Description of two new species of Chiloglanis (Teleostei, Mochokidae) from the Eastern Zimbabwe Highlands freshwater ecoregion: an overlooked hotspot of rheophilic fishes

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
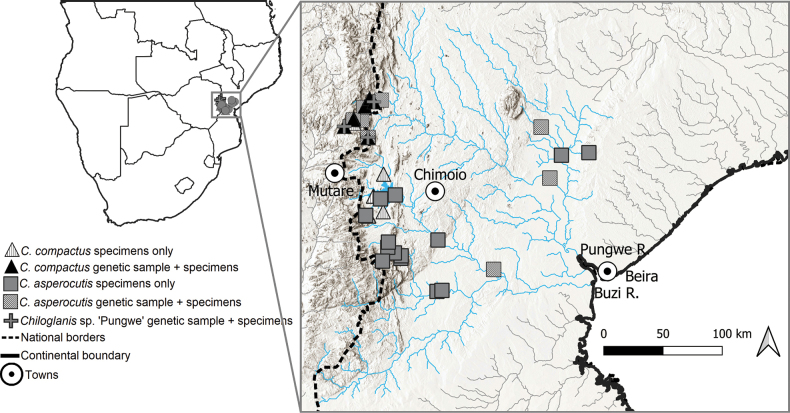 Figure 1
Figure 1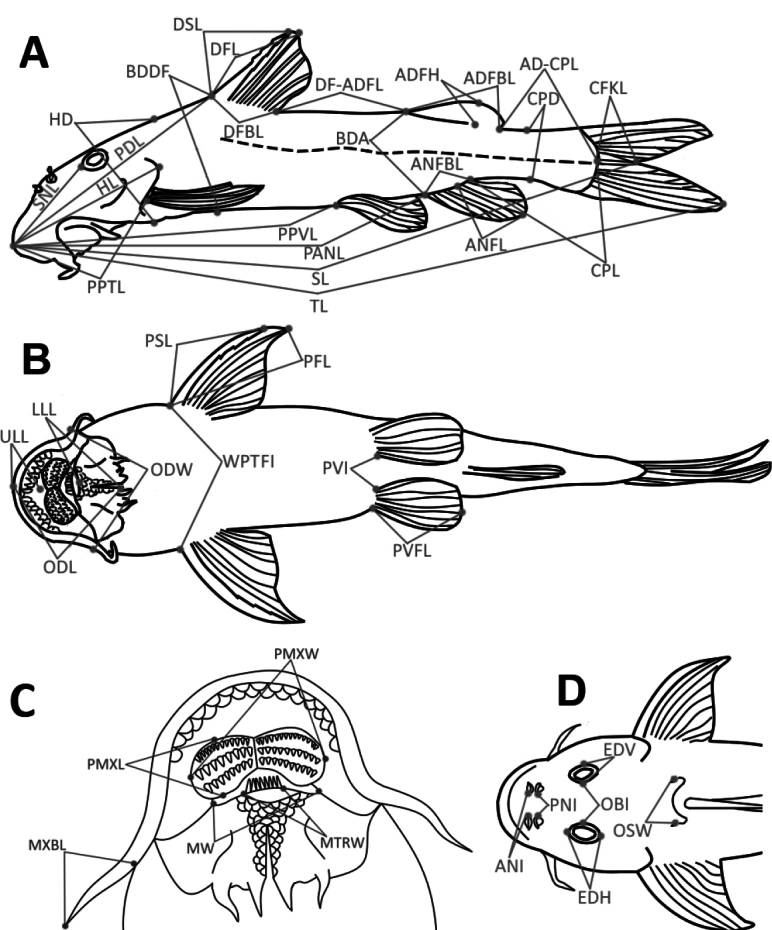 Figure 2
Figure 2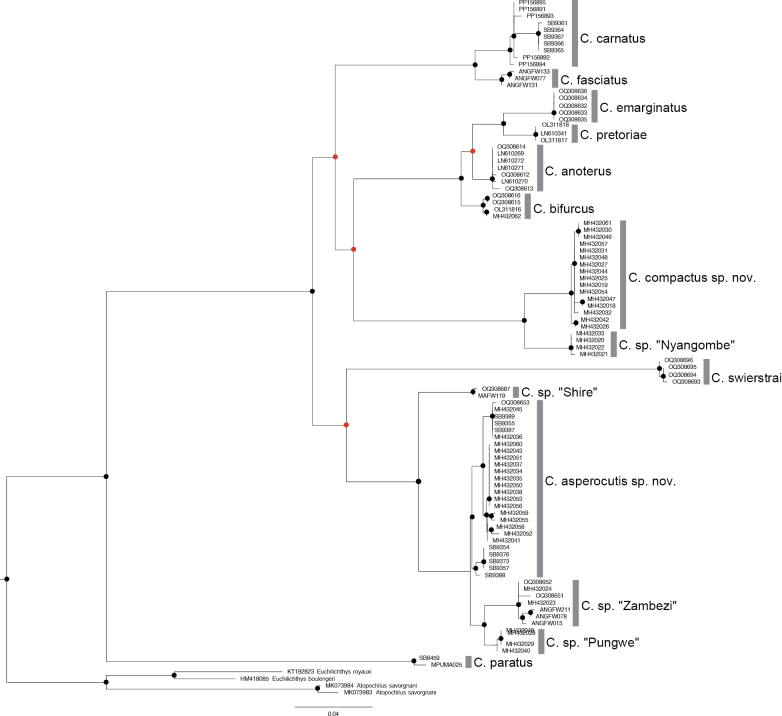 Figure 3
Figure 3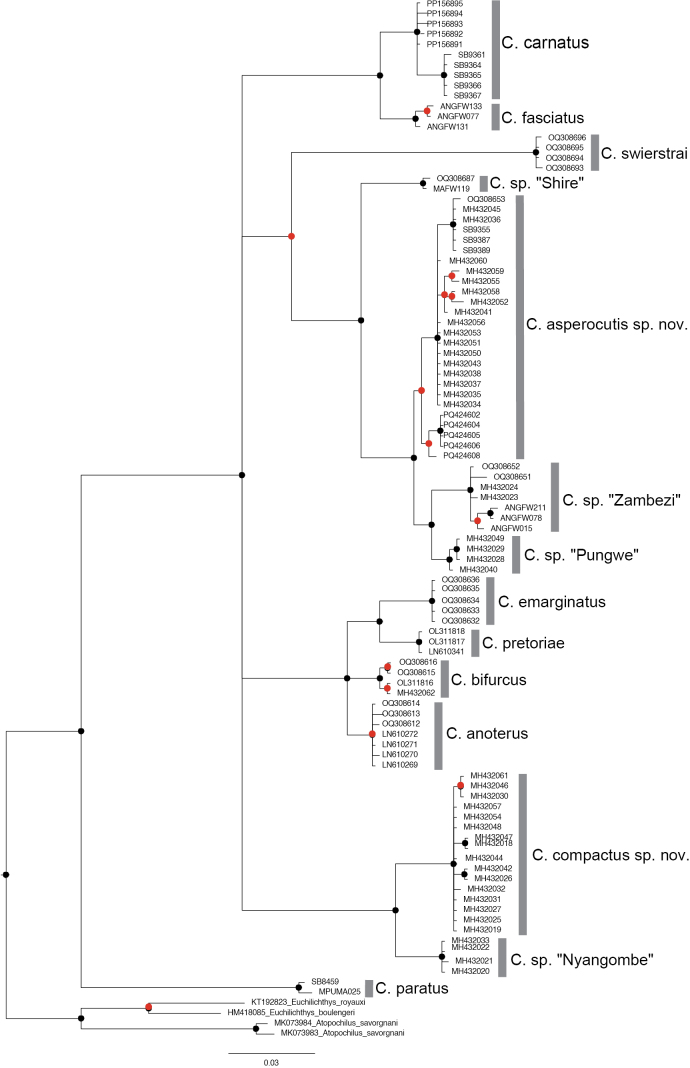 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
| 0–1.7 | ||||||||||||||||
|
|
| 2.9–3.7 | 0.2–0.6 | |||||||||||||||
|
|
| 14.6–15.5 | 14.6–15.4 | 0.0–0.4 | ||||||||||||||
|
|
| 11.7–12.2 | 10.3–10.5 | 14.9–15.4 | 0 | |||||||||||||
|
|
| 10.8–11.7 | 9.9–10.6 | 14.5–15.5 | 5 | 0.0–0.4 | ||||||||||||
|
|
| 10.8–11.7 | 9.9–10.6 | 14.0–14.7 | 4.4–5.0 | 2.5–2.9 | 0.0–0.6 | |||||||||||
|
|
| 11.7–12.4 | 10.5–10.8 | 14.7–15.2 | 3.7 | 4.3 | 3.5–3.9 | 0 | ||||||||||
|
|
| 16.0–16.7 | 15.4–16.4 | 18.1–18.9 | 17.3–17.8 | 14.6–15.6 | 14.9–15.8 | 15.3–15.6 | 0.0–0.6 | |||||||||
|
|
| 13.2–14.5 | 12.3–13.2 | 14.3–15.3 | 11.5–11.7 | 11.3–12.0 | 12.5–13.2 | 11.2–11.5 | 17.2–17.9 | 0–0.8 | ||||||||
|
|
| 10.6–12.5 | 11.5–12.5 | 12.6–13.8 | 10.1–10.6 | 11.5–12.0 | 10.6–11.3 | 12.4–12.9 | 16.2–18.5 | 11.8–13.5 | 0–1.7 | |||||||
|
| 11.6–13.3 | 12.8–13.3 | 13.3–13.8 | 11.2–11.5 | 11.7–12.0 | 10.6–11.3 | 12.0–12.4 | 16.9–17.4 | 12.5–13.5 | 1.7–3.3 | 0–0.2 | |||||||
|
| 11.1–13.3 | 11.8–13.0 | 13.1–13.8 | 11.0–11.5 | 11.1–11.5 | 10.6–11.5 | 11.9–12.4 | 16.2–16.9 | 12.0–13.0 | 2.7–4.1 | 2.1–3.1 | 0–1.3 | ||||||
|
| 11.6–12.3 | 11.6–11.8 | 11.9–12.3 | 10.3–10.5 | 10.4–10.6 | 10.8–11.3 | 11.2–11.4 | 15.7–16.2 | 12.5–13.2 | 4.7–5.8 | 5.4–5.8 | 5.4–6.2 | 0–0.2 | |||||
|
| 14.5–15.8 | 13.7–14.2 | 15.9–16.6 | 12.5–12.7 | 11.8–12.5 | 12.5–13.2 | 11.3–11.5 | 16.4–16.9 | 4.1–4.8 | 10.0–11.4 | 11.1–11.6 | 10.0–10.9 | 11.3–11.8 | 0–0.2 | ||||
|
|
| 18.2–19.0 | 17.5–18.3 | 19.2–19.8 | 17.3–17.8 | 17.8–18.1 | 17.5–18.1 | 17.2–17.5 | 17.1–17.8 | 18.5–19.0 | 17.5–19.2 | 18.3–19.2 | 17.6–18.9 | 16.5–17.0 | 18.3–19.4 | 0–1.2 | ||
|
|
| 17.0–18.0 | 17.5–18.0 | 16.7–16.9 | 17.2–17.2 | 17.0–17.6 | 17.1–17.6 | 17.3–17.3 | 14.9–15.2 | 17.8–18.0 | 16.5–18.1 | 16.8 | 17.3–18.1 | 16.5–16.7 | 17.6–17.8 | 12.8–13.0 | _ | |
|
|
| 18.5–18.8 | 17.8–18.3 | 17.9–18.1 | 19 | 19.3–19.8 | 19.3–20.1 | 19.5 | 15.1–15.6 | 19.5–20.3 | 18.0–20.2 | 18.5 | 19.1–20.2 | 17.2–17.4 | 19.3–19.6 | 12 | 7.1 | _ |
| Species |
|
|
|
|
|
|
|
| ||
|---|---|---|---|---|---|---|---|---|---|---|
| Number of specimens | 55 | 110 | 19 | 16 | 1.0 | 10 | 10 | 10 | 2 | 7 |
| Total length | 28.9–57.2 | 32.3–87.6 | 45.3–62.2 | 31.7–67.1 | 80.1 | 68.7–84.9 | 50.2–66.6 | 37.7–53.3 | 44–51.9 | 45.2–65.7 |
| Standard length | 26.0–45.9 | 33.9–66.6 | 35.5–48.9 | 26.5–54.6 | 61.7 | 51.4–63.9 | 40.3–55.6 | 30.3–41.7 | 35.8–42.4 | 34.9–51.9 |
| Head length | 7.1–14.4 | 11.2–22.8 | 12.1–15.6 | 8.8–19.4 | 20.3 | 15.7–19.5 | 12.3–15.7 | 10.0–13.7 | 11.1–13.8 | 9.2–13.6 |
| % Standard length | ||||||||||
| Caudal peduncle depth | 10.0–12.4 | 7.5–10.0 | 11.3–13.2 | 11.0–13.8 | 12.2 | 11.1–14.1 | 10.2–11.9 | 7.5–8.8 | 9.6–9.9 | 7.2–8.7 |
| Dorsal-fin base length | 9.9–14.2 | 7.9–13.4 | 10.7–14.1 | 12.8–18.0 | 8.6 | 9.5–13.2 | 8.8–12.9 | 10.4–13.7 | 12.8–14.6 | 7.5–9.6 |
| % Head length | ||||||||||
| Lower lip length | 17.5–22.6 | 18.4–27.4 | 18.3–26.6 | 22.4–27.7 | 25.1 | 17.7–25.6 | 23.5–28.8 | 18.8–24.9 | 22.8–27.6 | 19.2–26.0 |
| Mandibular tooth row width | 5.6–10.4 | 3.6–7.2 | 4.6–8.1 | 16.0–25.6 | 10.5 | 10.4–17.3 | 9.6–13.5 | 4.8–6.6 | 7.2–7.7 | 10.0–16.6 |
| Upper lip length | 6.9–17.1 | 11.4–22.2 | 11.1–16.2 | 11.7–18.8 | 16.7 | 8.4–12.3 | 6.6–10.6 | 8.8–15.5 | 12.3–12.4 | 7.0–10.5 |
| Meristics | ||||||||||
| Pectoral-fin rays | 7 | 8 | 8 (6–8) | 8 | 8.0 | 8 (7–8) | 7 | 8 | 8 | 8 |
| Dorsal-fin rays | 5 | 6 (6–7) | 6 (5–7) | 6 | 5.0 | 5 (5–6) | 6 (5–6) | 6 (5–6) | 5 | 5 (5–6) |
| Mandibular teeth | 8 (6–8) | 10 (8–10) | 10 | 12 | 12.0 | 8 | 8 (6–8) | 8 | 12 | 11 (11–14) |
| Primary premaxillary teeth | 38 (31–53) | 80 (68–128) | 60 (43–69) | 51–59 | 86.0 | 54 (50–64) | 36–54 | 64 (51–65) | 39–51 | 50 (34–59) |
