Isolation and characterization of LEAFY-homologous genes from two Tricyrtis spp. showing different inflorescence architecture
Sota Takanashi, Yuto Imamura, Haruki Ouchi, Shoichi Sato, Masahiro Otani, Masaru Nakano

TL;DR
This study explores how different inflorescence structures in two Tricyrtis species may be linked to the expression patterns of LEAFY-homologous genes.
Contribution
The study identifies and compares LFY-homologous genes in two Tricyrtis species with distinct inflorescence architectures.
Findings
ThirLFY and TforLFY share similar amino acid sequences and functional motifs.
Transgenic Arabidopsis plants overexpressing either gene showed early flowering and secondary inflorescences.
Expression of ThirLFY occurred in both apical and lateral buds, while TforLFY was only in apical buds.
Abstract
For ornamental plants, inflorescence architecture is one of the most important traits to determine their commercial values. However, molecular mechanisms of inflorescence architecture determination have not yet been fully elucidated. LEAFY (LFY), which encodes a plant-specific transcriptional factor, has been shown to play a key role in the switch from vegetative to reproductive phases. Recent studies have demonstrated that LFY is involved not only in floral meristem induction but also in inflorescence architecture determination. Tricyrtis spp., which belong to the family Liliaceae, show two different types of inflorescence architecture: T. hirta produces both apical and lateral flowers, whereas T. formosana produces only apical flowers. In the present study, we isolated LFY-homologous genes from T. hirta and T. formosana (designated as ThirLFY and TforLFY, respectively) and analyze…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
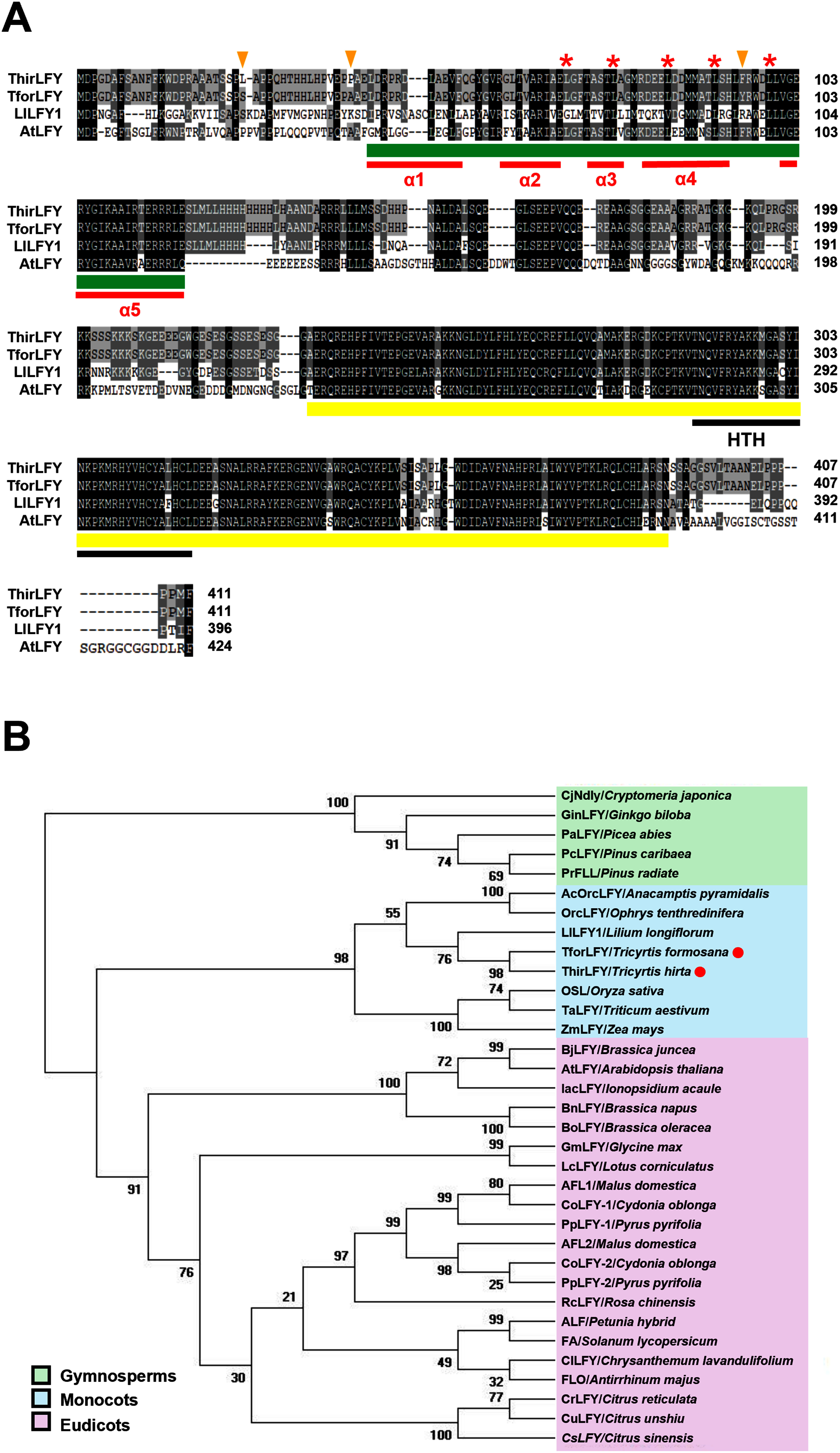 Figure 1
Figure 1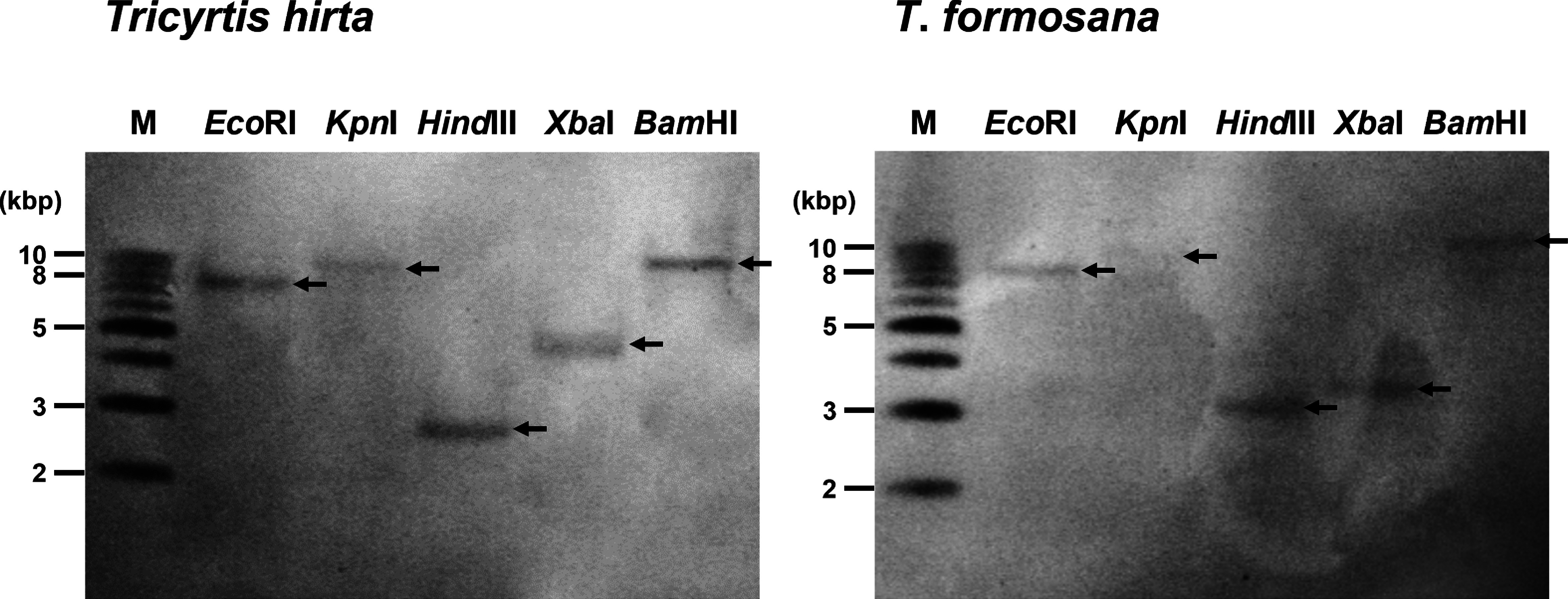 Figure 2
Figure 2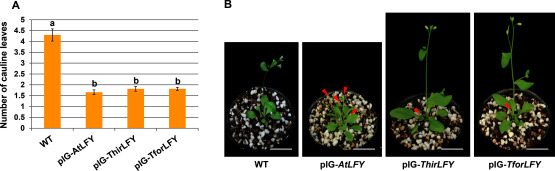 Figure 3
Figure 3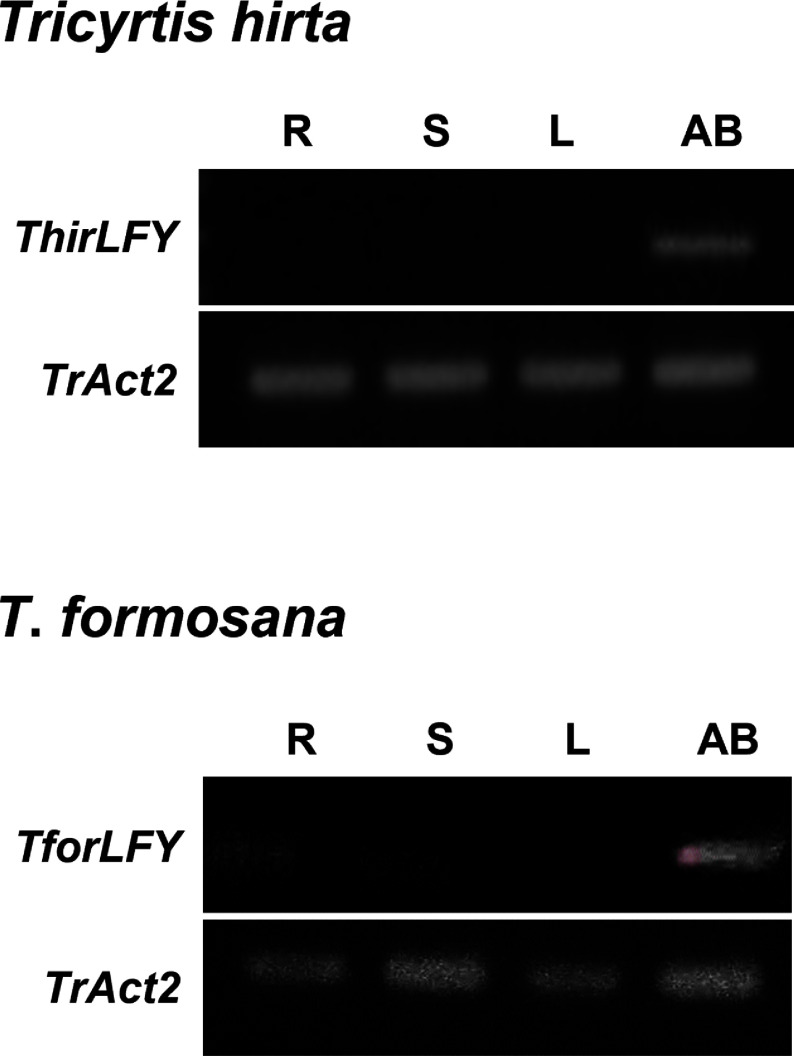 Figure 4
