Non-radiative origin for alpine endemics of Draba (Brassicaceae) in the central mountains of the Japanese Archipelago
Ryutaro Koda, Yoshinori Murai, Hajime Ikeda

TL;DR
This study shows that alpine Draba species in central Japan evolved independently rather than through a single radiation event.
Contribution
The paper demonstrates a non-radiative origin for alpine Draba endemics in central Japan using phylogenetic and ancestral area analyses.
Findings
Four Draba taxa from central Honshu do not form a monophyletic group.
Divergence occurred from at least two independent evolutionary lineages.
Ancestral species likely originated from different geographical regions.
Abstract
Diversity of endemic species may result from two evolutionary processes: in-situ diversification (radiation) or phylogenetically independent divergences (non-radiation). To explore the evolutionary history of endemic-rich alpine flora in the Japanese Archipelago, we investigated the phylogenetic relationships of four endemic taxa of Draba (Brassicaceae) from the high mountains of central Honshu: D. kitadakensis, D. sachalinensis var. shinanomontana, D. sakuraii var. nipponica, and D. shiroumana. Using molecular phylogenetic analyses on plastid trnL-F and nuclear ITS sequence of 157 taxa, we found that the four taxa endemic to central Honshu did not form a monophyletic group and diverged from at least two evolutionary independent lineages. Moreover, ancestral area reconstruction further revealed that some of their ancestral species may have originated from different geographical regions.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
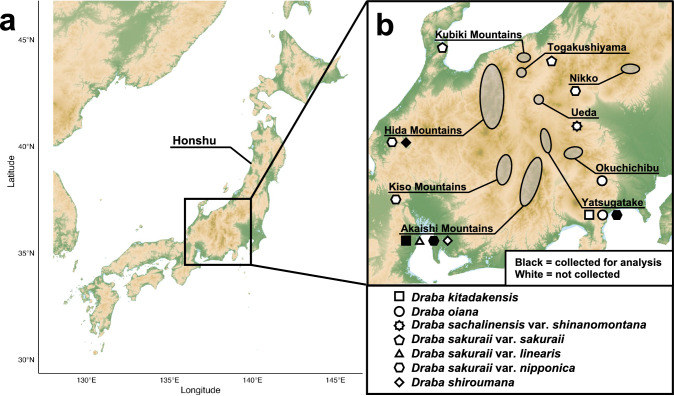 Figure 1
Figure 1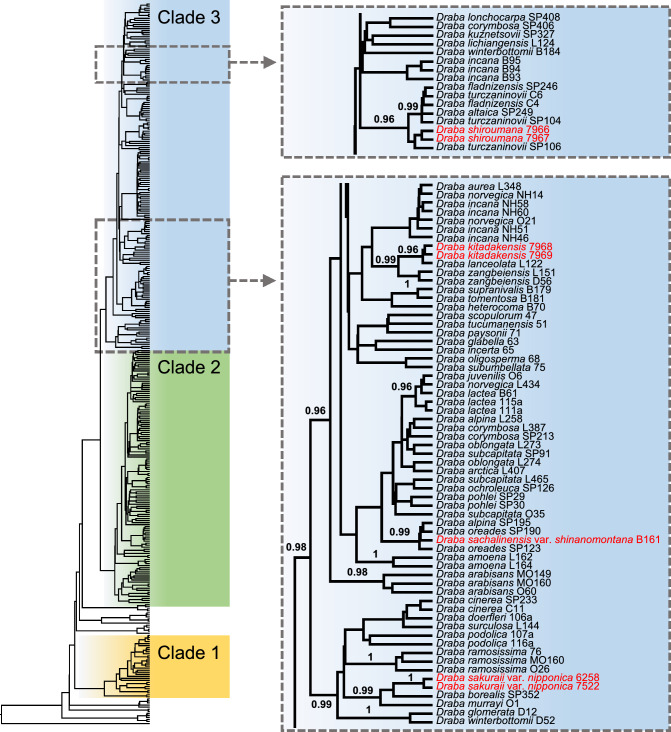 Figure 2
Figure 2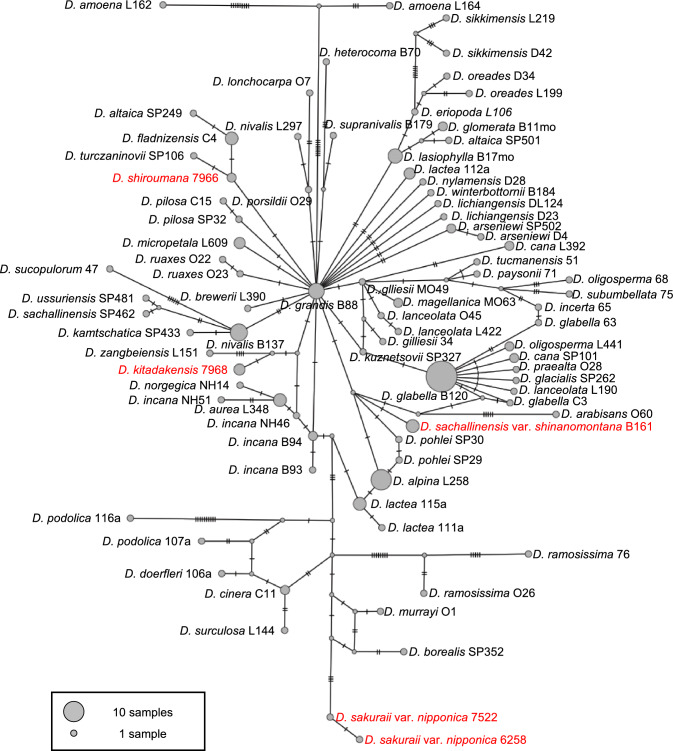 Figure 3
Figure 3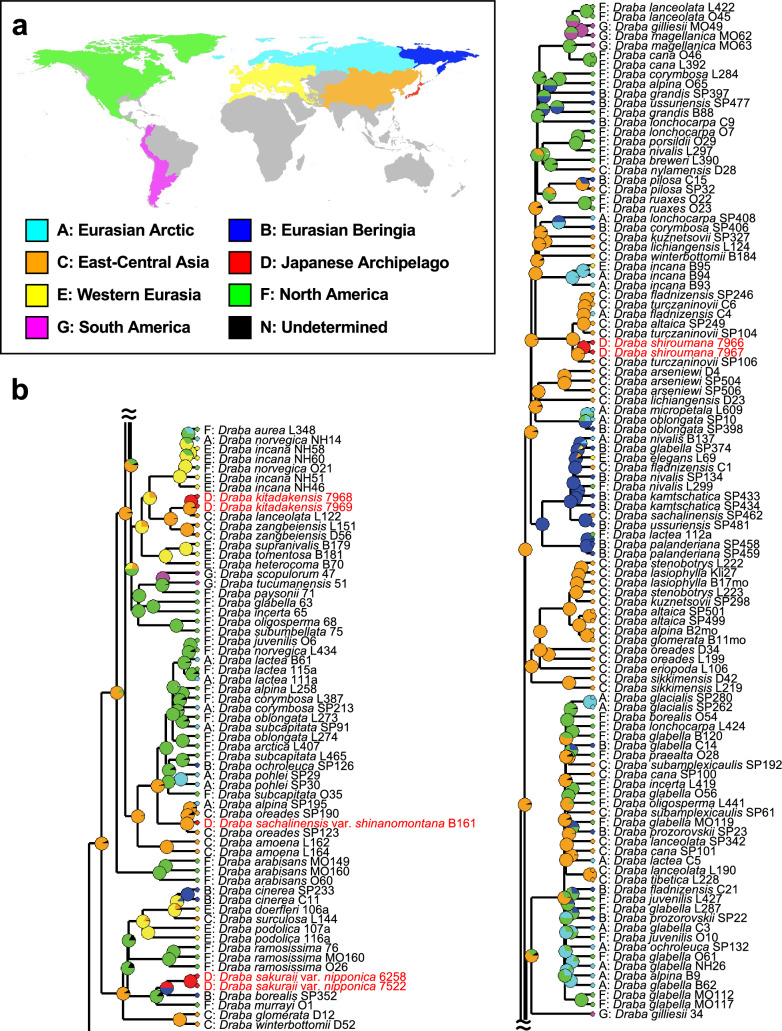 Figure 4
Figure 4- —http://dx.doi.org/10.13039/501100001691Japan Society for the Promotion of Science
- —The University of Tokyo
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Ecology and Taxonomy Studies · Plant and Fungal Species Descriptions · Plant Diversity and Evolution
Introduction