| SAIAB catalogue number | 201026 | 201026 | _ |
| Number of specimens | 1 | 42 | 67 |
| Total length | 81.6 | 32.3–77.5 | 43.5–87.6 (65.1) |
| Head length | 21.5 | 17.8–21.5 | 11.2–22.8 (16.8) |
| Standard length | 66 | 61.4–66 | 33.9–66.6 (51.7) |
| % | |||
| Adipose fin to caudal peduncle length | 17.6 | 16–17.6 | 13.5–21.6 (16.4) |
| Adipose-fin base length | 11.7 | 11.7–14.5 | 10.4–17.9 (13.7) |
| Adipose-fin height | 4.7 | 4.7–4.9 | 3.9–7 (5.4) |
| Anal-fin base length | 9.7 | 9.7 | 8.2–13.2 (10.9) |
| Anal-fin length along longest ray | 12.1 | 12.1–16.1 | 10.9–18.9 (14) |
| Body depth at anus | 12.8 | 12.8–15.4 | 12.5–16.9 (14.8) |
| Body depth at dorsal-fin insertion | 19.6 | 18.1–19.6 | 17.2–22.8 (19.4) |
| Caudal peduncle depth | 8.1 | 8.1–8.4 | 7.5–10 (8.9) |
| Caudal peduncle length | 19.1 | 18.8–19.1 | 13.9–21.3 (17.4) |
| Dorsal fin to adipose fin length | 26.4 | 24.9–26.4 | 18.7–29.2 (24.6) |
| Dorsal-fin base length | 12.4 | 11.6–12.4 | 7.9–13.4 (10.2) |
| Dorsal-spine length | 19.4 | 16.7–19.4 | 13.8–25.4 (17.5) |
| Pre-anal length | 73.6 | 70.4–73.6 | 68.4–77.6 (72.8) |
| Pre-dorsal length | 42.5 | 39.2–42.5 | 37.6–46.3 (40.9) |
| Pre-pectoral length | 29 | 26.5–29 | 25.7–33 (29.7) |
| Pre-pelvic length | 57.8 | 57.1–57.8 | 53.6–61.8 (58.8) |
| Pectoral-spine length | 20.8 | 20.8–21.3 | 13.4–26.6 (20.2) |
| Pectoral-fin length | 21.8 | 21.8–25.5 | 19.4–27.4 (22.7) |
| Pelvic-fin length | 14.3 | 13.2–14.3 | 11.7–17.5 (14.6) |
| Width at pectoral-fin insertion | 23.9 | 22.7–23.9 | 21–27.1 (23.9) |
| Pelvic-fin interspace | 4.8 | 4.8–5.4 | 2.8–9.2 (5.1) |
| Head length | 32.6 | 29–32.6 | 28.6–36.3 (32.6) |
| Dorsal-fin length along longest ray | 20.5 | 20.5–20.6 | 12.3–22.8 (17.8) |
| Caudal fork length | 10.3 | 8.5–10.3 | 9.4–15.9 (12.5) |
| % | |||
| Anterior nares interspace | 13.9 | 13.4–13.9 | 10.3–19.5 (14.4) |
| Eye diameter (vertical axis) | 12.5 | 12.5–13.4 | 7.9–15.9 (12.7) |
| Lower lip length | 21.3 | 18.7–21.3 | 18.4–27.4 (22.4) |
| Mandibular tooth row width | 6.4 | 6.4–7.2 | 3.6–6.9 (5.5) |
| Maxillary barbel length | 27.8 | 23.9–27.8 | 20.2–38.9 (28.3) |
| Mouth width | 25.9 | 25.9–31 | 26.6–34.1 (31) |
| Orbital interspace | 20.8 | 20.4–20.8 | 16.5–22.6 (19.9) |
| Oral disc length | 57.2 | 53.7–57.2 | 47.1–63.3 (52.6) |
| Oral disc width | 51.7 | 51.7–59 | 49.6–69.2 (57.9) |
| Premaxillary tooth-patch length | 10.5 | 10.1–10.5 | 7.6–14 (10.6) |
| Premaxillary tooth-patch width | 36.4 | 36.4–46.8 | 29.7–51.2 (43.5) |
| Posterior nares interspace | 17.3 | 10.1–17.3 | 7.5–14.9 (11.3) |
| Snout length | 65 | 60.1–65 | 54.8–68 (63.9) |
| Upper lip length | 13 | 13–16.8 | 11.4–22.2 (15.6) |
| Eye diameter (horizontal axis) | 15.7 | 15.7–17 | 22.3–51.4 (34.2) |
| Occipital shield width | 36.1 | 36.1–40.1 | 48.4–60 (54.2) |
| Head depth | 52 | 52–55.8 | 22.5–89.5 (61.6) |
|
| |||
| Pelvic fin count | 7 | 7 | 7 |
| Pectoral fin count | 8 | 8 | 8 |
| Primary premaxillary tooth row count | 4 | 3–5 | 4 (3–5) |
| Primary premaxillary tooth count total | 102 | 74–102 | 80 (68–128) |
| Mandibular tooth count | 10 | 10 | 10 (8–10) |
| Dorsal-fin rays | 6 | 6 | 6 (6–7) |
| Anal-fin rays | 10 | 10–11 | 9 (9–11) |
| Total vertebrate | 29 | 28–29 | 27 (27–29) |
| Abdominal vertebrae | 13 | 11–13 | 12 (11–13) |
| Caudal vertebrae | 16 | 15–17 | 15–17 |
| SAIAB catalogue number | 210377 | 210377 | _ |
| Number of specimens | 1 | 12 | 42 |
| Total length | 42.7 | 29.8–37.5 | 31.4–57.2 (44.1) |
| Head length | 11.5 | 9.1–11.7 | 7.1–14.4 (11.3) |
| Standard length | 34 | 28.5–41.2 | 26–45.9 (35.2) |
| % | |||
| Adipose fin to caudal peduncle length | 16.2 | 16.2–20 | 11.3–19.8 (16) |
| Adipose-fin base length | 17.3 | 16.2–18.4 | 13.2–19.6 (15.7) |
| Adipose-fin height | 4.6 | 4.6–5.2 | 2.8–7.1 (5.2) |
| Anal-fin base length | 14.8 | 10.6–17 | 10.6–16.4 (13.5) |
| Anal-fin length along longest ray | 18.1 | 14.9–18.1 | 11.8–20.5 (16) |
| Body depth at anus | 16.9 | 16.1–17.2 | 14.3–20.8 (17.9) |
| Body depth at dorsal-fin insertion | 20.9 | 20–21.3 | 16.8–25.3 (21) |
