First karyotype description of Epiplatysspilargyreius (Duméril, 1861) with comments on chromosome evolution in the genus Epiplatys Gill, 1862 (Nothobranchiidae)
Sergey A. Simanovsky, Alexandra Yu. Skuratovskaya, Margarita G. Simonian, Dmitry A. Medvedev, Fekadu Tefera, Alexander S. Golubtsov

TL;DR
This paper describes the karyotype of Epiplatys spilargyreius and discusses chromosome evolution in the genus.
Contribution
The study provides the first karyotype description of Epiplatys spilargyreius and explains chromosome number reduction via Robertsonian translocations.
Findings
Epiplatys spilargyreius has a diploid chromosome number of 2n = 34.
Chromosome arm number (FN) is 52 in both male and female karyotypes.
Robertsonian translocations are suggested as the main cause of 2n reduction in the genus.
Abstract
The African non-annual killifish genus Epiplatys Gill, 1862 (family Nothobranchiidae) comprises 36 valid species distributed in West, Central and East Africa. The available cytogenetic information for the genus indicates a wide variability in diploid chromosome number (2n) and number of chromosome arms (FN). Here, we report the karyotype of Epiplatysspilargyreius (Duméril, 1861), one of the two species with the lowest diploid chromosome number (2n = 34) in the genus, from the White Nile basin in Ethiopia. Male and female karyotypes contained 18 metacentric/acrocentric and 16 subtelocentric/acrocentric chromosomes. The number of chromosome arms is, respectively, FN = 52. Analysis of karyotype differentiation in the genus allowed us to suggest that the 2n reduction in E.spilargyreius and many other members of the genus Epiplatys is mainly due to Robertsonian translocations (reduction…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
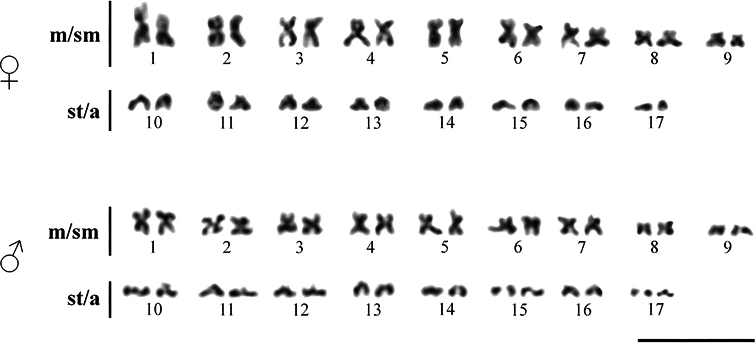 Figure 1
Figure 1| Taxon | 2n | Karyotypic formula |
| Origin | References |
|---|---|---|---|---|---|
| 50 | 20m/sm + 30st/a | 70 | Guinee, Liberia | ||
| 46 | 4m/sm + 42st/a | 50 | Gabon | ||
| 48 | 2m/sm + 46st/a | 50 | Gabon | ||
| 34 | 14m/sm + 20st/a | 48 | Sierra Leone, Liberia | ||
| 40 | 8m + 32a | 48 | Ghana, Nigeria | ||
| 50 | 50st/a | 50 | Ghana | ||
| 48 | – | – | Democratic Republic of the Congo |
| |
| 50 | 32m/sm + 18st/a | 82 | Liberia | ||
| 50 | 36m/sm + 14st/a | 86 | Ghana |
| |
| 48 | – | 48 | Congo | ||
| 42 | 8m + 34a | 50 | Cameroon | ||
| 42 | 8m/sm + 34st/a | 50 | Cameroon |
| |
| 36 | – | – | Liberia, Sierra Leone | ||
| 38 | – | – | |||
| 40 | 10m/sm + 30st/a | 50 | |||
| 48 | 2sm + 46a | 50 | Cameroon, Equatorial Guinea | ||
| 46 | 6m/sm + 40a | 52 | Gabon | ||
| 48 | – | – | Cameroon | ||
| 48 | 2m/sm + 46st/a | 50 | Cameroon (three populations) |
| |
| 48 | 4m/sm + 44a | 52 | Liberia | ||
| 48 | – | – | Guinea |
| |
| 48 | 48a | 48 | Central African Republic | ||
| 46 | 14m/sm + 32st/a | 60 | Zaire, Congo | ||
| 38 | 12m/sm + 26st/a | 50 | Ivory Coast | ||
| 46 | – | – | Liberia | ||
| 48 | – | – | Cameroon | ||
| 48 | 48st/a | 48 | Ghana, Nigeria, Cameroon, Equatorial Guinea | Scheel, 1972, 1975, 1990; | |
| 42 | – | – | Zaire | ||
| 34 | – | – | Nigeria | ||
| 34 | – | – | Democratic Republic of the Congo |
