Fatty Acids and Inflammatory Protein Biomarkers From Coronavirus Disease 2019 Patients
Niharika Bala, Alaa H. Habib, Marianne Kozuch, Nancy D. Denslow, Neha S. Dhaliwal, Anna H. Owings, Sarah C. Glover, Abdel A. Alli

TL;DR
This study identifies changes in fatty acids and inflammatory proteins in the blood of COVID-19 patients, which could help assess disease severity and treatment options.
Contribution
The study establishes a unique plasma signature of fatty acids and inflammatory biomarkers specific to COVID-19 patients.
Findings
Fatty acid levels like arachidic and myristic acid decreased in COVID-19 patients.
Chemokines like IP-10, MCP-1, and cytokines like IL-1 alpha were elevated in COVID-19 patients.
Inflammatory markers such as ICAM-1 and E-selectin were higher in the plasma of infected individuals.
Abstract
The coronavirus disease 2019 (COVID‐19) is a viral infection caused by severe acute respiratory syndrome coronavirus 2 (SARS‐CoV‐2) and associated with systemic inflammation. Inflammation is an important process that follows infection and facilitates the body's innate immune response and repair of damaged tissue. Polyunsaturated fatty acids play an important role in the inflammatory process. These lipids can target transcription factors to modulate gene expression and protein function. Here, we performed a fatty acid methyl ester (FAME) analysis and immunoassays to evaluate whether differences in basal levels of different types of biomarkers can be detected in freshly frozen plasma samples from patients with and without COVID‐19. FAME analysis showed a decrease in arachidic acid and myristic acid, but an increase in caprylic acid, palmitic acid, and eicosenoic acid in the plasma of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
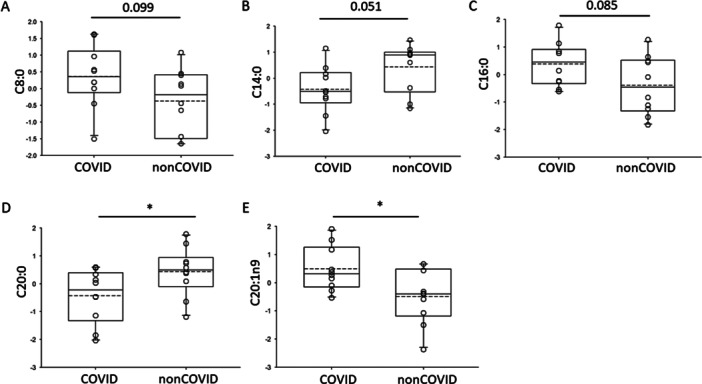 Figure 1
Figure 1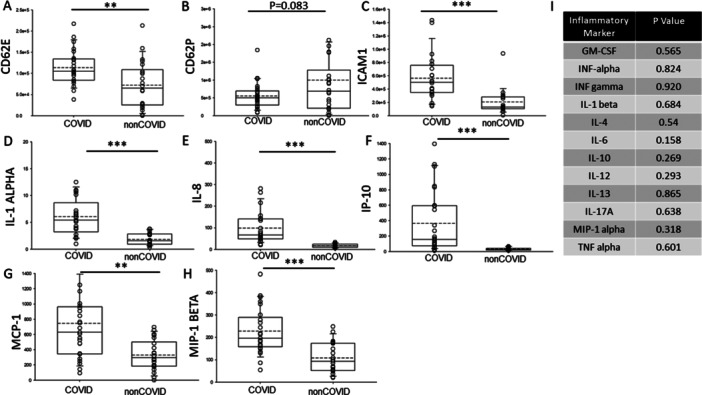 Figure 2
Figure 2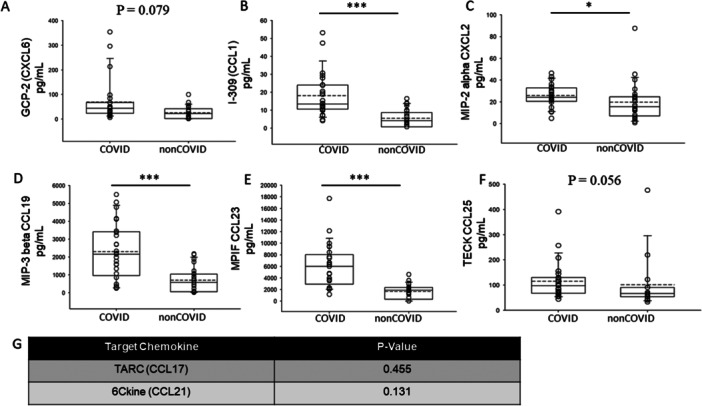 Figure 3
Figure 3| Analyte | Bead number |
|---|---|
| 6Ckine | 46 |
| GCP‐2 | 64 |
| I‐309 | 75 |
| MCP‐4 | 73 |
| MIP‐2 alpha | 56 |
| MIP‐3 beta | 74 |
| MPIF | 53 |
| TARC | 30 |
| TECK | 67 |
| Marker | Biomarker for | |
|---|---|---|
| CD62E/E‐selectin | Critical disease and increased risk of ICU admission | Mezine et al. [ |
| ICAM‐1 | Elevated in patients with mild and severe cases | Tong et al. [ |
| IL‐1 | Heightened inflammatory response hallmark of severe COVID‐19 | Chang et al. [ |
| IL‐8 | Found elevated in ICU patients | Chang et al. [ |
| MCP‐1 | Thrombosis‐related indicator | Chen et al. [ |