| Caudal peduncle depth | 11.1 | 10.7–11.1 | 10–12.4 (11.4) |
| Caudal peduncle length | 18.7 | 18.7–26.2 | 15.6–24.5 (19.9) |
| Dorsal fin to adipose fin length | 26 | 24.6–31.2 | 19.5–30.2 (24.2) |
| Dorsal-fin base length | 13.5 | 11.1–14.2 | 9.9–11.8 (11.1) |
| Dorsal-spine length | 16.3 | 14.7–19.3 | 11.6–21.6 (15.7) |
| Pre-anal length | 64.4 | 64.1–66.5 | 60.9–71 (67.1) |
| Pre-dorsal length | 39.9 | 39.8–40.7 | 35.4–46.2 (39.7) |
| Pre-pectoral length | 29.1 | 28.8–29.1 | 24.3–33.6 (29.3) |
| Pre-pelvic length | 55.5 | 53.8–57.1 | 51.5–59.3 (55.8) |
| Pectoral-spine length | 18 | 14.9–18 | 13.7–21 (17.1) |
| Pectoral-fin length | 23.8 | 21.7–23.8 | 15.4–25.6 (21.2) |
| Pelvic-fin length | 13.2 | 12.7–13.6 | 10.2–18 (13.7) |
| Width at pectoral-fin insertion | 23.6 | 23–24.8 | 21.6–27.6 (24.2) |
| Pelvic-fin interspace | 4.5 | 4.3–4.5 | 1.8–7.7 (4) |
| Head length | 33.8 | 28.3–33.8 | 25.9–34.9 (31.9) |
| Dorsal-fin length along longest ray | 23.7 | 17.9–23.7 | 7.6–22.4 (15.6) |
| Caudal fork length | 15.9 | 15.4–16.8 | 12.4–24.5 (17.5) |
| % | |||
| Anterior nares interspace | 12.5 | 12.5–17.3 | 10.9–23 (16.4) |
| Eye diameter (vertical axis) | 10.9 | 10.9–13.7 | 11.7–18 (15) |
| Lower lip length | 18.5 | 18.5–22.6 | 17.5–21.3 (19.1) |
| Mandibular tooth row width | 5.6 | 5.6–6.9 | 5.9–10.4 (8.6) |
| Maxillary barbel length | 22.2 | 22.2–37.6 | 17.4–45.7 (30.6) |
| Mouth width | 25.8 | 25.8–32.7 | 28.2–35.1 (31) |
| Orbital interspace | 20.7 | 20.7–23.6 | 24.7–27.5 (25.7) |
| Oral disc length | 46.4 | 46.4–56.5 | 38.6–54.7 (47.6) |
| Oral disc width | 47.3 | 47.3–57.4 | 43.8–69.7 (51.7) |
| Premaxillary tooth-patch length | 6.9 | 6.9–11 | 5.4–14.3 (8.6) |
| Premaxillary tooth-patch width | 39.9 | 39.9–47.9 | 29.8–46.8 (38.9) |
| Posterior nares interspace | 17.8 | 17–17.8 | 12.8–20 (15.6) |
| Snout length | 51.9 | 51.9–65.3 | 51.1–69.7 (57.4) |
| Upper lip length | 9.6 | 9.6–12.1 | 6.9–17.1 (12.7) |
| Eye diameter (horizontal axis) | 13.5 | 13.5–16.5 | 22.9–46 (35.5) |
| Occipital shield width | 37.2 | 37.2–53.1 | 68.7–83.6 (74.3) |
| Head depth | 55.6 | 55.6–60.2 | 59.2–82.3 (69.7) |
|
| |||
| Pelvic fin count | 7 | 7 | 7 |
| Pectoral fin count | 7 | 7 | 7 |
| Primary premaxillary tooth row count | 3 | 2–3 | 2 (2–3) |
| Primary premaxillary tooth count total | 44 | 42–47 | 38 (31–53) |
| Mandibular tooth count | 6 | 6 | 8 (6–8) |
| Dorsal-fin rays | 5 | 5 | 5 |
| Anal-fin rays | 10 | 9–10 | 10 (9–12) |
| Total vertebrate | 28 | 27–29 | 29 (27–29) |
| Abdominal vertebrae | 11 | 11–12 | 11 (11–12) |
| Caudal vertebrae | 17 | 17–18 | 17 (17–18) |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFish Biology and Ecology Studies · Fish biology, ecology, and behavior · Ichthyology and Marine Biology
Introduction
Previous and ongoing studies indicate that many of the currently recognised freshwater fish species in southern African harbour hidden diversity (e.g., Swartz et al. 2009; Chakona and Swartz 2013; Chakona et al. 2015, 2018; Kambikambi et al. 2021; Mazungula and Chakona 2021; Sithole et al. 2023; Mutizwa et al. 2024; Scheepers et al. 2024). Recently, Chakona et al. (2018) provided the first evidence of undocumented diversity and taxonomic conflicts in fishes from high attitude streams in the Eastern Zimbabwe Highlands freshwater ecoregion (EZH), one of the poorly explored regions in southern Africa. Similar findings were reported from a number of studies on species from other high altitude streams in east and west Africa (e.g., Friel and Vigliotta 2011; Schmidt et al. 2015, 2016, 2017, 2023; Morris et al. 2016; Schmidt and Barrientos 2019; Schedel et al. 2024). This indicates that, despite being historically assumed to have a depauperate fish fauna, these high altitude freshwater ecosystems represent previously overlooked hotspots of diversity and endemism (e.g., Schmidt et al. 2017). Growing evidence shows that many groups of fishes inhabiting high altitude streams in Africa are in need of major taxonomic reassessments (e.g., Friel and Vigliotta 2011; Schmidt et al. 2015, 2016, 2017, 2023; Morris et al. 2016; Schmidt and Barrientos 2019).