| |
| 34 | 18m/sm + 16st/a | 52 | Ethiopia |
|
- —Russian Science Foundation 501100006769 https://ror.org/03y2gwe85 http://doi.org/10.13039/501100006769
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsChromosomal and Genetic Variations · Fish Biology and Ecology Studies · Parasite Biology and Host Interactions
Introduction
The genus Epiplatys belongs to the family Nothobranchiidae and comprises 36 valid species of non-annual killifish endemic to Africa (Wildekamp 1996; Fricke et al. 2025; Froese and Pauly 2025). Representatives of the genus are distributed in West, Central and East Africa – from Senegal to Ethiopia – in small rivers, streams and swamps in rainforests and savannas (Wildekamp 1996; Froese and Pauly 2025). The genus Epiplatys is reported to have the widest distribution of all Nothobranchiidae (Collier 2015).
Cytogenetic information for the genus, available for 22 of the 36 species, shows a wide variability in diploid chromosome number from 34 to 50 and the number of chromosome arms from 48 to 82 (Scheel 1966, 1972, 1975, 1990; Arai 2011; Collier et al. 2009; Collier 2015). Analysis of cytogenetic and phylogenetic data suggests that the ancestral diploid chromosome number was 2n = 48 and that karyotype differentiation in the genus occurred mainly by chromosomal fusions and pericentric inversions, and less frequently by chromosomal fissions (Collier et al. 2009; Collier 2015). In this paper, we describe the karyotype of Epiplatysspilargyreius (Duméril, 1861), one of the two species with the lowest 2n in the genus, from the White Nile Basin in Ethiopia (East Africa), to further our understanding of chromosome evolution in the genus. Previously, E.spilargyreius has been cytogenetically studied from Nigeria and the Democratic Republic of the Congo (West and Central Africa respectively), but only 2n has been reported (Scheel 1966, 1972, 1990; Collier 2015).
Material and methods
Four individuals, two females and two males of E.spilargyreius (standard length SL = 21–28 mm), were obtained from an oxbow pond at the right bank of the Baro River, a tributary of the Sobat River, the White Nile system, at the Village of Baziel Kebele (08°18'51.2"N, 34°04'24.9"E), in southwestern Ethiopia. Fish were collected by the Joint Ethio-Russian Biological Expedition (JERBE) with the permission of the National Fisheries and Aquatic Life Research Center under the Ethiopian Institute of Agricultural Research (EIAR) and the Ethiopian Ministry of Science and Technology. Three individuals, two females and one male, were successfully karyotyped.
After colchicine treatment, fish were euthanized with an overdose of tricaine methanesulfonate (MS-222), identified, measured with an accuracy of 1 mm, dissected for gonad examination and tissue sampling, and preserved in 70% ethanol. Species identification was done based on morphological characters (Golubtsov et al. 1995). The experiments were carried out in accordance with the rules of the Severtsov Institute of Ecology and Evolution (IEE). Vouchers are deposited at the Severtsov Institute of Ecology and Evolution (Moscow), under provisional labels of JERBE.