| IP‐10 | Thrombosis‐related indicator | Chen et al. [ |
| MIP‐1 beta | Mild to moderate COVID severity marker | Codina et al. [ |
| CXCL‐2 | Chronic inflammation and fibrosis | Scott et al. [ |
| Analyte | Bead number |
|---|---|
| GM‐CSF | 44 |
| IFN alpha | 48 |
| IFN gamma | 43 |
| IL‐1 alpha | 62 |
| IL‐1 beta | 18 |
| IL‐4 | 20 |
| IL‐6 | 25 |
| IL‐8 | 27 |
| IL‐10 | 28 |
| IL‐12p70 | 34 |
| IL‐13 | 35 |
| IL‐17 | 36 |
| TNF alpha | 45 |
| IP‐10 | 22 |
| MCP1 | 51 |
| MIP‐1 alpha | 12 |
| MIP‐1 beta | 47 |
| ICAM‐1 | 73 |
| CD62E | 77 |
| CD62P | 55 |
- —This study was funded by an NIH R01 Grant R01DE031928‐01A1 (to S.C.G.) and foundation funds from the University of Mississippi Medical Center.
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCOVID-19 Clinical Research Studies · Long-Term Effects of COVID-19 · SARS-CoV-2 and COVID-19 Research
Introduction
1
Fatty acids play a key role in signal transduction and in the production of mediators involved in inflammatory pathways associated with viral infection. Polyunsaturated fatty acids are known to regulate lipid raft formation associated with SARS‐CoV‐2 entry mechanisms including transmembrane protease serine‐2 (TMPRSS2) and angiotensin‐converting enzyme‐2 (ACE2) [1]. The overall effect of an inflammatory response is dependent on the interplay between pro‐ and anti‐inflammatory mediators. Pro‐inflammatory cytokines are involved in early responses and amplify inflammatory processes, while anti‐inflammatory cytokines have the opposite effect. Several studies have reported that cytokines are released into the bloodstream after severe COVID‐19 and the resulting cytokine storm can be a consequence of different mechanisms. Hirano and Murakami suggested that one possible mechanism of the cytokine storm in COVID‐19 is mediated by nuclear factor‐κB (NF‐κB) activation, reduction in ACE2 expression, and a subsequent increase in angiotensin 2 [2]. Numerous strategies have been employed to control the cytokine storm and improve the prognosis and survival of the patients. These therapies include cytokine antagonists [3, 4] and immunomodulators [5]. There have been clinical studies performed involving immunomodulatory approaches to combat lung inflammation induced by cytokine storm [6]. Additionally, studies involving the use of small‐molecule‐based medications to enhance regulatory T cell (Tregs) activity to mitigate the cytokine storm due to severe viral infection have been described by Dhawan et al. [7].
