hbl-1 does not contribute to the misexpression of an adult cell fate marker in daf-16 mutant dauer larvae
Matthew J Wirick, Xantha Karp

TL;DR
This study shows that the gene hbl-1 does not help control a specific cell marker in worm larvae lacking daf-16.
Contribution
The paper clarifies that hbl-1 is not a downstream regulator of daf-16 in dauer larvae.
Findings
A gain-of-function hbl-1 allele does not suppress col-19p::gfp expression in daf-16 mutant dauer larvae.
HBL-1 protein is not detectable in control or daf-16 mutant dauer larvae.
HBL-1 does not function downstream of DAF-16 to regulate col-19p::gfp expression.
Abstract
In Caenorhabditis elegans dauer larvae, the FOXO ortholog, daf-16 , opposes the expression of the col-19p::gfp adult cell fate marker in the lateral hypodermis. daf-16 acts in part via lin-41 , a heterochronic gene that promotes larval seam cell fate during non-dauer development. Here, we show that a different heterochronic gene, hbl-1 , does not function downstream of daf-16 to regulate col-19p::gfp expression during dauer. A gain-of-function hbl-1 allele did not suppress ectopic col-19p::gfp expression in daf-16 (0) dauer larvae, and HBL-1 protein was not detectable in control or daf-16 (0) dauer larvae.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
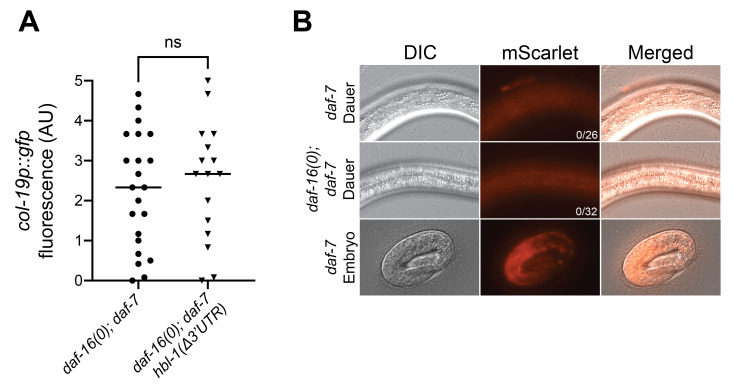 Figure 1
Figure 1|
Strain name |
Genotype |
|
|
|
|
|
|
|
|
|
|
|
|
- —U.S. National Science Foundation (United States)https://ror.org/021nxhr62
- —National Institute of General Medical Sciences (United States)https://ror.org/04q48ey07
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCell Image Analysis Techniques
Description
In * Caenorhabditis elegans , * lateral hypodermal seam cells are multipotent progenitor cells that undergo a particular pattern and sequence of cell divisions at each larval stage until differentiating at adulthood (Sulston and Horvitz 1977). Adult seam cell fate includes the expression of adult-enriched collagens (Ambros and Horvitz 1984; Liu et al. 1995). In dauer larvae, seam cells remain quiescent and multipotent, despite a potentially lengthy period of developmental arrest (Liu and Ambros 1991). We have previously reported that the * C. elegans * ortholog of the Forkhead Box O (FOXO) transcription factor, DAF-16 , blocks adult cell fate during dauer (Wirick et al. 2021). Loss of * daf-16 * ( * “ daf-16 (0)” * ) results in ectopic expression of the col-19p::gfp adult cell fate marker in dauer larvae. The regulation of *col-19p::gfp * by * daf-16 * is at least partially mediated by positive regulation of * lin-41 * , a heterochronic gene required for the temporal patterning of seam cell divisions in larvae during non-dauer development (Slack et al. 2000; Wirick et al. 2021). Though * lin-41 * is required to block ectopic expression of *col-19p::gfp * during dauer, downregulation of * lin-41 * via RNAi incompletely phenocopied the expression of col-19p::gfp in * daf-16 (RNAi) * dauer larvae. Additionally, a gain-of-function * lin-41 * allele only partially suppressed the expression of col-19p::gfp in * daf-16 (RNAi) * dauer larvae (Wirick et al. 2021). Therefore, we hypothesized that * daf-16 * may oppose *col-19p::gfp * expression through additional * lin-41 * -independent mechanisms.