Cold-adapted plants inhabiting high mountains, such as alpine plants, are renowned for their exceptionally high diversity of endemic species (Hughes and Atchison 2015). One scenario explaining this high diversity of endemic species is in situ diversification (radiation) (Rahbek et al. 2019; Rangel et al. 2018), a commonly observed pattern across various mountain ranges (e.g., Madagascar, Camacho et al. 2021; European Alps, Comes and Kadereit 2003; Rocky Mountains, Drummond et al. 2012; the Andes, Hughes and Eastwood 2006). This pattern of evolutionary history is largely driven by the “island-like” geographically isolated distribution of alpine zones (Hughes and Eastwood 2006), which limits the gene flow (Morgan and Venn 2017), and enhances genetic differentiation (DeChaine and Martin 2005). In addition, complex topography in mountain regions provides mosaic environmental conditions, which lead to local adaptation and subsequent divergence of endemic species (DeChaine et al. 2014). Thus, endemic species in a particular mountain range will form monophyly under this evolutionary history (Hughes and Eastwood 2006). On the other hand, endemic species may diverge from phylogenetically independent lineages (non-radiation). In this case, each species originated through migrations—including long-distance dispersal—from multiple source areas and/or independent adaptation events, resulting in polyphyletic relationships (e.g., Dagallier et al. 2020; Salomón et al. 2022; Smyčka et al. 2022). Since the evolutionary history of endemic species diversity was limited to studies in major mountain ranges (e.g. the Andes, Rocky Mountains, and European Alps; see Hughes and Atchison 2015 for a comprehensive review), it remains to be explored which of these evolutionary histories could explain the high endemism in mountain ranges with limited geographic extension.