Previous studies have shown that oxidative stress induced during COVID‐19 is associated with changes in lipid profiles and systemic inflammation [8]. The inability to resolve inflammatory responses and/or the persistent activation of the inflammasome can trigger a cascade of deleterious outcomes on multiple organ systems. Free fatty acids or nonesterified fatty acids are important in human pathophysiology and have been found to positively or negatively correlate with various diseases including obesity [9], insulin resistance [10], diabetes mellitus [11], metabolic syndrome [12], diabetic nephropathy [13], cardiovascular disease [14], and cerebrovascular accident [15].
The primary goal of this study was to first evaluate whether the levels of different types of circulating biomarkers from COVID‐19 patients compared to non‐COVID‐19 patients can be determined from freshly frozen plasma samples. Secondary outcomes of this study were to examine putative correlations between these circulating fatty acids, cytokines, and chemokines in the context of COVID‐19 pathophysiology.
Materials and Methods
2
Inclusion and Exclusion Criteria
2.1
The inclusion and exclusion criteria for this study were similar to a previous study by Ziegler et al. [16]. Briefly, participants were enrolled from outpatient clinics, medical‐surgical units, intensive care units (ICU), or endoscopy units at the University of Mississippi Medical Center (UMMC) between April and September 2020. The UMMC Institutional Review Board approved the study under IRB#2020‐0065. To qualify for the COVID‐19 group, individuals had to be at least 18 years old, test positive for SARS‐CoV‐2 through a nasopharyngeal PCR swab, present with COVID‐19 symptoms such as fever, chills, cough, shortness of breath, and sore throat, and have a body weight exceeding 110 pounds. The Control group consisted of individuals who were at least 18 years old, had a recent negative SARS‐CoV‐2 test (PCR or rapid antigen), and weighed more than 110 pounds. Table S1 gives the COVID‐19 variant, age, and race of each participant. Informed consent was provided by each participant or their legally authorized representative in both groups. Blood samples were collected by trained healthcare personnel using sodium heparin‐containing tubes. Plasma was obtained by centrifuging the tubes at 1200g for 10 min and transferring the supernatant to clean tubes for storage in a −20°C freezer.
Blood Plasma FAME Derivatization
2.2
Blood plasma samples were prepared for analysis with a direct derivatization technique as previously described [17]. Briefly, the samples were weighed in round bottom glass tubes, and 14% boron trifluoride in methanol was added (Millipore Sigma, Burlington, MA), before being incubated in a 100°C sand bath for 1 h. After cooling, Optima grade hexanes (Fisher Scientific, Hampton, NH) and ultrapure water (MilliQ, Millipore Sigma) were added to the samples, and the samples were shaken to allow the transfer of FAME into the hexane. Hexane extraction was repeated once, and Na_2_SO_4_ (Fisher Scientific) scavenged any remaining water from the hexane extracts. Sample extracts were reduced to 0.3 mL under a gentle stream of Nitrogen (N_2_) at 35°C and spiked with US‐108N mixed deuterated Polycyclic Aromatic Hydrocarbon (PAH) for use as an internal standard. Fatty acid methyl ester (FAME) analysis was carried out on an Agilent 7890B gas chromatograph (GC) coupled to an Agilent 7000 C triple quad mass spectrometer (MS). Running conditions for the GC were as follows: 1 µL of sample was injected into the GC through a 280°C inlet in splitless mode, and helium carrier gas was maintained at 1.2 mL/min at constant flow. Samples passed through a DB‐FATWAX capillary column (30 m length, 250 µm outer diameter, 0.25 µm film thickness, Agilent). The initial oven temperature was 60°C, which was held for 2 min, after which the temperature was ramped at a rate of 10°C/min to 180°C. After a 5‐min hold time, the temperature was ramped to 250°C. This final temperature was held for 3 min. Chromatographically separated sample components passed into the MS through a 280°C transfer line, and were ionized in EI mode. The MS source temperature was 280°C; Q1 quadrupole temperature was held at 150°C. Sample FAME was detected in full scan mode, and quantitated by comparison to pure FAME calibration dilutions 0.01–5 µg/mL (NuChek Prep, Elysian, MN). C14:0C16:0C16:1n9C18:0C18:1n9Sample LOD0.0650.0650.0650.0650.065Sample LOQ0.5090.2540.5090.5090.509John Wiley & Sons, Ltd.