The aim of the present study was to investigate the taxonomic status of suckermouth catfishes of the genus Chiloglanis Peters, 1868 from the EZH. The first published detailed checklist of freshwater fishes of Zimbabwe by Jubb (1961) contained one species of suckermouth catfish, Chiloglanisneumanni Boulenger, 1911, from the EZH. For reasons that remain unclear in the literature, subsequent publications such as Bell-Cross (1976) listed two species, Chiloglanisemarginatus Jubb and Le Roux, 1969 and C.neumanni, whereas Skelton (2001) recognised three species, C.emarginatus, C.neumanni and Chiloglanispretoriae Van der Horst, 1931 from the same region. Marshall (2011), who summarised the existing knowledge of the fishes of Zimbabwe, concluded that the Limpopo River system represented the northern-most distribution limit for C.pretoriae, and questioned the taxonomic assignment of the EZH suckermouth catfishes to C.neumanni, a species that was described from the Bubu River, a tributary of the Great Ruaha River in Tanzania. Marshall (2011) further commented that C.neumanni and C.emarginatus are unlikely to occur in Zimbabwe and suggested that specimens that were previously assigned to these two species likely represented several undescribed species whose taxonomic identity required further investigation.
Marshall’s (2011) tentative proposition was supported by subsequent results from DNA barcoding studies that showed considerable genetic divergence of suckermouth catfishes of the EZH from currently described species of Chiloglanis from southern Africa (Chakona Et Al. 2018; Mutizwa et al. 2024) and specimens from the Great Ruaha River system in Tanzania (Day Et Al. 2023). Chakona et al. (2018) identified at least three candidate species of Chiloglanis from the Pungwe and Buzi River systems in the EZH. Here, we apply an integrative taxonomic approach combining genetic and morphological analyses to provide evidence supporting the recognition of two of these lineages tentatively named Chiloglanis sp. “rough skin” and Chiloglanis sp. “dwarf” in Chakona et al. (2018), as distinct species.
Materials and methods
Sample collection
Voucher specimens and tissue samples were collected during surveys of the EZH conducted in 2013, 2014 and 2022 in the Buzi and Pungwe river systems (Fig. 1). Sampling was done using a Samus-725M electrofisher with a seine net placed downstream to capture immobilised fish in fast-flowing current. Species were identified using regional identification guides by Skelton (2001) and Marshall (2011). Captured fishes were euthanized with clove oil, some specimens were digitally photographed, and a small piece of muscle tissue was dissected from the right side of each specimen and preserved in 95% ethanol. Tissue samples were stored at -80 °C at the National Research Foundation-South African Institute for Aquatic Biodiversity (NRF-SAIAB), Makhanda. Voucher specimens were fixed in 10% formalin in the field. They were then transferred through 10% and 50% to 70% ethanol for long-term storage. All voucher specimens were deposited into the fish collection facility at the NRF-SAIAB as reference material. In addition, the study included specimens collected from the Pungwe and Buzi Rivers prior to the 2013 that were lodged at the National Fish Collection at NRF-SAIAB (see Material examined).
Collection sites of Chiloglanis specimens from the Pungwe and Buzi river systems.
DNA extraction, amplification, and sequencing
This study used sequences generated by Chakona et al. (2018) and Mutizwa et al. (2024) deposited in GenBank (accession numbers: MH432018–MH432062, PP156890–PP156895). An additional eight COI sequences were generated for this study using specimens collected from tributaries of the Pungwe River in Gorongosa National Park, Mozambique in surveys conducted between July and August 2022 (GenBank accession numbers: PQ424602–PQ424609). Preparation and sequencing of genetic material was done in the Aquatic Genomics Research Platform at the NRF-SAIAB. DNA extraction, amplification, and sequencing followed Mutizwa et al. (2024). Genomic DNA was extracted from preserved tissues using the salting-out method (Sunnucks and Hales 1996). The mitochondrial DNA cytochrome c oxidase subunit I (COI) gene was amplified by polymerase chain reaction (PCR) using the universal fish DNA barcoding primer set FishF1 and FishR1 (Ward et al. 2005). PCRs were performed with a Veriti 96 well thermal cycler (Applied Biosystems, USA) and each reaction mixture (25 µL) contained 50–100 ng) of template DNA, 6.5 µL of water, 0.5 µL of each primer (10 µM), and 12.5 µL Taq DNA polymerase 2 × master mix red (Amplicon PCR enzymes and reagents, Denmark). The PCR amplification profile had an initial denaturation step of 3 min at 94 °C followed by 38 cycles of 30 sec at 94 °C, annealing at 55 °C for 30 sec, and extension at 72 °C for 50 sec, and final extension at 72 °C for 7 min. The amplicons were purified using an Exonuclease I-Shrimp Alkaline Phosphate (Exo/SAP, Thermo Fisher Scientific, USA) protocol (Werle et al. 1994), sequenced using standard fluorescent BigDye v. 3.1 (Applied Biosystems, USA) terminator chemistry in the forward direction, and analysed on a 3500 Genetic Analyser (Applied Biosystems, USA) at the NRF-SAIAB. Locality details for Chiloglanis sp. “dwarf”, Chiloglanis sp. “rough skin” and comparative sequences from southern African Chiloglanis species and outgroup species are given in the Suppl. material 1: table S1).
Phylogenetic analyses
Phylogenetic relationships among Chiloglanis species from southern Africa were inferred using Bayesian and maximum likelihood approaches. Mitochondrial DNA sequences were edited, aligned, and trimmed in MEGA-X (Kumar et al. 2018). The sequences were translated into amino acid sequences in MEGA-X to check for stop codons and gaps to ensure that they were copies of functional mitochondrial protein coding sequences. Haplotype groups were identified using DNASP 6 (Rozas et al. 2017). Bayesian analysis was performed using MrBayes 3.1.2 (Ronquist et al. 2012). The dataset was partitioned by codon position and sampled using a reversible-jump Markov chain Monte Carlo (RJ-MCMC), with time-reversible substitution models and gamma distributed rate heterogeneity (Huelsenbeck et al. 2004). Two parallel analyses of four Markov chains were ran for 10 million generations, sampling of trees was done every 1000 generations discarding the first 25% of trees as burn-in. An average standard deviation of split frequencies of < 0.05 indicated the convergence of the two runs in MrBayes. Effective Sample Size (ESS) values and the potential-scale reduction factor for all parameters examined were > 100 and 1.0, respectively. Resulting trees were visualized in FigTree, v. 1.4.3 (Rambaut 2016). Maximum likelihood (ML) analysis of the same dataset was performed in IQ-TREE (Nguyen et al. 2015) through the webserver at http://iqtree.cibiv.univie.ac.at/ (Trifinopoulos et al. 2016). A total of 1000 Ultrafast bootstraps were performed (Minh et al. 2013). Bootstrap values equal to or higher than 70% (Hillis and Bull 1993), and posterior probability values at 0.95 or higher (Alfaro and Holder 2006) were considered to indicate well supported nodes.
Morphological analysis
The study examined 110 specimens of Chiloglanis sp. “rough skin”, 55 specimens of Chiloglanis sp. “dwarf”, and five specimens of Chiloglanis sp. “Pungwe” from the EZH. These specimens were compared to their southern African congeners using raw data from Mutizwa et al. (2024). A total of ten counts and 43 measurements (Fig. 2) were considered for this study following Friel and Vigliotta (2008) and Skelton and White (1990). The morphometric characters are defined in Suppl. material 1: table S2). Measurements were taken to the nearest 0.1 mm using digital callipers. Postcranial meristics (i.e., total vertebrae, abdominal vertebrae, and caudal vertebrae) were taken from radiographs following criteria described by Skelton and White (1990). Vertebrae counts excluded the Weberian vertebrae. The ural centrum was counted as a single element. Abdominal vertebrae included vertebrae anterior to the leading anal-fin pterygiophore. The caudal vertebrae included all vertebrae posterior to the leading anal-fin pterygiophore. The specimens were sexed by external examination. Males possess an elongated and pointed genital papilla, while females are distinguished by possession of longitudinal invagination between the anus and the uro-genital papilla (Skelton 2001).