Before preparation, fish were treated intraperitoneally with 0.1% colchicine (0.01 ml / 1 g of their weight) for 3–5 hours. After euthanasia, chromosome preparations were obtained from kidney tissue following Kligerman and Bloom (1977) with some modifications as described in Simanovsky et al. (2022). The chromosome spreads were stained conventionally with 4% Giemsa solution in a phosphate buffer solution at pH 6.8 for 8 min. The analysis was performed using an Axioplan 2 Imaging microscope (Carl Zeiss, Germany) equipped with a CV-M4+CL camera (JAI, Japan) and Ikaros software (MetaSystems, Germany). Final images were processed using Photoshop software (Adobe, USA). Karyotypes were arranged according to the centromere position following the nomenclature of Levan et al. (1964), but modified as metacentric/submetacentric (m/sm) and subtelocentric/acrocentric (st/a), similar to other cytogenetic studies on the members of the genus Epiplatys (Table 1). Chromosome pairs were arranged according to their size in each chromosome category. To determine the chromosomal arm number per karyotype (fundamental number, FN), metacentrics/submetacentrics were considered as biarmed, and subtelocentrics/acrocentrics as one arm chromosomes. The total number of complete metaphases studied for three E.spilargyreius individuals was 30.
Results and discussion
The karyotype of the E.spilargyreius population from Ethiopia has 2n = 34 and consists of 18 metacentric/submetacentric and 16 subtelocentric/acrocentric chromosomes, FN = 52 (Fig. 1). No distinguishable sex chromosomes were observed in the female and male complements, similar to the previous cytogenetic studies on the representatives of the genus Epiplatys (Scheel 1966, 1972, 1975, 1990; Arai 2011; Collier et al. 2009; Collier 2015). The three studied populations of E.spilargyreius from Nigeria, the Democratic Republic of the Congo and Ethiopia have identical chromosome numbers (Scheel 1966, 1972, 1990; Collier 2015; this work). We could not perform a more detailed comparative analysis of karyotypes in these populations because only 2n has been reported for populations from Nigeria and the Democratic Republic of the Congo (see Table 1). Analysis of mitochondrial genes also revealed a close similarity between populations, despite the wide distribution of E.spilargyreius (Collier 2015). The cytogenetic characteristics (2n, FN) of the studied members of the genus Epiplatys are shown in Table 1.
Female and male karyotypes of Epiplatysspilargyreius after conventional Giemsa staining. Scale bar: 10 μm.
According to phylogenetic data, the genus is divided into two major groups: “western and savanna” and “eastern and coastal” clades (Collier et al. 2009; Collier 2015). E.spilargyreius belongs to the first clade, which also includes E.bifasciatus (2n = 40, FN = 48), E.duboisi (2n = 48), E.fasciolatus (2n = 36–40, FN = 50), E.lamottei (2n = 48, FN = 52) and E.roloffi (2n = 46) among the karyotyped species. The diploid chromosome number in this clade varies widely from 34 to 48, but FN is more stable and varies only from 48 to 52. This suggests a substantial role for structural rearrangements of centric fusion type (Robertsonian translocations) in the karyotype differentiation. It is worth noting that E.duboisi and E.lamottei, which have cytogenetic characteristics closest to the putative ancestral karyotype of the genus with 2n = 48 and FN = 48, are at the basal position on the phylogenetic tree of the clade (Collier et al. 2009; Collier 2015). Karyotypes of E.bifasciatus, E.fasciolatus and E.spilargyreius differ from the putative ancestral karyotype by 4, 4–6 and 7 centric chromosome fusions, respectively.