Figure 4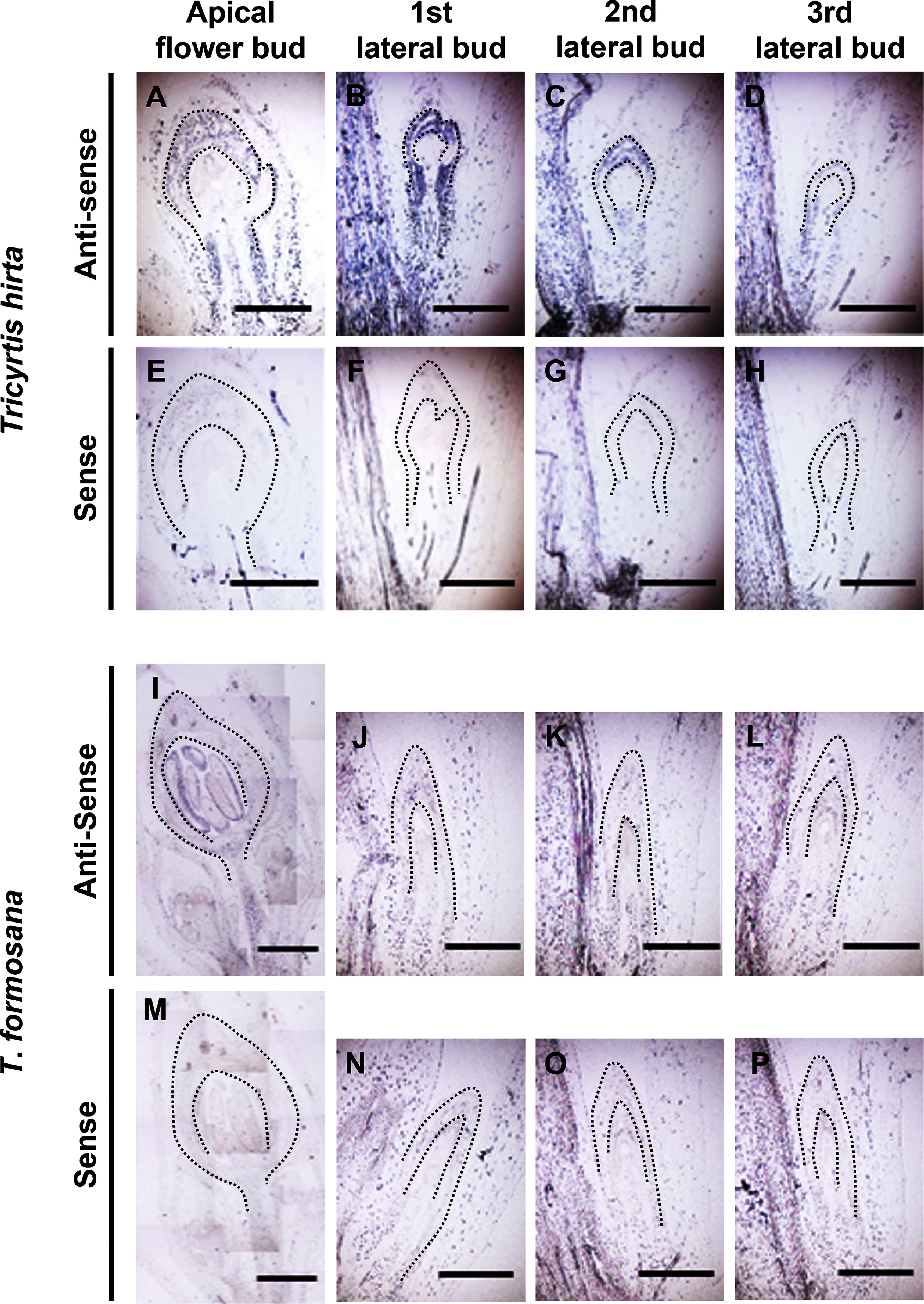 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhotosynthetic Processes and Mechanisms · Plant Reproductive Biology · Plant Gene Expression Analysis
Introduction
LFY, which encodes a plant-specific transcriptional factor, has been shown to play a key role in the switch from vegetative to reproductive phases (Blazquez et al. 1997; Liljegren et al. 1999; Weigel et al. 1992). In Arabidopsis thaliana, lfy mutants exhibited late flowering and produced leaf-like abnormal tepals (Shannon and Meeks-Wagner 1993; Weigel et al. 1992). On the other hand, overexpression of LFY induced early flowering and production of terminal flowers (Weigel and Nilsson 1995).