Illustrations of linear measurements recorded from Chiloglanis specimens A lateral view B ventral view of the body C ventral view of the oral disc D dorsal view of the head. Character abbreviations are explained in the morphological analysis section.
Results
Molecular analysis
The Maximum Likelihood analysis based on 97 sequences (525 base pairs) recovered 14 distinct clades (Fig. 3). These clades corresponded to the eight species from the genus Chiloglanis that are currently recognised in southern Africa. The remaining clades correspond to the six candidate species that were identified in previous studies and include the two taxa, Chiloglanis sp. ‘rough skin’ and Chiloglanis sp. ‘dwarf’ whose taxonomic integrity is the subject of investigation for the present study (Fig. 3). A similar number of clades was recovered from the Bayesian Inference analysis (Fig. 4). The two lineages, Chiloglanis sp ‘rough skin’ and Chiloglanis sp. ‘dwarf’ were recovered as well-supported monophyletic clades (Figs 3, 4) that are deeply divergent (10.1–18.5%) from all the formally described congeneric species in southern Africa (Table 1). Chiloglanis sp. “dwarf” was deeply divergent from Chiloglanis sp. “rough skin” (11.8–13.5%, Table 1). Intraspecific genetic divergence was low in Chiloglanis sp. “dwarf” (0–0.8%) and Chiloglanis sp. “rough skin” (0–1.7%). Chiloglanis sp. “dwarf” was recovered as sister to an undescribed lineage from the Zambezi River, Chiloglanis sp. “Nyangombe” with a genetic divergence of 4.1–4.8%. Chiloglanis sp. “rough skin” sp. nov. was recovered as sister to a clade with undescribed lineages from the Pungwe River, Chiloglanis sp. “Pungwe”, and the neighbouring Zambezi River, Chiloglanis sp. “Zambezi”. There was relatively low genetic divergence between Chiloglanis sp. “rough skin”, Chiloglanis sp. “Pungwe” (1.7–3.3%) and Chiloglanis sp. “Zambezi” (2.7–4.1%). However, the low genetic divergence between Chiloglanis sp. “rough skin”, Chiloglanis sp. “Pungwe”, and Chiloglanis sp. “Zambezi” was in some cases higher than the interspecific genetic divergence found between some valid southern African Chiloglanis species. For example, C.anoterus Crass, 1960 and C.bifurcus Jubb and Le Roux, 1969 were separated by 2.5–2.9% genetic divergence while C.emarginatus and C.pretoriae were separated by 3.7% (Table 1). The clade formed by Chiloglanis sp. “rough skin”, Chiloglanis sp. “Pungwe” and Chiloglanis sp. “Zambezi” was separated from Chiloglanis sp. “Shire” a lineage from the Shire River by a genetic divergence of 4.7–6.2%. Both Chiloglanis sp. “rough skin” and Chiloglanis sp. “dwarf” occurred in sympatry within the Buzi River system. Within the Pungwe River, Chiloglanis sp. “rough skin” and Chiloglanis sp. “dwarf” co-occurred with Chiloglanis sp. “Pungwe” (Fig. 1). These results further corroborate the presence of undescribed Chiloglanis lineages, such as Chiloglanis sp. “Nyangombe” from the Nyangombe River (Zambezi River system), Chiloglanis sp. “Shire” from the Shire River (Zambezi River system), Chiloglanis sp. “Pungwe” from the Pungwe River and Chiloglanis sp. “Zambezi” from the Zambezi River that were identified in previous studies.
Maximum likelihood tree of the species and lineages of the genus Chiloglanis found in southern African based on cytochrome oxidase I (COI). The grey bars represent sequences belonging to the same species. Node colours indicate bootstrap values: black circle ≥ 70; red circle < 70.
Bayesian inference tree of the species and lineages of the genus Chiloglanis found in southern African based on cytochrome oxidase I (COI). The grey bars represent sequences belonging to the same species. Node colours indicate Bayesian posterior probabilities: black circle ≥ 0.95; red circle < 0.95..
Morphological analysis
Detailed morphological examination revealed a number of phenotypic characters that consistently separate Chiloglanis sp ‘rough skin’ and Chiloglanis sp. ‘dwarf’ from all the eight formally described and allopatrically distributed species from southern Africa. These characters include the caudal peduncle depth, dorsal-fin base length, lower lip length, mandibular tooth row width, upper lip length, number of pectoral-fin rays, number of dorsal-fin rays, number of mandibular teeth, number of primary premaxillary teeth, shape and depth of the caudal fin and the shape of the tubercles (Fig. 5, Table 2).
Some of the readily identifiable diagnostic characters separating Chiloglanisasperocutis from C.compactus. Ridge-like tubercles on the dorsum of aC.asperocutis and conical tubercles on bC.compactus. Deeply forked caudal peduncle with pointed lobes in cC.asperocutis and shallow fork with rounded lobes in dC.compactus.
Taxonomic decisions
Based on the genetic and morphological divergence of Chiloglanis sp. ‘rough skin’ and Chiloglanis sp. ‘dwarf’ from all the formally described congeners from southern Africa and the other candidate species identified from the Eastern Zimbabwe Highlands freshwater ecoregion, below we provide formal descriptions of these two species as Chiloglanisasperocutis sp. nov. and Chiloglaniscompactus sp. nov., respectively. The other lineages identified from the previous and present study will be considered for description in a future study once we have gathered additional material for detailed morphological examination.
Taxonomic accounts
Chiloglanis
asperocutis
Taxon classificationAnimaliaSiluriformesMochokidae
Mutizwa, Bragança & Chakona sp. nov.
11ECD2A0-E470-5006-952C-460FD7B53CF1
https://zoobank.org/0DDBD844-CD0F-432C-9AD0-E11FE7C7095A
Chiloglanis sp. “rough skin”: Chakona et al. 2018: 76–79.
Type material.
Holotype. • SAIAB 246255, female specimen 66.0 mm SL, Honde River, Bridge on road to Honde Mission, Pungwe River system, Manicaland Province, Zimbabwe, 18.5438°S, 32.8044°E, 13 December 2014, Albert Chakona, Wilbert Kadye and Taurai Bere, genseq-1 COIMH432036. Paratypes. • SAIAB 201026, 14 male, 29 female specimens (32.3–77.5 mm SL), Honde River, Pungwe River system, Bridge on road to Honde Mission, Manicaland Province, Zimbabwe, 18.5438°S, 32.8044°E, 13 December 2014, Albert Chakona, Wilbert Kadye and Taurai Bere.
Other material examined.
Specimens detailed in Suppl. material 1.
Diagnosis.
A higher number of primary premaxillary teeth (68–128) readily distinguishes C.asperocutis sp. nov. from all its congeners from southern African that consistently have fewer than 68 primary premaxillary teeth except for C.anoterus and C.carnatusMutizwa et al., 2024. Possession of ten closely packed mandibular teeth further distinguishes C.asperocutis from C.fasciatus Pellegrin, 1936 that has eight closely packed mandibular teeth; C.compactus sp. nov., C.bifurcus and C.emarginatus that have eight widely spaced mandibular teeth; C.anoterus, C.paratus Crass, 1960, and C.pretoriae that have 12 closely packed mandibular teeth; and C.swierstrai Van der Horst, 1931 that has 14 closely packed mandibular teeth. Chiloglanisasperocutis sp. nov. possesses more dorsal fin rays (6–7) compared to C.compactus sp. nov. (5 rays). Possession of eight pectoral fin rays distinguishes C.asperocutis from C.compactus sp. nov. and C.emarginatus that possess seven pectoral fin rays. A deeply forked caudal fin readily separates C.asperocutis sp. nov. from C.compactus sp. nov., C.pretoriae, C.paratus, and C.swierstrai that have shallower fork depths in the caudal fin; C.emarginatus that has an emarginated caudal fin with a very shallow fork; and from C.anoterus that has a caudal fin with no fork due to extended median rays in males and emarginated in females. A caudal fin with an upper lobe shorter than lower lobe distinguish C.asperocutis sp. nov. from C.bifurcus that has a caudal fin with an upper lobe that is longer than the lower lobe. An oral disc with a well-developed mid-ventral cleft distinguishes C.asperocutis sp. nov. from C.swierstrai that lacks a mid-ventral cleft. Chiloglanisasperocutis sp. nov. has a dorsal spine with gently serrated anterior and posterior margins that distinguish it from C.paratus that has a dorsal spine with a serrated posterior margin. Absence of a fleshy skin on the basal portion of the dorsal fin separates C.asperocutis sp. nov. from C.carnatus, a character present in the latter species. Chiloglanisasperocutis sp. nov. is further distinguished from its southern African congeners by long ridge-like tubercles distributed across the dorsal and lateral surfaces of the head and body compared to conical tubercles found in C.compactus sp. nov., C.bifurcus, C.emarginatus, C.fasciatus, C.paratus, C.carnatus, and C.swierstrai; and the lack of conspicuous tubercles in C.anoterus and C.pretoriae. A narrow caudal peduncle depth distinguishes C.asperocutis sp. nov. (7.5–10.0%SL) from C.carnatus (11.3–13.2%SL), C.pretoriae (11.0–13.8%SL), C.anoterus (12.2%SL), C.bifurcus (11.1–14.1%SL), and C.emarginatus (10.2–11.9%SL) that have deeper caudal peduncles. A narrow mandibular tooth row width separates C.asperocutis sp. nov. (3.6–7.2%HL) from C.pretoriae (16.0–25.6%HL), C.anoterus (10.5%HL), C.bifurcus (10.4–17.3%HL), C.emarginatus (9.6–13.5%HL), C.paratus (7.2–7.7%HL), and C.swierstrai (10.0–16.6%HL) that have wider mandibular tooth row widths. A longer upper lip length further separates C.asperocutis sp. nov. (11.4–22.2%HL) from C.emarginatus (6.6–10.6%HL) and C.swierstrai (7.0–10.5%HL) that have shorter upper lips.