The high mountains of the Japanese Archipelago host species-rich alpine flora, of which approximately 51.3% of its alpine flora is endemic to the archipelago, making it a significant hotspot of endemic species (Kato and Ebihara 2011; Shimizu 1982, 1983). Given that nearly half of alpine endemics to the archipelago have closely related species distributed in the Arctic, northern Pacific (Beringia), northern Asia, and North America (Shimizu 1983), it implies that the ancestor of endemic species likely derived from cold-adapted plants distributed in these regions. Since the Japanese Archipelago had minimum glacial coverage during the glacial periods of the Pleistocene climatic oscillation (Tsukada 1983), cold-adapted plants could have migrated to the archipelago during these periods as far as central Honshu, the main island of the Japanese Archipelago (Ikeda 2022; but see Ikeda et al. 2014 for exception), and accumulated unique genetic divergence in numerous alpine plants (Fujii and Senni 2006; Ikeda 2022). Moreover, the alpine zone of central Honshu harbors several genera containing multiple endemic species, for example, Leontopodium (Pers.) R.Br. ex Cass., Taraxacum F. H. Wigg., and Ranunculus L. (Kato and Ebihara 2011; Shimizu 1982, 1983). However, the evolutionary history of high endemic species diversity on high mountains in the Japanese Archipelago has never been explored.
Draba Dill. ex L. (Brassicaceae) is one of the most species-rich genera (ca. 390 species) that occur throughout the major mountain systems and arctic tundra around the world (Jordon-Thaden and Koch 2008; Jordon-Thaden et al. 2010, 2013). Draba provide an appropriate study system to explore the evolutionary origin of the high endemic species diversity in mountain flora. Although some species are widely distributed in circum-polar areas and temperate mountains, the majority of species are endemic to narrow geographical regions (Jordon-Thaden et al. 2013), representing a significant proportion of total species richness in the mountainous flora (Figueroa et al. 2022; Jordon-Thaden et al. 2013). Regardless of the high diversity of endemic species in this genus, a previous study investigated phylogenetic relationships among ca. 43% of 169 species in Draba using both plastid (trnL-F) and nuclear (ITS) DNA sequences (Jordon-Thaden et al. 2010), providing a framework for further phylogenetic comparisons (Kucs et al. 2021).