Human Chemokine Panel 9 Plex
2.3
Human COVID‐19 and non‐COVID‐19 plasma samples were centrifuged at 1000 g for 10 min at room temperature. Standards and the reagents were freshly prepared before the experiment. Magplex capture beads were added into each well followed by standards and samples. The assay was performed following the manufacturer's manual (Cat No. EPX090‐15840‐90, ThermoFisher Scientific, USA). A Luminex 200 instrument was used to read the plate followed by data processing in ThermoFisher cloud Procartaplex analysis app. The analytes detected included 6Ckine, GCP‐2, I‐309, MCP‐4, MPIF, MIP‐2 alpha, MIP‐3 beta, TARC, TECK as indicated in Table 1. Sigmaplot 15 software was used to perform a Student's t‐test and to plot the data (Table 1). The correlation of these inflammatory protein biomarkers with COVID‐19 and associated comorbidities is given in Table 2.
Human Inflammation Panel 20 Plex
2.4
Human COVID‐19 and non‐COVID‐19 plasma samples were centrifuged at 1000 g for 10 min at room temperature. Standards and reagents were freshly prepared before the experiment. Magplex capture beads were added into each well followed by standards and samples. The assay was performed following the manufacturer's manual (Cat No. EPX200‐12185‐901, ThermoFisher Scientific, USA). A Luminex 200 instrument was used to read the plate followed by data processing in ThermoFisher cloud Procartaplex analysis app. The cytokines detected included GM‐CSF, IFN alpha, IFN gamma, IL‐1 alpha, IL‐1 beta, IL‐4, IL‐6, IL‐8, IL‐10, IL‐12p70, IL‐13, IL‐17A, TNF alpha. The chemokines detected included IP‐10, MCP‐1, MIP‐1 alpha, MIP‐1 beta, and cell adhesion and inflammatory response markers ICAM‐1, CD62E (E‐selectin), CD62P (P‐Selectin) as noted in Table 3. Sigmaplot 15 software was used to perform a Student's t‐test and plot the data.
Results
3
Circulating Levels of Fatty Acids in Patients With or Without COVID‐19
3.1
Polyunsaturated fatty acids have been shown to play a role in SARS‐CoV‐2 entry and infection. Therefore, we investigated basal levels of prominent fatty acids in the systemic circulation of COVID‐19 patients compared to non‐COVID‐19 patients. Here, FAME analysis of freshly frozen plasma samples showed higher basal levels of caprylic acid (C8:0), palmitic acid (C16:0), and eicosenoic acid (C20:1n9) in COVID‐19 patients compared to non‐COVID‐19 patients (Figure 1). Conversely, basal levels of myristic acid (C:14:0) and arachidic acid (C20:0) were lower in the COVID‐19 group compared to the non‐COVID‐19 group (Figure 1).
Analysis of free fatty acids in plasma of patients with or without COVID‐19. (A) caprylic acid (C8:0), (B) myristic acid (C14:0), (C) palmitic acid (C16:0), (D) arachidic acid (C20:0), (E) eicosenoic acid (C20:1n9). * represents a p < 0.05.
Circulating Levels of Pro‐Inflammatory and Anti‐Inflammatory Cytokines in COVID‐19 and Non‐COVID‐19 Patients
3.2
Pro‐inflammatory cytokines play an important role in early responses and amplify inflammatory reactions while anti‐inflammatory cytokines limit the inflammatory reactions. Therefore, we measured both pro‐inflammatory and anti‐inflammatory cytokines in the plasma of COVID‐19 and non‐COVID‐19 patients. As shown in Figure 2, the levels of CD62E, ICAM1, IL‐1 alpha, IL‐8, IP‐10, MCP‐1, and MIP‐1 beta were augmented in the COVID‐19 group while CD62P was greater in the non‐COVID‐19 group.