Description.
Figs 6, 7 shows general body features of adult male and female C.asperocutis. Morphometric and meristic data for holotype, paratypes, and non-type specimens are presented in Table 3. Where meristics counts have a range, counts from the holotype are given in parentheses.
Chiloglanisasperocutis female holotype SAIAB 246255. Scale bars: 2 cm.
Chiloglanisasperocutis male paratype SAIAB 201026. Scale bars: 2 cm.
Body elongate. Depressed pre-dorsal region, mid-sections more cylindrical, slightly laterally compressed posterior section. Pre-dorsal profile convex, sharply slopping from snout to posterior nostril, gentler slope from poster nostril to dorsal-fin origin. Body greatest depth at dorsal-fin insertion. Post-dorsal profile almost straight to adipose fin insertion, gently concave from adipose-fin origin to caudal fin. Ventral profile gently convex from region just posterior of oral disc to anal-fin origin, gently concave from anal-fin origin to caudal fin.
Skin numerous tubercles spread across body with exception of ventral surface, forming distinct ridge like structures most conspicuous on head dorsum giving skin a distinct “rough” texture. Lateral line complete, originating just above horizontal level of orbit, below anterior dorsal fin origin, gently undulating along midlateral of body to base of caudal fin.
Head relatively big; longer than body depth; ~ 2/3 of pre-dorsal length. Eye large and oval, orbit lacks free margin; located dorsally, closer to margin of operculum than tip of snout. Interorbital space slightly wider than space between nares spaces. Posterior nares slightly closer together than anterior nares. Anterior nares with short skin flaps on their posterior margins. Posterior nares with short skin flaps along anterior margin. Gill opening restricted, located adjacent to pectoral fin origin.
Mouth inferior, projecting below ventral body surface, upper and lower lips combined into a distinct large oral disc. Oral disc width greater than length. Upper and lower lips with pronounced roundish papillae, largest papillae concentrated around mid-ventral cleft of lower lip. Three unbranched pairs of barbels. Maxillary barbel, originating from lateral region of oral disc, extending to posterior region of oral disc. Lateral mandibular barbel longer than medial mandibular barbel, both integrated into lower lip.
Dentation variable number 68–128 (102) of primary pre-maxillary teeth arranged in 3–5 (4) rows on two ovoid tooth patches. Secondary pre-maxillary teeth small and scattered on posterior surface of premaxillae. Tertiary teeth small and needle-like; in a row near midline of dorsal edge of tooth plate. Up to 10 (10) closely packed mandibular teeth; central teeth projecting higher than outer teeth forming a gentle arc; replacement tooth row emerges anteriorly to functional row, number of mandibular teeth may vary due to loss of teeth and their replacements, but the number does not exceed ten.
Vertebral counts Total vertebrae 27–29 (29), abdominal vertebrae 11–13 (13), caudal vertebrae 15–17 (16).
Urogenital papillae distinctly elongate and sharply pointed in males, reduced to a shallow invagination in females.
Pectoral fin ray count 8, origin adjacent to gill opening; pectoral spine slightly curved, anterior margin smooth; gently serrated posterior margin; pectoral spine length ~80% of longest pectoral-fin ray. Dorsal fin originating in anterior 1/3 of body, posterior to pectoral-fin origin, 6–7 (6) branched rays. Dorsal spine, length ~ 80% of longest dorsal-fin ray, gentle serrations on anterior and posterior margins of the terminal 1/4 of dorsal spine. Pelvic fin ray count 7, origin posterior to midpoint between end of dorsal-fin and adipose-fin origin; rounded. Adipose fin relatively small, margin convex, located in posterior 1/3 of body, origin just anterior of anal-fin origin, ridge-like tubercles sometimes present on its surfaces. Anal fin ray count 12–13 (13), origin just posterior to origin of adipose fin; terminating just before posterior end of adipose fin; rounded. Caudal fin deeply forked, lobes unequal, lower lobe generally longer, both lobes with gently pointed tips.
Coloration in ethanol dorsal and lateral skin generally dark brown becoming a lighter shade of brown below mid lateral line. Dark melanophores distributed throughout dorsal and lateral surfaces. Dorsal surface has blotches of lighter shades of brown located between orbits and posterior nostril, at dorsal fin insertion, between dorsal and adipose fin, and posterior to adipose fin, all these blotches extend to lateral surface. Lateral surface with large pale blotches such as those extending from dorsal surface. Lateral line clearly visible as a continuous pale brown stripe with very small pale blotches distributed above and below it. Ventral surface pale cream to yellowish in colour with dark melanophores only present at base of pelvic fins. Dorsal and adipose fins dark brown base and middle with hyaline margins. Pectoral fins have dark brown dorsal surfaces and lighter ventral surface, hyaline margins. Pelvic and anal fins pale brown in colour with dark melanophores and hyaline margins. Dark band stretches across middle of upper caudal fin lobe, lower lobe mostly covered by a dark blotch originating from the fin base. Live colouration displayed similar patterns as described in persevered specimens with a few differences. All pale brown sections in live specimens have a golden colour. Live specimens display a range of pigmentation intensity, some specimens are very dark with patterns described in the Coloration in ethanol section being very clear whilst other specimens are lightly coloured (golden) with little pigmentation visible.
Reproduction: there are no dedicated studies on the breeding biology of C.asperocutis, but spawning is likely to begin in summer (October–November) based on the general pattern of other congeners (Skelton 2001), and other mochokid fishes from this region.
Distribution, habitat, and feeding ecology.
this species is known from multiple localities in the Pungwe and Buzi River systems. It is a rheophilic species that occurs in rocky habitats with fast flowing water. Macroinvertebrates from the families Simuliidae, Chironomidae, Hydropsychidae and Libellulidae were the dominant prey item for this species with algae forming a smaller component of the diet of this species (Matomela et al 2018).
Etymology.
a combination of the Latin words, aspero, meaning rough, and cutis, meaning skin. The name refers to the distinct ridges on the skin (more pronounced on the head dorsum) which is characteristic of this species.
Chiloglanis
compactus
Taxon classificationAnimaliaSiluriformesMochokidae
Mutizwa, Bragança & Chakona sp. nov.
63245AD1-94A7-555D-9175-1FCC90425310
https://zoobank.org/3EEC0C8D-4992-4A8C-9226-1BE59BAAB312
Chiloglanis sp. “dwarf”: Chakona et al. 2018: 76–79.
Type material.
Holotype. • SAIAB 246256, male, right middle cut, 44.4 mm SL, Makanga River Bridge on road to Hauna growth point, Pungwe River system, Manicaland Province, Zimbabwe, 18.5438°S, 32.801°E, 16 December 2014, Albert Chakona, Wilbert Kadye and Taurai Bere, genseq-1 COIMH432044. Paratypes. • SAIAB 210377, 10 males (29.8–37.5 mm SL), 2 females (29.1–40.3 mm SL), Makanga River bridge on road to Hauna growth point, Pungwe River system, Manicaland Province, Zimbabwe, 18.5438°S, 32.801°E, 16 December 2014, Albert Chakona, Wilbert Kadye and Taurai Bere.
Other material examined.
Specimens detailed in Suppl. material 1.
Diagnosis.