The second, “eastern and coastal” clade, includes 11 karyotyped representatives. Five species – E.grahami, E.infrafasciatus, E.sangmelinensis, E.mesogramma and E.chevalieri – share cytogenetic characteristics close to the putative ancestral state, occupying different positions on the phylogenetic tree. In E.esekanus (2n = 42; FN = 50), E.huberi (2n = 46, FN = 52) and E.singa (2n = 42) chromosome number is independently reduced presumably by centric fusions, similar to members of the first clade. The closely related E.chaperi, E.dageti and E.annulatus have 2n = 50. It can be suggested that these karyotypes were derived by tandem fissions. It is noteworthy that in E.annulatus (FN = 70) and E.dageti (FN = 82/86) the number of chromosome arms is greatly increased, probably due to pericentric inversions or other types of centromere repositioning.
Among the karyotyped species for which the position on the phylogenetic tree is not known, it is important to mention E.ansorgii (2n = 46/48, FN = 50), E.barmoiensis (2n = 34, FN = 48), E.multifasciatus (2n = 46, FN = 60) and E.olbrechtsi (2n = 38, FN = 50), in which 2n is reduced probably due to centric chromosome fusions, similar to the representatives of the first and second clades. The karyotype of E.multifasciatus also has a markedly increased number of biarmed elements. E.sexfasciatus (2n = 48, FN = 48), a species also of an unknown phylogenetic position, has cytogenetic characteristics close to the putative ancestral state.
To summarize, chromosome evolution in the genus Epiplatys is characterized by the following: 1) the putative ancestral karyotype with 2n = 48 and FN = 48; 2) independent events of structural rearrangements of centric fusion type; 3) increase in the number of chromosomal arms by pericentric inversions or other types of centromere repositioning, and 4) at least one chromosome fission event. The cytogenetic study in the genus Epiplatys is interesting in the context of the study of the family Nothobranchiidae, which is characterized by high rates of chromosome and genome evolution, with maximum expression in African annual killifishes of the genus Nothobranchius Peters, 1868 (Krysanov, Demidova 2018; Cui et al. 2019; Štundlová et al. 2022; Krysanov et al. 2023; Lukšíková et al. 2023; Voleníková et al. 2023). Based on the phylogenetic tree (Cui et al. 2019), it can be assumed that the genus Epiplatys shares more basal features of karyotype differentiation compared to the genus Nothobranchius.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arai R (2011) Fish karyotypes – a Check List. Springer, 340 pp. 10.1007/978-4-431-53877-6 · doi ↗
- 2Collier GE (2015) Molecular phylogeny and inferred path of dispersal of the genus Epiplatys.Journal of the American Killifish Association 48(5–6): 131–149.
- 3Collier GE Murphy WJ Espinoza M (2009) Phylogeography of the genus Epiplatys (Aplocheiloidea: Cyprinodontiformes).Molecular Phylogenetics and Evolution 50: 190–196. 10.1016/j.ympev.2008.10.00618984059 · doi ↗ · pubmed ↗
- 4Cui R Medeiros T Willemsen D Iasi LNM Collier GE Graef M Reichard M Valenzano DR (2019) Relaxed selection limits lifespan by increasing mutation load.Cell 178: 385–399. [e 20] 10.1016/j.cell.2019.06.00431257025 · doi ↗ · pubmed ↗
- 5Fricke R Eschmeyer W Nvan der Laan R (2025) Catalog of Fishes: Genera, Species, References. [accesed 22.02.2025]
- 6Froese R Pauly D (2025) Fish Base. http://www.fishbase.org [accessed 22.02.2025]
- 7Golubtsov AS Darkov AA Dgebuadze YY Mina MV (1995) An artificial key to fish species of the Gambela region (the White Nile basin in the limits of Ethiopia).Joint Ethio-Russian Biological Expedition, Addis Abeba, 84 pp.
- 8Kligerman AD Bloom SE (1977) Rapid chromosome preparations from solid tissues of fishes.Journal of the Fisheries Research Board of Canada 34(2): 266–269. 10.1139/f 77-039 · doi ↗