Several studies have reported that LFY is also involved in determining inflorescence architecture. In A. thaliana, lfy mutants showed abnormal branching patterns through inflorescence-to-vegetative shoot conversion (Schultz and Haughn 1991; Weigel et al. 1992). Similar alterations in inflorescence architecture have also been observed in lfy mutants of Antirrhinum majus, Solanum lycopersicum and Petunia hybrida (Immink et al. 1999; Masiero et al. 2004; Molinero-Rosales et al. 1999). In addition, expression patterns of LFY-homologous genes in meristems have been reported to be different among four brassicaceous species showing different inflorescence architectures (Shu et al. 2000; Sliwinski et al. 2006, 2007; Yoon and Baum 2004). However, no direct evidence for the relationship between LFY-homologous genes and inflorescence architecture has yet been obtained because no transformation system has been established in these brassicaceous species except for A. thaliana.
Tricyrtis spp., which belong to the family Liliaceae, show two different types of inflorescence architecture: T. hirta produces both apical and lateral flowers, whereas T. formosana produces only apical flowers. In addition, an efficient Agrobacterium-mediated transformation system have already been established for Tricyrtis spp. (Adachi et al. 2005). Therefore, T. hirta and T. formosana are expected to be useful as model plants for molecular studies on inflorescence architecture. In the present study, we isolated and characterized LFY-homologous genes from these two species as a first step toward elucidation of molecular mechanisms of inflorescence architecture determination in Tricyrtis spp.
Materials and methods
Plant materials
Potted plants of T. hirta and T. formosana ‘Seiryu’ were cultivated in a greenhouse without heating. A. thaliana plants were cultivated in an incubator under a long-day conditions (16 h light/8 h dark) at 22°C.
Isolation of LFY-homologous genes
Total RNAs were extracted from immature flower buds (ca. 5 mm in length) of potted Tricyrtis plants using RNeasy Plant Mini Kit (QIAGEN, Hilden, Germany) and treated with RNase-Free DNase (QIAGEN) according to the manufacturer’s instructions. In order to amplify full-length cDNA fragments of LFY-homologous genes, RACE was performed using 3′/5′-Full RACE Core Set (Takara, Shiga, Japan) according to the manufacturer’s instructions. Primers used for isolation of LFY homologous genes are listed in Supplementary Table S1.
Phylogenetic analysis
Deduced amino acid sequences of LFY-homologous genes were aligned using ClustalW program (Thompson et al. 2002). Molecular phylogenetic tree based on amino acid sequences was constructed using the maximum likelihood method with 1,000 bootstrap replicates in MEGA-X (Felsenstein 1985; Kumar et al. 2018). Accession numbers of LFY-homologous genes used for phylogenetic analysis are listed in Supplementary Table S2.
Southern blot analysis
Total DNAs were extracted from young leaves of potted Tricyrtis plants the using CTAB method (Doyle 1990; Doyle and Doyle 1987). DNAs were then digested with EcoRI, KpnI, HindIII, XbaI or BamHI. Common sequences in 3′-terminal regions among LFY-homologous genes of Tricyrtis spp. (647 bp) were used as a probe. DNA probe was prepared using PCR DIG Probes Synthesis Kit (Roche, Basel, Switzerland). Hybridization was carried out under 42°C for 16 h with gentle shaking. Blocking, washing and detection were performed using DIG Nucleic Acid Detection Kit (Roche) according to the manufacturer’s instruction.
Transformation of A. thaliana
Agrobacterium tumefaciens strains, EHA101/pIG-ThirLFY and EHA101/pIG-TforLFY, were used. T-DNA regions of the binary vectors pIG-ThirLFY and pIG-TforLFY contain LFY-homologous gene of T. hirta and T. formosana respectively, both under the control of the CaMV35S promoter. T-DNA regions of both binary vectors also contain the NPTII gene under the control of the NOS promoter, and the HPT gene under the control of the CaMV35S promoter. A. tumefaciens strain EHA101/pIG-AtLFY (Sankhuan et al. 2022) was also used as a control. Transformation of A. thaliana was performed using the floral dip method (Clough and Bent 1998). Transgenic plants were cultivated as described above.