In the alpine zone of the Japanese Archipelago, Draba is the most endemic species-rich genus containing11 endemic taxa (Kadota 2016; Kato and Ebihara 2011) of which seven occur in the mountains of central Honshu (Kadota 2016): Draba kitadakensis Koidz. found in the Akaishi Mountains and Yatsugatake; Draba oiana Honda, restricted to Okuchichibu and Yatsugatake; Draba sachalinensis (F. Schmidt) Trautv. var. shinanomontana (Ohwi) Okuyama occurring only in Ueda, Nagano Prefecture; Draba sakuraii Makino var. sakuraii, confined to the Kubiki Mountains and Togakushiyama; Draba sakuraii var. linearis (Kitam.) Sugim., found exclusively in the Akaishi Mountains; Draba sakuraii var. nipponica (Makino) Takeda, sporadically occurring across Nikko, the Hida Mountains, Kiso Mountains, Akaishi Mountains, and Yatsugatake; Draba shiroumana Makino, endemic to the Hida and Akaishi Mountains (Fig. 1; Kadota 2016). Of these seven endemic taxa, only D. sachalinensis var. shinanomontana has been analyzed in the previous phylogenetic study (accession B161 corresponding to the herbarium voucher B100127351, Curator Herbarium B 2000 in Jordon-Thaden et al. 2010). Among the remaining taxa, three varieties of D. sakuraii differ only in their hair density (Kadota 2016), while D. oiana is distinguished from D. kitadakensis only in the number of stem leaves and style length (Kadota 2016), and is often considered as synonym with D. kitadakensis (Kato and Ebihara 2011; Ohwi 1965). According to this classification, the phylogenetic relationships among the three robust species, D. sakuraii, D. kitadakensis, and D. shiroumana, may inform about the evolutionary history of the endemic Draba to central Honshu.Fig. 1. Topological map of a the Japanese Archipelago and b distribution of endemic Draba in central Honshu. Black and white shapes indicate whether samples were collected for the present study or not
In this study, we aim to clarify the evolutionary history that led to the high diversity of endemic species in the alpine zone of the Japanese Archipelago, through unraveling the phylogenetic relationship of the endemic taxa of Draba. We newly sequence the plastid (trnL-F) and nuclear (ITS) DNA of the three species D. kitadakensis, D. sakuraii var. nipponica, and D. shiroumana, and investigate their phylogenetic relationships with other Draba species covered in the previous study (Jordon-Thaden et al. 2010). Furthermore, we reconstruct the ancestral area for each endemic taxa to infer their phylogeographical origins. Through these analyses, we examine whether the endemic Draba to central Honshu have a radiative or non-radiative origin. If these endemic taxa have a radiative origin, a monophyletic relationship among these endemic species is expected. Alternatively, if these endemic taxa have a non-radiative origin, they would present polyphyletic relationships, each being closely related to species outside the Japanese Archipelago.
Materials and methods
Chloroplast and nuclear DNA sequencing
Leaf material of three endemic taxa of Draba were collected from the following sites: D. kitadakensis from Kitadake (Akaishi Mountains), D. sakuraii var. nipponica from Yatsugatake and Kitadake, and D. shiroumana from Shiroumadake (Hida Mountains) (Table 1). DNA of two samples of each endemic taxon was extracted from a silica gel dried sample using DNeasy Plant Mini Kit (Qiagen, Hilden, Germany). The intron and intergenic spacer of chloroplast and nuclear regions were amplified and sequenced. For the chloroplast DNA, the trnL–F region was obtained through the separate amplification of the trnL introns and the trnL–F intergenic spacer using the following primers: trnL intron forward primer 5′-CGA AAT CGG TAG ACG CTA CG-3′; trnL intron reverse primer 5′-GGG GAT AGA GGG ACT TGA AC-3′; intergenic