Pro‐ and anti‐inflammatory cytokines detected in the plasma of COVID‐19 and non‐COVID‐19 patients. (A) CD62E, (B) CD62P, (C) ICAM1, (D) IL‐1 alpha, (E) IL‐8, (F) IP‐10, (G) MCP‐1, (H) MIP‐1 beta, (I) table showing inflammatory markers that were comparable between the two groups. ** represents a p < 0.01. *** represents a p < 0.001.
Circulating Levels of Chemokines in COVID‐19 and Non‐COVID‐19 Patients
3.3
Chemokines are a class of cytokines that are known to be expressed locally at sites of virally infected tissues [37]. Therefore, we measured the expression of various chemokines in the plasma of COVID‐19 and non‐COVID‐19 patients. As shown in Figure 3, several chemokines including GCP‐2, CCL1, MIP‐2 alpha CXCL2, MIP‐3 beta CCL19, MPIF CCL23, and TECK CCL25 were found to be significantly elevated in the COVID‐19 group compared to the non‐COVID‐19 group.
Expression of chemokines in the plasma of COVID‐19 and non‐COVID‐19 patients. (A) GCP‐2, (B) CCL1, (C) MIP‐2 alpha CXCL2, (D) MIP‐3 beta CCL19, (E) MPIF CCL23, (F) TECK CCL25, (G) table showing chemokines that were comparable between the two groups. * represents a p < 0.05, ** represents a p < 0.01. *** represents a p < 0.001.
Discussion
4
COVID‐19 remains a public health problem and the long‐term effects are still not known. In addition, there is a need to identify novel biomarkers from the systemic circulation that can be used to predict the severity of infection. Published studies suggest the use of never‐frozen plasma samples offers superior results when compared to the use of freshly frozen plasma samples [38]. In many cases, the recruitment of patients and collection of samples occurs over a period of months and the samples are frozen to be run at a later time. Running samples at the same time offers important advantages that includes achieving a desired sample size per study group, allowing for economic feasibility, and minimizing technical variation. Here, we determined whether statistically significant basal levels of multiple fatty acids, cytokines, and chemokines could be detected in freshly frozen plasma samples from patients with and without COVID‐19.
Fatty acids are important for the construction of complex lipids. These lipids can be synthesized by cells or they can be incorporated from extracellular sources. The quantification of long‐chain fatty acids in samples such as plasma represents a specific class of biomarkers present in the systemic circulation that can provide useful information including elucidating lipid pathways involved in pathophysiology. A study by Spiller and colleagues showed free fatty acids levels are elevated in newly diagnosed and long‐term controlled T2DM patients [11]. Liu and colleagues showed that docosahexaenoic acid negatively correlates with microalbuminuria in diabetic nephropathy [13]. Among the fatty acids that our FAME analysis showed decreased basal levels within the plasma of patients with COVID‐19 compared to patients without COVID‐19 was the 14‐carbon saturated fatty acid, myristic acid. The addition of myristic acid to the amino‐terminal glycine residue of proteins is known to stabilize proteins at the membrane and effect their cellular function. A study by Du and colleagues reported a reduction in myristic acid‐containing dimyristoyl‐PC and palmitoylmyristoyl‐PC in lung surfactant may promote SARS‐CoV‐2 infection and suggested increasing these lipids may be beneficial in treating severe COVID‐19 disease [39]. Importantly, a study by Rampoldi and colleagues showed amino‐terminal myristoylation is crucial for T cell receptor (TCR) signaling as well as the release of cytokines [40]. Another fatty acid that we found to be downregulated in the plasma of COVID‐19 patients compared to non‐COVID‐19 patients was arachidic acid. Fretts and colleagues reported higher levels of circulating arachidic acid are associated with a reduced risk of atrial fibrillation [41]. Circulating arachidic acid has been shown to have beneficial cardiovascular effects [42]. The results from our study suggest that the lower circulating levels of arachidic acid correlate with poor cardiovascular outcomes, which is observed in a large number of COVID‐19 patients. Caprylic acid, palmitic acid, and eicosenoic acid were among the three fatty acids that were found by our FAME analysis to be greater in the plasma of the COVID‐19 group compared to the non‐COVID‐19 group. The increased levels of palmitic acid in the plasma of COVID‐19 patients are consistent with data from published studies. Joshi and colleagues showed palmitic acid contributes to COVID‐19 pathogenesis by modulating molecules involved in the regulation of the immune system and apoptosis [43]. A nutrigenomic analysis study by Barh and colleagues suggested increased severity and higher death rate from COVID‐19 in western countries compared to that in India may be attributed to increased activation of cytokine storm related pathways, as a result of increased consumption of red meat and processed foods products rich in palmitic acid in the former [44]. Presumably, the ability of palmitic acid to induce endoplasmic reticulum stress, as shown by Ma et al. [45], and its ability to induce ACE2 expression in cultured hepatocytes, as shown by Cao et al. [46], may also contribute to increased morbidity and mortality associated with COVID‐19.