Chiloglaniscompactus sp. nov. attains the smallest size (< 46 mm SL) for all congeners currently known from southern Africa. Possession of seven pectoral fin rays in C.compactus sp. nov. distinguishes this species from C.asperocutis, C.carnatus, C.anoterus, C.fasciatus, C.paratus, C.pretoriae, and C.swierstrai that all have eight pectoral fin rays. Possession of eight widely spaced mandibular teeth further distinguishes C.compactus sp. nov. from C.fasciatus (eight closely packed mandibular teeth); C.asperocutis and C.carnatus (ten closely packed mandibular teeth); C.anoterus, C.paratus, and C.pretoriae (12 closely packed mandibular teeth); and C.swierstrai (14 closely packed mandibular teeth). Fewer primary premaxillary teeth in C.compactus sp. nov. (31–53) readily distinguished it from C.asperocutis (68–128) and C.anoterus (86). Fewer dorsal fin rays (5) readily distinguish C.compactus sp. nov. from C.asperocutis (6–7). A relatively shallow forked tail and rounded caudal fin lobes readily separate C.compactus sp. nov. from C.asperocutis, C.carnatus, and C.fasciatus (deeply forked caudal fin with pointed lobes); C.emarginatus (emarginated caudal fin with very shallow fork); and from C.anoterus that has a caudal fin with no fork due to extended median rays in males and emarginated in females. A lower caudal fin lobe longer than the upper lobe separates C.compactus sp. nov. from C.bifurcus that has a longer upper caudal fin lobe. A well-developed mid-ventral cleft in the oral disc of C.compactus sp. nov. distinguishes it from C.swierstrai that lacks a mid-ventral cleft. Gently serrate anterior and posterior margins of the dorsal spine in C.compactus sp. nov. distinguish it from C.paratus that has a dorsal spine serrated only on the posterior margin. Absence of a fleshy skin on the basal portion of the dorsal fin separates C.compactus sp. nov. from C.carnatus, a character present in the latter species. Conical tubercles distributed across dorsal and lateral surfaces of the head and body distinguish C.compactus sp. nov. from C.asperocutis with ridge like tubercles; C.anoterus and C.pretoriae that lack conspicuous tubercles. A deep caudal peduncle distinguishes C.compactus sp. nov. (10.0–12.4%SL) from C.asperocutis (7.5–10.0%SL), C.fasciatus (7.5–8.8%SL), C.paratus (9.6–9.9%SL), and C.swierstrai (7.2–8.7%SL) that have narrower caudal peduncles. A longer dorsal-fin base separates C.compactus sp. nov. (9.9–14.2%SL) from C.swierstrai (7.5–9.6%SL) and C.anoterus (8.6%SL) that possess shorter dorsal-fin bases. Shorter lower lips distinguish C.compactus sp. nov. (17.5–22.6%HL) from C.pretoriae (22.4–27.7%HL), C.anoterus (25.1%HL), C.emarginatus (23.5–28.8%HL), and C.paratus (22.8–27.6%HL) that have longer lower lips. A narrow mandibular tooth row separates C.compactus sp. nov. (5.6–10.4%HL) from C.asperocutis (3.6–7.2%HL), C.pretoriae (16.0–25.6%HL), C.bifurcus (10.4–17.3%HL), and C.swierstrai (10.0–16.6%HL) that possess greater mandibular tooth row widths.
Description.
Figs 8, 9 shows general body features of adult male and female C.compactus. Morphometric and meristic data for holotype, paratypes and non-type specimens are presented in Table 4. Where meristics counts have a range counts from the holotype are given in parentheses.
Chiloglaniscompactus male holotype SAIAB 246256. Scale bars: 2 cm.
Chiloglaniscompactus female paratype SAIAB 210377. Scale bars:2 cm.
Body short and rotund. Depressed pre-dorsal region, mid-sections more cylindrical, slightly laterally compressed posterior section. Pre-dorsal profile convex, almost continuous rounded profile from snout to dorsal-fin origin. Body greatest depth at dorsal-fin insertion. Post-dorsal profile convex to adipose fin insertion, becoming gently concave from adipose-fin origin to caudal fin. Ventral profile convex from region just posterior of oral disc to anal-fin origin, becoming gently concave from anal-fin origin to caudal fin.
Skin numerous conical tubercles spread across body with exception of ventral surface. Lateral line complete, originating above horizontal level of orbit, below anterior dorsal fin origin, almost completely straight along midlateral of body to base of caudal fin.
Head relatively big; longer than body depth; ~ 2/3 of pre-dorsal length. Eye large and oval, orbit lacks free margin; located dorsally, closer to margin of operculum than tip of snout. Interorbital space wider than space between nares spaces. Posterior nares slightly closer together than anterior nares. Anterior nares with short skin flaps on posterior margin. Posterior nares with short skin flaps along anterior margin. Gill opening restricted, located adjacent to pectoral fin origin.
Mouth inferior, projecting below ventral body surface, upper and lower lips combined into a distinct large oral disc. Oral disc width greater than length. Upper and lower lips with pronounced roundish papillae, largest papillae concentrated around mid-ventral cleft of lower lip. Three pairs of barbels. Maxillary barbel unbranched, originating from lateral region of oral disc, extending to posterior region of oral disc. Lateral mandibular barbel longer than medial mandibular barbel, both integrated into lower lip.
Dentation variable number 31–53 (44) of primary pre-maxillary teeth arranged in 2–3 (3) rows on two ovoid tooth patches. Secondary pre-maxillary teeth small and scattered on posterior surface of premaxillae. Tertiary teeth small and needle-like; in a row near midline of dorsal edge of tooth plate. Up to 8 (6) spaced out mandibular teeth; replacement tooth row emerges anteriorly to the functional row, number of mandibular teeth may vary due to loss of teeth and their replacement, but the number hardly ever exceeds 8.
Vertebral counts Total vertebrae 27–29 (28), abdominal vertebrae 11–12 (11), caudal vertebrae 17–18 (17)
Urogenital papillae distinctly elongate and sharply pointed in males, reduced to a shallow invagination in females.
Pectoral fin ray count 7, origin adjacent to gill opening; pectoral spine slightly curved, anterior margin smooth; gently serrated posterior margin; pectoral spine length ~ 80% of longest pectoral-fin ray. Dorsal fin originating in anterior 1/3 of body, posterior to pectoral-fin origin, five branched rays, conical tubercles sometime present on its surfaces. Dorsal spine, length ~ 80% of longest dorsal-fin ray, gentle serrations on anterior and posterior margins of terminal 1/4 of dorsal spine. Pelvic fin ray count 7, origin posterior to midpoint between end of dorsal-fin and adipose-fin origin; rounded. Adipose fin relatively small, margin convex, located in posterior 1/3 of body, origin just anterior of anal-fin origin, conical tubercles sometimes present on its surfaces. Anal fin ray count 12–13 (13), origin just posterior to origin of adipose fin; terminating just before posterior end of adipose fin; rounded. Caudal fin with shallow fork, lobes unequal with lower slightly longer, both lobes rounded, conical tubercles sometime present on their surfaces.
Coloration in ethanol dorsal and lateral skin generally dark brown becoming a lighter shade of brown below mid lateral line. Dark melanophores distributed throughout dorsal and lateral surfaces. Dorsal surface has large blotches of lighter shades of brown located at base of dorsal fin, between dorsal and adipose fin, and posterior end of adipose fin; these blotches extend to lateral surface. Lateral surface with large pale blotches such as those extending from the dorsal surface. Additional large blotches on lateral surface are found below lateral line above pelvic fin, anal fin and caudal peduncle. Approximately seven small circular blotches just above lateral line, two or three similar blotches found above each of them. Approximately four small circular blotches below the lateral line distributed from below dorsal fin to pelvic fin. Lateral line clearly visible as a continuous pale brown stripe. Ventral surface pale cream to yellowish in colour with dark melanophores only present at base of pelvic fins and caudal peduncle. Dorsal and adipose fins brown base and middle with hyaline margins. Pectoral fins have dark brown dorsal surfaces and lighter ventral surface, hyaline margins. Pelvic and anal fins pale brown in colour with hyaline margins. Dark band stretches across middle of upper caudal fin lobe, lower lobe mostly covered by a dark blotch originating from the fin base. Live colouration displayed similar patterns as described in the preserved specimens with a few differences. Live specimens display a range of pigmentation intensity, some specimens are very dark with patterns described in the Coloration in ethanol section being very clear whilst other specimens are lightly coloured with little pigmentation visible.
Reproduction: there have been no dedicated studies on the breeding biology of C.compactus, but spawning is likely to begin in summer (October–November) based on the general pattern of other congeners (Skelton 2001), and other mochokid fishes from this region.
Distribution, habitat, and feeding ecology.
this species occurred at multiple localities in the Pungwe and Buzi river systems with the majority of the collections occurring at high elevation. It is a rheophilic species that occurs in rocky habitats with fast flowing water. Its diet was not examined.
Etymology.
the name is drawn from the word compact which is inspired by the short and rotund body shape of this species as well as it being the smallest of all the currently known congeners in southern Africa.