spacer forward primer 5′-CGA AAT CGG TAG ACG CTA CG-3′; intergenic spacer reverse primer 5′-ATT GAA CTG GTG ACA CGA G-3′ (Taberlet et al. 1991). For the nuclear DNA, the internal transcribed spacer (ITS) region of nuclear ribosomal DNA including ITS1, 5.8S, and ITS2 were amplified using the ITS-18 forward primer 5′-GCA TGT TTT CCC AGT CAC GAC GGA AGG AGA AGT CGT AAC AAG G-3′ and the ITS-25 reverse primer 5′-ACT TCA GGA AAG AGC TAT GAC GGG TAA TCC CGC CTG ACC TGG-3′. The PCR amplifications were performed using fast reaction polymerase (SapphireAmp® Fast PCR Master Mix; TaKaRa BioInc., Shiga, Japan), following the manufacturer’s protocol, in which thermal cycling condition involved initial denaturation at 94 °C for 1 min, followed by 30 cycles at 98 °C for 5 s, 55 °C for 5 s, 72 °C for 10 s, and a final extension at 72 °C for 5 min. These amplicons were purified using ExoSAP-IT (USB Corporation, Ohio, USA) and used as direct sequencing templates. Cycle sequencing was conducted with a BigDye Terminator 3.1 cycle sequencing kit (Applied Biosystems, Foster City, California, USA), and the reaction mixture was sequenced using an ABI 3130 Genetic Analyzer (POP-7 polymer and 50-cm capillary; Applied Biosystems, Foster City, CA).Table 1. Plant materials collected in this studyTaxon and localityLatitudeLongitudeSample IDDraba kitadakensis Koidz Kitadake, Akaishi Mountains, Yamanashi Prefecture35.70138.27968 Kitadake, Akaishi Mountains, Yamanashi Prefecture35.70138.27969Draba sakuraii Makino var. nipponica Makino Yatsugatake Mountains, Nagano Prefecture35.98138.376258 Kitadake, Akaishi Mountains, Yamanashi Prefecture35.68138.247522Draba shiroumana Makino Shiroumadake, Hida Mountains, Nagano Prefecture36.79137.797966 Shiroumadake, Hida Mountains, Nagano Prefecture36.79137.797967
Phylogenetic analysis and haplotype network
Chromatograms of obtained sequenced data were manually checked and the contig was created using ChromasPro 2.1.10.1 and combined with published trnL–F and ITS sequences used in Jordon-Thaden et al. (2010). In order to reduce the bias in subsequent phylogenetic analysis, we retained only the core Draba (as defined in Jordon-Thaden et al. 2010) and Pseudoturris turrita (L.) Al-Shehbaz, a sister taxa to Draba (Jordon-Thaden et al. 2010), as the outgroup. To account for the polyphyletic nature of several species, potentially due to the existence of cryptic species (Jordon-Thaden et al. 2010), we treated each sample as a distinct molecular lineage, in which each lineage was identified by its species epithet and internal sample ID (see Table S1 in Supplementary Information). Subsequently, all the sequences were aligned using MUSCLE (Edgar 2004), built in AliView (Larsson 2014).
For phylogenetic analyses, each sequence dataset was analyzed by maximum likelihood (ML) using RAxML-NG 1.2.0 (Kozlov et al. 2019) and Bayesian Inference (BI) using BEAST 2 2.7.6 (Bouckaert et al. 2014). MEGA11 (Tamura et al. 2021) was used to select the best-fitting substitution model for each DNA region, and the best-fit model was chosen based on the Akaike information criterion (AIC) (Akaike 1974). For ML analysis, the bootstrap supports (BS) were assessed using 1,000 bootstrap pseudo-replicates. For BI analysis, four simultaneous Monte Carlo Markov chains (MCMCs) were run for 1.0 × 10^8^ generations, and trees were sampled every 1,000 generations. The first 10% were discarded as burn-in.
For the haplotype network, a subset of sequence data was created based on the phylogenetic tree constructed in the previous step. Samples belonging to a clade included all endemic species in central Honshu and with strong support (either BS > 80 or PP > 0.95) were used. The network was constructed using the TCS network method conforming to POPART (Leigh and Bryant 2015).