Prior studies have investigated the role of cholesterol‐enriched lipid rafts as signaling platforms for the entry of SARS‐CoV‐2 [47]. Lipid rafts do not only serve as binding sites for SARS‐CoV‐2, but they are also involved in the activation and internalization of the virus important for transmission from one cell to another [48]. Our group previously showed that extracellular vesicles isolated from the plasma of COVID‐19 patients increase the density of lipid rafts in human small airway epithelial cells [49]. A study by Teixeira and colleagues showed that the HMG CoA reductase inhibitor, simvastatin, disrupts lipid rafts and inhibits SARS‐CoV‐2 entry into cells and the production of inflammatory cytokines [50]. A study by El Khoury and Naim demonstrated that simvastatin and fluvastatin disrupt the interaction of ACE2 with the lipid rafts and its sorting at the brush border membrane of intestinal epithelial cells [51]. Bailly and Vergoten discussed the role of glycyrrhizic acid in the disorganization of the lipid rafts to inhibit coronavirus entry into cells [52]. A study by Alboni and colleagues showed that depletion of membrane cholesterol from hydroxypropyl‐beta‐cyclodextrin inhibits the entry of SARS‐CoV‐2 into human ACE2 overexpressing cells [53]. Multiple groups have suggested targeting lipid rafts may be effective in the treatment of COVID‐19 as it has been previously used to successfully treat other viral infections [48, 54, 55].
Chemokines play an important role in the recruitment of lymphocytes and leukocytes to injured tissue. They trigger biological activities through the activation of G protein‐coupled chemokine receptors. Some of these receptors recognize multiple chemokines and various chemokines bind to multiple receptors [56]. Liu and colleagues showed CCL1 promotes pulmonary fibrosis development through the activation of the AMFR–Ras–ERK–p70S6K signaling pathway [57]. Our study showed basal levels of multiple chemokines including CCL1 were significantly higher in the plasma of COVID‐19 patients when compared to levels in the plasma of non‐COVID‐19 patients.
Some studies suggest that using freshly obtained and never‐frozen plasma samples are necessary to obtain statistically significant results for the identification of biomarkers. Our study demonstrates that the use of stored plasma samples from patients with or without COVID‐19 can be used to successfully assay different types of biomarkers including unsaturated fatty acids, cytokines, and chemokines. We detected five or more biomarkers from each biomarker type and this data were consistent with expected results while considering similar data from other studies. Limitations of our study include not comparing fresh samples to freshly frozen samples, lack of inclusion of a healthy control group, and not comparing biomarker levels in the plasma from COVID‐19 and non‐COVID‐19 patients at different time points after viral infection. Nevertheless, the data presented here suggest that freshly frozen plasma samples are a rich source of circulating biomarkers that can differentiate between active COVID‐19 viral infection and non‐COVID‐19 disease. Finally, another limitation of our study is that we were unable to group patients based on their comorbidities. It is possible that some patients may have had other diseases that could contribute to the cytokine storm and changes in the different markers we investigated in this study.