One candidate regulator of *col-19p::gfp * downstream of * daf-16 * is the heterochronic gene * hbl-1 , * which encodes the ortholog of the hunchback transcription factor (Fay et al. 1999; Abrahante et al. 2003; Lin et al. 2003). * hbl-1 * and * lin-41 * are both expressed early in larval development and promote stage-specific seam cell divisions. Later in development , * hbl-1 * and * lin-41 * are downregulated by * let-7
family microRNAs via their 3'UTRs (Reinhart et al. 2000; Abrahante et al. 2003; Lin et al. 2003; Abbott et al. 2005). * hbl-1 , * like * lin-41 , * can modulate the decision to enter dauer (Karp and Ambros 2011; Cale and Karp 2020). In addition, * hbl-1 * has been implicated in the regulation of seam cell fate during the dauer life history as misexpression of * hbl-1 * during or after dauer appears to cause heterochronic phenotypes (Karp and Ambros 2012).
To determine whether * hbl-1 * plays a role in the expression of *col-19p::gfp * observed in * daf-16 (0) * dauer larvae, we first asked if a gain-of-function allele of * hbl-1 * could suppress the *col-19p::gfp * expression phenotype. The gain-of-function allele of * lin-41 * that suppresses this phenotype is a 3'UTR deletion that abrogates the microRNA-mediated downregulation of * lin-41 * (Ecsedi et al. 2015; Wirick et al. 2021). We created a strain containing the * daf-16 ( mgDf50 ) * null mutation and a large deletion in the * hbl-1 * 3'UTR, * hbl-1 ( ma354 ) * , that removes all predicted * let-7 * complementary sites (Ilbay and Ambros 2019). We also included the * daf-7 ( e1372 ) * allele to obtain populations of dauer larvae in dauer-defective * daf-16 (0) * mutants, as this mutation induces dauer formation in * daf-16 (0) * mutants at 24-25°C (Vowels and Thomas 1992; Wirick et al. 2021). We found * hbl-1 ( ma354 [∆3'UTR]) * had no effect on the expression of *col-19p::gfp * in * daf-16 (0) * dauer larvae ( Fig. 1A ).
We next asked whether HBL-1 protein expression was altered in * daf-16 (0) * dauer larvae. Although our prior mRNA-seq data did not show a difference in * hbl-1 * mRNA levels in * daf-16 (0); daf-7 * vs. * daf-7 * larvae (Wirick et al. 2021), levels or localization of HBL-1 protein could be altered in * daf-16 (0); daf-7 * dauer larvae. Indeed, subcellular localization of HBL-1 is a known mode of regulation during continuous development (Ilbay and Ambros 2019). Furthermore, modENCODE data displayed on WormBase (WS296) show a DAF-16 binding peak upstream of * hbl-1 * , suggesting that * daf-16 * may regulate * hbl-1 * expression in some contexts (Gerstein et al. 2010). However, the expression of an endogenously tagged * hbl-1 * allele, HBL-1 ::mScarlet (Ilbay and Ambros 2019), was not detectable in either the * daf-7 * or * daf-16 (0); daf-7 * dauer larvae at exposure settings where HBL-1 ::mScarlet was detectable in embryos ( Fig. 1B ).
Taken together, we conclude that * daf-16 * likely opposes the expression of *col-19p::gfp * during dauer independently of * hbl-1 * . It will be interesting to explore other genes that may act in parallel to * lin-41 * in future work.
Methods
Strains and maintenance
C. elegans * strains were grown and maintained at 15˚C or 20°C on Nematode growth media (NGM) plates seeded with E. coli strain OP50 as a food source (Brenner 1974).
Dauer induction
Dauer was induced at 24˚C using the * daf-7 ( e1372 ) * allele which was present in all strains. For strains harboring the * hbl-1 (mScarlet) * endogenous tag, gravid adults were placed onto NGM plates seeded with OP50 and allowed to lay eggs for 3 hours at 24°C. Gravid adults were then removed from the plates. For * hbl-1 ( ma354 ) * mutants, gravid adults were dissected to release fertilized eggs onto NGM plates seeded with OP50 . Populations of eggs on seeded NGM plates were then incubated at 24°C for 48-50 hours until dauer formation.