Ancestral area reconstruction
To reconstruct the ancestral distribution of endemic Draba taxa to central Honshu, we defined seven geographical regions: (A) Eurasian Arctic, encompassing Arctic Asia, Lappland, coastal Norway, and the North Atlantic Islands; (B) Eurasian Beringia, encompassing Chukchi/Kamchatka, Wrangel Island, Cherskii/Kolyma Mountain/Tundra, and the Kuril Islands; (C) East-Central Asia, encompassing of the Greater Asian Mountains, along with the Korean Peninsula and Okhotsk; (D) Japanese Archipelago; (E) Western Eurasia, encompassing the Mediterranean, Central Europe, Turkey/Caucasus/Iran, and Atlantic European forests/grasslands; (F) North America, encompassing North American Beringia, the North American Cordillera, Eastern North America, and Greenland; and (G) South America. This classification basically depends on ecoregions that explain species composition of Draba (Jordon-Thaden et al. 2013), whereas we designated the Japanese Archipelago as an independent geographic area. This classification aligns with defining Beringia as extending from west of the Lena River to east of the Mackenzie River (Abbott and Brochmann 2003), thereby excluding both the Korean Peninsula and Okhotsk. To prevent overestimation of the distribution range for species containing potential cryptic species, each lineage was assigned a single geographical region described above based on the geographical coordinates of each sample provided by Jordon-Thaden et al. (2010). The best consensus tree topology and 1,000 randomly sampled trees of BI phylogeny from prior analyses were then used to examine a likelihood-based biographical model in BioGeoBears (Matzke 2013) implemented in RASP 4.3 (Yu et al. 2020). The model test for BioGeoBears was conducted using three different models: DEC, DIVALIKE, and BAYAREALIKE (Matzke 2013). Despite the ongoing controversy over the jump dispersal (+ J) model (Matzke 2014, 2022; Ree and Sanmartín 2018), we also tested this model, considering the island-like nature of the alpine zone (DeChaine and Martin 2005). The best-fit model was chosen based on the corrected AIC (AICc).
Results
Phylogenetic analysis
The genus-wide trnL-F dataset contained 351 samples comprising 157 taxa, and their sequences consisted of a total of 907 bp with 299 variable sites and 175 parsimony informative sites. Besides, the ITS dataset contained 352 samples comprising 157 taxa, and their sequences consisted of a total of 625 bp with 265 variable sites and 175 parsimony informative sites. The best-fit substitution model was selected as GTR + G + I for both trnL-F and ITS regions. The phylogenetic trees constructed using trnL–F showed low interspecific polymorphism, while trees based on ITS (Figs. S1, S2) showed greater interspecific variation. Since many branches displaying near-zero lengths in trnL–F (Figs. S3, S4) unlikely reflect divergent history among species, we solely used ITS trees for further analyses.
The BI tree constructed from the ITS supported three distinct clades with high posterior probabilities (PPs > 0.95) (Fig. 2). Clade 1 primarily comprised taxa distributed across Western Eurasia, while Clade 2 was dominated by taxa from East-Central Asia, North America, and South America. Clade 3 included taxa predominantly from the Eurasian Arctic, Eurasian Beringia, East-Central Asia, and North America. All endemic taxa from central Honshu belonged to Clade 3 (Fig. 2). While two samples of each D. kitadakensis, D. sakuraii var. nipponica, and D. shiroumana formed monophyly per taxa, and there was not a monophyletic relationship among the four endemic species (Fig. 2). In particular, two clades strongly supported a polyphyletic relationship, one clade containing D. sakuraii var. nipponica (PP = 0.99) and another clade containing D. kitadakensis, D. sachalinensis var. shinanomontana, and D. shiroumana (PP = 0.96). The sister lineage for each endemic taxon was estimated as follows: D. kitadakensis with D. lanceolata Royle (L122; central China; PP = 0.96), D. sachalinensis var. shinanomontana with D. oreades Schrenk (SP190; the Altay Mountains) + D. alpina (SP195; the Polar Urals) (PP = 0.99), D. sakuraii var. nipponica with D. borealis D.C. (SP352; Kunashir Island in the Kuril Islands; PP = 0.86); D. shiroumana with D. turczaninowii Pohle & N. Busch (SP106; East Sayan; PP = 0.57) (Fig. 2).Fig. 2. Bayesian tree based on ITS. Three highly supported clades (PP ≥ 0.95) are highlighted. Two topologies, detailing the relationships of endemic taxa from central Honshu and their closely related taxa, are shown. Endemic taxa from central Honshu are indicated with colored text. Posterior probability support values (PP ≥ 0.95) are shown