Future studies may be performed to determine whether basal levels of these fatty acids, cytokines, and chemokines change in response to vaccination or recovery of COVID‐19. Additionally, it would be interesting to evaluate the levels of these systemic biomarkers in a prospective study aimed at studying the impact of long COVID‐19 associated with a decline in function of a specific organ system.
Author Contributions
Niharika Bala: data curation, formal analysis, investigation, writing – original draft, writing – review and editing. Alaa H. Habib: data curation, formal analysis, investigation, resources, writing – original draft, writing – review and editing. Marianne Kozuch: formal analysis, investigation, writing – original draft, writing – review and editing. Nancy D. Denslow: formal analysis, funding acquisition, supervision, writing – original draft, writing – review and editing. Neha S. Dhaliwal: investigation, writing – original draft, writing – review and editing. Anna H. Owings: investigation, writing – original draft, writing – review and editing. Sarah C. Glover: funding acquisition, resources, supervision, writing – original draft, writing – review and editing. Abdel A. Alli: conceptualization, formal analysis, investigation, project administration, resources, supervision, writing – original draft, writing – review and editing.
Ethics Statement
The UMMC Institutional Review Board approved the study under IRB#2020‐0065.
Consent
Written informed consent was obtained from all participants or their legally authorized representatives.
Supporting information
supmat.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1P. K. Baral , M. T. Amin , M. M. O. Rashid , and M. S. Hossain , “Assessment of Polyunsaturated Fatty Acids on COVID‐19‐Associated Risk Reduction,” Revista Brasileira de Farmacognosia 32, no. 1 (2022): 50–64.34876760 10.1007/s 43450-021-00213-x PMC 8638948 · doi ↗ · pubmed ↗
- 2T. Hirano and M. Murakami , “COVID‐19: A New Virus, but a Familiar Receptor and Cytokine Release Syndrome,” Immunity 52, no. 5 (2020): 731–733.32325025 10.1016/j.immuni.2020.04.003PMC 7175868 · doi ↗ · pubmed ↗
- 3M. Davidson , S. Menon , A. Chaimani , et al., “Interleukin‐1 Blocking Agents for Treating COVID‐19,” Cochrane Database of Systematic Reviews 1, no. 1 (2022): CD 015308.35080773 10.1002/14651858.CD 015308 PMC 8791232 · doi ↗ · pubmed ↗
- 4S. Murthy and T. C. Lee , “IL‐6 Blockade for COVID‐19: A Global Scientific Call to Arms,” Lancet Respiratory Medicine 9, no. 5 (2021): 438–440.33676592 10.1016/S 2213-2600(21)00127-2PMC 8078876 · doi ↗ · pubmed ↗
- 5Z. Mohseni Afshar , M. Barary , A. Babazadeh , et al., “The Role of Cytokines and Their Antagonists in the Treatment of COVID‐19 Patients,” Reviews in Medical Virology 33, no. 1 (2023): e 2372.35621229 10.1002/rmv.2372 PMC 9347599 · doi ↗ · pubmed ↗
- 6G. Verma , M. Dhawan , A. A. Saied , G. Kaur , R. Kumar , and T. B. Emran , “Immunomodulatory Approaches in Managing Lung Inflammation in COVID‐19: A Double‐Edge Sword,” Immunity, Inflammation and Disease 11, no. 9 (2023): e 1020.37773723 10.1002/iid 3.1020 PMC 10521379 · doi ↗ · pubmed ↗
- 7M. Dhawan , A. A. Rabaan , S. Alwarthan , et al., “Regulatory T Cells (Tregs) and COVID‐19: Unveiling the Mechanisms, and Therapeutic Potentialities With a Special Focus on Long COVID,” Vaccines 11, no. 3 (2023): 699.36992283 10.3390/vaccines 11030699 PMC 10059134 · doi ↗ · pubmed ↗
- 8J. Pincemail , E. Cavalier , C. Charlier , et al., “Oxidative Stress Status in COVID‐19 Patients Hospitalized in Intensive Care Unit for Severe Pneumonia. A Pilot Study,” Antioxidants 10, no. 2 (2021): 257–S 120.33562403 10.3390/antiox 10020257 PMC 7914603 · doi ↗ · pubmed ↗