Discussion
Consistent with Marshall’s (2011) proposition regarding the taxonomic uncertainties within the genus Chiloglanis in central southern Africa, and the subsequent work by Chakona et al. (2018), this study identified two new suckermouth catfishes that were previously lumped together into a single species, C.neumanni. The two newly described species, C.asperocutis and C.compactus, are sympatrically distributed in the Pungwe and Buzi river systems. The description of these new species from the EHZ, together with the recent discovery of C.carnatus in the middle Zambezi River (Mutizwa et al. 2024) and the occurrence of several lineages (e.g., Chiloglanis sp. “Pungwe”, Chiloglanis sp. “Zambezi”, Chiloglanis sp. “Nyangombe” and Chiloglanis sp. “Shire”) highlights this region as a potentially overlooked biodiversity hotspot of rheophilic taxa. This is consistent with the presence of several recently discovered rheophilic species in other genera such as Anoplopterusmarshalli, A.leopardus, Heteromormyrus sp. “Pungwe”, Heteromormyrus sp. “Buzi”, and Zaireichthysmonomotapa that are endemic to the Buzi and Pungwe river systems as well as the tributaries of the Lower Zambezi that drain the EZH ecoregion (Eccles et al. 2011; Mazungula and Chakona 2021; Mutizwa et al. 2021).
Preliminary examination of specimens of the undescribed Chiloglanis lineages (e.g., Chiloglanis sp. “Zambezi”, Chiloglanis sp. “Nyangombe”, and Chiloglanis sp. “Shire”) show some consistent characters that distinguish them suggesting that they are distinct species yet to be described. In the present study, we refrained from making taxonomic decisions for these lineages because we require additional specimens, particularly mature specimens for a thorough morphological examination. Chiloglanisasperocutis morphologically resembles the sympatric Chiloglanis sp. “Pungwe” lineage. The most conservative decision would be to include Chiloglanis sp. “Pungwe” under C.asperocutis, but these two taxa are genetically divergent. Detailed morphological examination of a larger sample size of mature specimens, as well as additional evaluations, including, for example CT scan or cleared and stained samples may identify diagnostic characters that reliably distinguish these two taxa.
The discovery of high levels of previously unrecognised diversity of suckermouth catfishes with largely endemic distribution patterns, for example C.compactus and C.asperocutis endemic to the Pungwe and Buzi rivers, has created renewed interest in the biogeography and evolutionary history of the freshwater fishes of this region. The genus Chiloglanis is inferred to have originated in the Congo Basin in the mid-Eocene around 47.53 Ma (95% HPD: 44.65–50.8 Ma) and its current wide distribution across the African continent indicates a complex biogeographic history (Day et al 2023). Available evidence from ancestral range reconstruction for Chiloglanis suggests that the clade comprising taxa from the Zambezi, East African and Congo Basin ichthyofaunal provinces (Roberts 1975) underwent diversification as a result of multiple dispersal and recolonization events among these regions (Day et al. 2023). This complex biogeographic history is reflected in the close phylogenetic relationships of most southern African species such as C.compactus, C.asperocutis, C.carnatus, C.anoterus, C.pretoriae, C.bifurcus, and C.emarginatus with species and lineages from the Zambezi River, whilst a few species such as C.paratus and C.swierstrai have closer ties to species from the Congo Basin (Day et al. 2023). The polyphyletic relationship between the sympatrically distributed C.compactus and C.asperocutis also suggests complex speciation process within the EZH potentially influenced by dispersal between the Zambezi and the Pungwe and Buzi rivers. These hypotheses need to be tested using comparative biogeographic approaches, particularly integrating phylogenomics to more accurately reconstruct the evolutionary relationships and biogeographic patterns of these species. To achieve this, the resolution of the taxonomic uncertainties and more accurate mapping of species distribution ranges is a fundamental requirement. This study provided evidence of the occurrence of two new species in the EZH, highlighting additional taxonomic inconsistencies and paving the way for future studies.
The new species described in this study are restricted to rivers systems (Pungwe and Buzi rivers) that currently face multiple threats from anthropogenic activities. Rich deposits of precious minerals including diamonds and gold support commercial and artisanal mining activities in the EZH. Alluvial gold extraction, in particular, is carried out mainly along rivers and small streams leading to the destruction of the riparian vegetation, increased sediments loads, altered flow regimes and often mercury contamination (Ndunguru et al. 2006; Clark et al. 2019). Exacerbating the situation is the need for better enforcement of regulations to enable sustainable use of the mineral resources in the EZH (Ndunguru et al. 2006; Spiegel 2015; Clark et al. 2019; Kachena and Spiegel 2019). Additionally, climate change, large scale monoculture plantations and riverbank cultivation often alter physiochemical parameters of river systems negatively impacting aquatic biodiversity in the EZH (Clark et al. 2017, 2019; Mafuwe and Moyo 2020). Furthermore, rainbow trout Oncorhynchusmykiss (Walbaum 1792) was introduced into the rivers of the EZH for angling and this species negatively influences both the distribution and abundance of native fish and macro-invertebrate species in the rivers of this region (Kadye et al. 2013). Many of the threats to the environment in the EZH arise due to socio-economic activities, however, there is need to for balancing them with conservation needs in order to ensure long term sustainability of the unique fauna, flora and landscapes that play an important part in supporting the communities in this region.
The EZH is a recognised area of high plant diversity and endemism (van Wyk and Smith 2001; Clark et al. 2017). The present and previous studies have uncovered previously overlooked diversity of endemic rheophilic fishes at a time when critical instream habitats are being degraded by multiple human impacts. This places responsibility on conservation authorities to implement measures to reduce or halt the ongoing impacts on these fragile ecosystems. In addition to its unique biodiversity, this region provides ecosystems services of national importance to both Mozambique and Zimbabwe that include water production, carbon sequestration, tourism, non-forest timber products, commercial forestry and highland agriculture, historical, cultural and spiritual value (Clark et al. 2019). Although the benefits from this region are well establish, to date there has been few studies that rigorously quantify the status of ecosystems in this region and the benefits they provide. To address this, the description of species unique to this region and mapping of their distribution provides a base line for assessing the health of the ecosystems of the EZH. For example, due to their dependence on perennial rheophilic habitats, species from the genus Chiloglanis along with other habitat specific species (e.g., aquatic invertebrates) have been used as biological indicators to study habitat integrity and instream flow requirements (Kleynhans 1999; Kadye and Moyo 2008; Rashleigh et al. 2009; Soko and Gyedu-Ababio 2015; Achieng et al. 2021). There is need for raising awareness on the uniqueness of the EZH riverine habitats and their endemic biota with emphasis on promoting transdisciplinary approaches involving representatives from local communities, government, industry, academia, NGOs, and conservationists to identify effective strategies to balance socio-economic development with sustainable management and protection of the unique biodiversity of this previously overlooked freshwater ecoregion.
Supplementary Material
XML Treatment for Chiloglanis asperocutis
XML Treatment for Chiloglanis compactus
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Achieng AO Masese FO Coffey TJ Raburu PO Agembe SW Febria CM Kaunda-Arara B (2021) Assessment of the ecological health of Afrotropical rivers using fish assemblages: A case study of selected rivers in the Lake Victoria Basin, Kenya.Frontiers in Water 2(02): 620704. 10.3389/frwa.2020.620704 · doi ↗
- 2Alfaro ME Holder MT (2006) The posterior and the prior in Bayesian phylogenetics.Annual Review of Ecology, Evolution, and Systematics 37(1): 19–42. 10.1146/annurev.ecolsys.37.091305.110021 · doi ↗
- 3Bell-Cross G (1976) The fishes of Rhodesia. National Museums & Monuments of Rhodesia, Salisbury, 182–183 pp.
- 4Chakona A Swartz ER (2013) A new redfin species, Pseudobarbusskeltoni (Cyprinidae, Teleostei), from the Cape Floristic Region, South Africa.Zootaxa 3686(5): 565–577. 10.11646/zootaxa.3686.5.526473242 · doi ↗ · pubmed ↗
- 5Chakona A Malherbe WS Gouws G Swartz ER (2015) Deep genetic divergence between geographically isolated populations of the goldie barb (Barbuspallidus) in South Africa: potential taxonomic and conservation implications.African Zoology 50(1): 5–10. 10.1080/15627020.2015.1021164 · doi ↗
- 6Chakona A Kadye WT Bere T Mazungula DN Vreven EJWMN (2018) Evidence of hidden diversity and taxonomic conflicts in five stream fishes from the Eastern Zimbabwe Highlands freshwater ecoregion.Zoo Keys 768: 69–95. 10.3897/zookeys.768.21944 PMC 601947629955212 · doi ↗ · pubmed ↗
- 7Clark VR Timberlake JR Hyde MA Mapaura A Palgrave MC Wursten BT Ballings P Burrows JE Linder HP Mc Gregor GK Chapano C Plowes DCH Childes SL Dondeyne SMüller T Barker NP (2017) A first comprehensive account of floristic diversity and endemism on the Nyanga Massif, Manica Highlands (Zimbabwe–Mozambique).Kirkia 19(1): 1–53.
- 8Clark V Rde Deus Vidal Jr J Grundy IM Fakarayi T Childes SL Barker NP Linder HP (2019) Bridging the divide between intuitive social-ecological value and sustainability in the Manica Highlands of southern Africa (Zimbabwe-Mozambique). Ecosystem Services 39: 100999. 10.1016/j.ecoser.2019.100999 · doi ↗