Haplotype network
The haplotype network was constructed with samples belonging to clade 3 in the phylogenetic tree (Figs. 2, 3). Among the three endemic taxa that included more than one sample, D. sakuraii var. nipponica was the only polymorphic, while others (D. kitadakensis and D. shiroumana) had no nucleotide polymorphisms (Fig. 3). In congruence with the relationships observed in the phylogenetic tree (Fig. 2), D. kitadakensis was found to share a haplotype with D. lanceolata (L122), D. sachalinensis var. shinanomontana shared a haplotype with D. oreades Schrenk (SP123; SP190) and D. alpina (SP195), while D. sakuraii var. nipponica differed from D. borealis (SP352) by 5 to 6 nucleotides, and D. shiroumana differed from D. turczaninowii (SP106) by one nucleotide (Fig. 3).Fig. 3. Haplotype network of Draba belonging to clade 3 from the Bayesian tree based on ITS. Each tick on the line represents a nucleotide polymorphism, and the circumference of each circle is proportional to the number of taxa with no nucleotide polymorphism. Endemic taxa from central Honshu are indicated with colored text
Reconstruction of ancestral area
Ancestral area reconstruction was performed using the best-fitting model, such as the BAYAREALIKE + J model (Table 2). Ancestral area reconstruction estimated that clade 1 originated from Western Eurasia (E) (ancestral area likelihood [AAL] = 100.0%; Fig. S5), and clades 2 and 3 originated from East-Central Asia (C) (AAL = 99.7% and 92.3%, respectively; Figs. 4, S5). The ancestral area(s) of the endemic taxa from central Honshu was reconstructed as the following: D. kitadakensis and its sister lineage, D. lanceolata (L122), was East-Central Asia (C) (AAL = 94.8%); D. sachalinensis var. shinanomontana and its sister lineage, D. oreades (SP190) and D. alpina (SP195), was East-Central Asia (C) (AAL = 90.8%); D. sakuraii var. nipponica and its sister lineage, D. borealis (SP352), was Eurasian Beringia (B) + Japanese Archipelago (D) (AAL = 50.4% and 49.6%, respectively); and D. shiroumana and its sister lineage, D. turczaninowii (SP106), was East-Central Asia (C) (AAL = 96.9%) (Fig. 4).Table 2. Log-likelihood (LnL) and corrected Akaike information criterion (AICc) of the six different models tested using BioGeoBEARSModelsLnLAICcBAYAREALIKE + J− 411.6829.3DEC + J− 411.7829.5DIVALIKE + J− 412.6831.2DEC− 483969.2DIVALIKE− 494.1992.2BAYAREALIKE− 508.61021Fig. 4Ancestral area reconstruction of the Draba belonged to clade 3 from the Bayesian tree based on ITS. a Distribution map of the seven geographical regions used for reconstruction. b Estimated ancestral areas and likelihoods inferred using BioGeoBEARS under best-fitting model BAYAREALIKE + J. The alphabet beside taxonomic name represents the geographical region used for reconstruction. Endemic taxa from central Honshu are indicated with colored text
Discussion
The present study revealed the four endemic taxa of Draba to high mountains in central Honshu—D. kitadakensis, D. sachalinensis var. shinanomontana, D. sakuraii var. nipponica, and D. shiroumana—are not monophyletic (Fig. 2). Even though the phylogenetic trees have little resolution for relationships among closely related species due to generally low branch supports (< 0.95), D. sakuraii var. nipponica and the remaining three taxa (D. kitadakensis, D. sachalinensis var. shinanomontana, and D. shiroumana) belonged to different clades with high support values (PP = 0.99 and 0.96, respectively) (Figs. 2, S2). This finding implies that endemic taxa of Draba in central Honshu originated from at least two different evolutionary lineages. Consequently, our study suggests that the evolutionary history associated with richness of the endemic Draba in central Honshu is compatible with the idea of a non-radiative origin, instead of diversifying from a single common ancestor as expected in cases denoting a radiative origin (Hughes and Eastwood 2006).
