zhao 2023; Wang et al. 2023; Wang et al. 2024; Wu et al. 2022; Yuan et al. 2023; Zhao et al. 2024). Four new species and a new genus, from the Yunnan and Guizhou Provinces of China, serve as examples of the understudied fungal diversity present in the P.R. of China.. On a wider scale, this study enriches our knowledge on the diversity of wood-inhabiting fungi worldwide.
Supplementary Material
XML Treatment for Burgella albofarinacea
XML Treatment for Burgella fissurata
XML Treatment for Burgoa wumengshanensis
XML Treatment for Clavuliella
XML Treatment for Clavuliella sinensis
XML Treatment for Sistotrema sinense
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bernicchia A Gorjón SP (2010) Fungi Europaei 12: Corticiaceae s.l.Edizioni Candusso, Alassio, Italy, 1008 pp.
- 2Binder M Hibbett DS (2002) Higher-level phylogenetic relationships of Homobasidiomycetes (mushroom-forming fungi) inferred from four r DNA regions.Molecular Phylogenetics and Evolution 22(1): 76–90. 10.1006/mpev.2001.104311796031 · doi ↗ · pubmed ↗
- 3Binder M Hibbett DS Larsson KH Larsson E Langer E Langer G (2005) The phylogenetic distribution of resupinate forms across the major clades of mushroom-forming fungi (Homobasidiomycetes).Systematics and Biodiversity 3(2): 113–157. 10.1017/S 1477200005001623 · doi ↗
- 4Bondartseva MA Zmitrovich IV (2023) Order Cantharellales: Taxonomic and ecological diversification. Biology Bulletin Reviews 13(S 1): 1–16. 10.1134/S 2079086423070046 · doi ↗
- 5Cai LQ Zhao CL (2023) Molecular phylogeny and morphology reveal a new wood-rotting fungal species, Sistotremayunnanense sp. nov. from the Yunnan-Guizhou Plateau.Mycoscience 64(3): 101–108. 10.47371/mycosci.2023.04.00137397608 PMC 10308066 · doi ↗ · pubmed ↗
- 6Campo E Franchi P Marchetti M (2023) Clavulinaperplexa, una nuova specie trovata in Friuli-Venezia Giulia.66(1): 81–95.
- 7Chen YP Su PW Hyde KD Maharachchikumbura SSN (2023 a) Phylogenomics and diversifcation of Sordariomycetes.Mycosphere : Journal of Fungal Biology 14(1): 414–451. 10.5943/mycosphere/14/1/5 · doi ↗
- 8Chen YP Su PW Stadler M Xiang R Hyde KD Tian WH Maharachchikumbura SSN (2023 b) Beyond observation: Genomic traits and machine learning algorithms for predicting fungal lifestyles.Mycosphere: Journal of Fungal Biology 14(1): 1530–1563. 10.5943/mycosphere/14/1/17 · doi ↗