Fluorescence microscopy
Dauer larvae were mounted onto glass slides made with 2% agarose pads and paralyzed with 0.1M levamisole. A Zeiss AxioImager D2 compound light microscope with HXP 200 C fluorescent optics was used to image larvae. DIC and fluorescence images were captured with a AxioCam mRm Rev 3 camera and ZEN 3.2 software using a 63x objective. GFP images were captured with a Zeiss filter set 38HE and mScarlet images were captured with a Zeiss filter set 43 HE. The intensity of fluorescence can be altered on the HXP 200 C using a dial with 4 possible levels, where higher numbers indicate greater intensity. The expression of HBL-1 ::mScarlet in the representative embryo image was captured on the 2 ^nd^ intensity level of the HXP 200C exposed for 2000ms, while dauer larvae were imaged on the 3 ^rd ^ intensity level and exposed for 2000ms.
Phenotypic analysis
The experimenter was blinded to genotype prior to imaging and analysis. col-19p::gfp expression was assessed using a semi-quantitative method (Wirick et al. 2021). Briefly, 3 images of col-19p::gfp were captured for each dauer larva along the lateral hypodermis. Each image was compared to existing representative images given a value 0.5-5 based off of the expression of col-19p::gfp , quantified by ImageJ. The average was taken from the three values for each worm and plotted. GraphPad Prism was used to create graphs and conduct the statistical analyses described in the figure legend. To determine the presence or absence of detectable HBL-1 ::mScarlet expression, three images were captured along the lateral hypodermis of dauer larvae and assessed for expression of HBL-1 ::mScarlet.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abbott AL Alvarez-Saavedra E Miska EA Lau NC Bartel DP Horvitz HR Ambros V 200591 The let-7 Micro RNA family members mir-48, mir-84, and mir-241 function together to regulate developmental timing in Caenorhabditis elegans.Dev Cell 931534-580740341410.1016/j.devcel.2005.07.00916139228 PMC 3969732 · doi ↗ · pubmed ↗
- 2Abrahante JE Daul AL Li M Volk ML Tennessen JM Miller EA Rougvie AE 200351 The Caenorhabditis elegans hunchback-like gene lin-57/hbl-1 controls developmental time and is regulated by micro RN As.Dev Cell 451534-580762563710.1016/s 1534-5807(03)00127-812737799 · doi ↗ · pubmed ↗
- 3Ambros V Horvitz HR 19841026 Heterochronic mutants of the nematode Caenorhabditis elegans.Science 22646730036-807540941610.1126/science.64948916494891 · doi ↗ · pubmed ↗
- 4Brenner S 197451 The genetics of Caenorhabditis elegans.Genetics 7710016-6731719410.1093/genetics/77.1.714366476 PMC 1213120 · doi ↗ · pubmed ↗
- 5Cale AR Karp X 20201112 lin-41 controls dauer formation and morphology via lin-29 in C. elegans.Micro Publ Biol 202010.17912/micropub.biology.00032333313484 PMC 7721599 · doi ↗ · pubmed ↗
- 6Ecsedi M Rausch M Großhans H 201529 The let-7 micro RNA directs vulval development through a single target.Dev Cell 3231534-580733534410.1016/j.devcel.2014.12.01825669883 · doi ↗ · pubmed ↗
- 7Fay DS Stanley HM Han M Wood WB 1999115 A Caenorhabditis elegans homologue of hunchback is required for late stages of development but not early embryonic patterning.Dev Biol 20520012-160624025310.1006/dbio.1998.90969917360 · doi ↗ · pubmed ↗
- 8Gerstein MB Lu ZJ Van Nostrand EL Cheng C Arshinoff BI Liu T Yip KY Robilotto R Rechtsteiner A Ikegami K Alves P Chateigner A Perry M Morris M Auerbach RK Feng X Leng J Vielle A Niu W Rhrissorrakrai K Agarwal A Alexander RP Barber G Brdlik CM Brennan J Brouillet JJ Carr A Cheung MS Clawson H Contrino S Dannenberg LO Dernburg AF Desai A Dick L DoséAC Du J Egelhofer T Ercan S Euskirchen G Ewing B Feingold EA Gassmann R Good PJ Green P Gullier F Gutwein M Guyer MS Habegger L Han T Henikoff JG Henz SR Hinrichs A Holster H Hyman T Iniguez AL Janette J Je · doi ↗ · pubmed ↗