We acknowledge several cautions in our phylogenetic analyses due to the low resolution in molecular data (Jordon-Thaden et al. 2010; Kucs et al. 2021). In particular, polyphyletic relationships may represent the lack of phylogenetic information in the sequence data due to random sorting of ancestral polymorphisms during rapid speciation. However, the phylogenetic relationships mostly exhibited geographic patterns (Fig. 4), which unlikely resulted from random sorting of polymorphisms (Goodwillie and Stiller 2001; Ikeda 2022). Thus, the independent lineage of D. sakuraii var. nipponica from the remaining species would represent a robust evolutionary history. On the contrary, the low resolution of our phylogenetic trees prevents us from leading to the non-radiative origin of D. kitadakensis, D. sachalinensis var. shinanomontana, and D. shiroumana, even though they grouped with different species (Fig. 4).
In addition, it should be noted that the present study did not analyze all taxa endemic to central Honshu. Given the morphological similarity of D. sakuraii var. linearis and D. sakuraii var. sakuraii to D. sakuraii var. nipponica and D. oiana to D. kitadakensis (Kadota 2016; Kato and Ebihara 2011), the unanalyzed species may be closely related to the species analyzed in this study. Although some of their diversification may reflect a radiative origin, their radiative origin could not exclude the present finding that endemic Draba originated from at least two independent lineages.
Considering the evolutionary history noted in the non-radiative origin of alpine endemics to major mountain ranges (e.g., Dagallier et al. 2020; Salomón et al. 2022; Smyčka et al. 2022), the high endemism of Draba in the Japanese Archipelago could be explained by a similar scenario to the divergence of Helichrysum Mill. (Asteraceae) in the Drakensberg, the southernmost alpine zone in Africa (Blanco-Gavaldà et al. 2023). In Helichrysum, 12 species are endemic to the Drakensberg, which belong to at least seven distinct lineages. In particular, four lineages originated from nearby species-rich regions, three from the southern African grasslands (which harbor 98 species) and the one from the Fynbos biome (which contains 12 species) (Blanco-Gavaldà et al. 2023). This divergent history implies that multiple nearby regions with high species diversity may be associated with local high endemism, where migration of distantly related lineages from multiple sources contributes to endemic species with non-radiative origin within the alpine zone (Dagallier et al. 2020). In the case of the endemic Draba, the Japanese Archipelago is located nearby areas with high species diversity such as the Eurasian Arctic, Eurasian Beringia, and East-Central Asia, with 40, 43, and 99 species, respectively (Jordon-Thaden et al. 2013). The present polyphyletic relationships among the endemic taxa of Draba (Fig. 2), and different geographical origins estimated as their ancestral area (Fig. 4) suggest migrations from multiple regions, implying non-radiative origin of the high endemism of Draba in the Japanese Archipelago. Given that almost half of the endemic species in the alpine zone in the Japanese Archipelago are related to species distributed in the Arctic, northern Pacific, and northern Asia (Shimizu 1983), multiple endemic species in other alpine plant genera, such as Leontopodium, Taraxacum, and Ranunculus, may have non-radiative origin from these species-rich regions.
Conclusion
We showed that endemic taxa of Draba to central Honshu originated from at least two evolutionary independent lineages, suggesting that their divergence occurred in a non-radiative manner. Our finding in the alpine zone of the Japanese Archipelago did not align with the radiative origin of endemic species diversity commonly observed in several major mountain ranges (Hughes and Atchison 2015). In contrast, the present non-radiative origin is explained by a similar scenario with the previous studies (Blanco-Gavaldà et al. 2023; Dagallier et al. 2020); multiple source regions with high species diversity led to the high endemism of Draba in the Japanese Archipelago.
Supplementary Information
Below is the link to the electronic supplementary material.Supplementary file1 (PDF 1501 KB)Supplementary file2 (XLSX 39 KB)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Curators Herbarium B (2000) Digital specimen images at the Herbarium Berolinense [Draba sachalinense B 10 012735 1]. Botanic Garden and Botanical Museum Berlin. https://herbarium.bgbm.org/object/B 100127351. Accessed 17 Nov 2024
- 2Kadota Y (2016) Draba. In: Ohashi H, Kadota Y. Kihara H, Murata J, Yonekura K (eds) Wild flowers of Japan 4. Heibonsha, Tokyo, pp 60–63 (in Japanese)