Expression analysis by RT-PCR
Total RNAs were extracted from roots, stems, leaves and apical flower buds (ca. 5 mm in length) of potted Tricyrtis plants and then treated with RNase-Free DNase Set (QIAGEN) according to the manufacturer’s instructions. For cDNA synthesis, 500 ng of total RNAs were reverse-transcribed using PrimeScript™ RT reagent Kit (Takara) according to the manufacturer’s instructions.
RT-PCR was performed using EmeraldAmp® PCR Master Mix (Takara) on T100™ Thermal Cycler (Bio-Rad, California, USA). Each PCR reaction was carried out under the following conditions: denaturation at 94°C for 2 min; 30 cycles of 10 s at 98°C, 30 s at 58°C and 1 min at 72°C; and 72°C for 4 min. Amplified products were analyzed by electrophoresis on 1.5% agarose gels. The actin gene of Tricyrtis sp. TrAct2 (accession number AB196260 in the GenBank/EMBL/DDBJ databases) was used as an internal standard. Primers used for RT-PCR analysis are listed in Supplementary Table S1.
In situ hybridization
Apical and lateral buds were collected from potted Tricyrtis plants when apical flower buds had grown to ca. 5 mm in length. They were soaked in FAA (10% formalin, 5% acetic acid and 50% ethanol) with vacuum for 30–60 min and fixed for overnight at 4°C. Fixed samples were dehydrated in an increasing gradient series of ethanol and embedded in paraffin wax. Paraffin-embedded samples were cut into 10 µm sections using Microtome 2035 Biocut (Leica Biosystems, Nussloch, Germany).
RNA probe was designed for 647 bp of common sequences in 3′-terminal regions among LFY-homologous genes of Tricyrtis spp. and synthesized using DIG RNA Labelling Kit (Roche) according to the manufacturer’s instructions. Hybridization and detection were carried out following the DIG application manual for in situ hybridization (Roche).
Results
Isolation and phylogenetic analysis of LFY-homologous genes
In order to identify full-length coding sequences of LFY-homologous genes of T. hirta and T. formosana, we performed RACE using cDNAs, which were synthesized from total RNAs extracted from immature flower buds (ca. 5 mm in length). The specific primers used for 3′-RACE were designed based on the nucleotide sequences of highly conserved C-terminal regions among LFY homologous genes from other plant species. LFY-homologous genes of T. hirta and T. formosana were successfully isolated and designated as ThirLFY (accession number AB829895 in the GenBank/EMBL/DDBJ databases) and TforLFY (accession number AB829896 in the GenBank/EMBL/DDBJ databases), respectively. Both ThirLFY and TforLFY encode 411 amino acid residues, and characteristic motifs of LFY-homologous gene, such as short leucine zipper and DNA-binding domain, have been conserved in their amino acid sequences (Figure 1A). There were only three substitutions in full-length amino acid sequences between ThirLFY and TforLFY: Leu/Ser at position 25, Pro/Ala at position 40, and Phe/Tyr at position 95 (Figure 1A). Amino acid sequences of ThirLFY and TforLFY showed 49.3–67.3% homologies to those of LFY-homologous genes of other plant species. Phylogenetic analysis showed that both ThirLFY and TforLFY belong to the monocotyledon clade and clustered closely with LlLFY1 from Lilium longiflorum (Figure 1B).
Figure 1. Alignment and phylogenetic analysis of ThirLFY and TforLFY based on the amino acid sequences. (A) Alignment of deduced amino acid sequences of ThirLFY (Tricyrtis hirta), TforLFY (T. formosana), LlLFY1 (Lilium longiflorum) and AtLFY (Arabidopsis thaliana). Green and yellow lines indicate conserved N- (include Sterile Alpha Motifs) and C-terminal regions (include DNA-binding domains) of LFY proteins, respectively. Red lines and asterisks indicate alpha helix structures and leucine residues, which are essential component for LFY protein conformation and homodimerization, respectively. Black lines indicate helix-turn-helix (HTH) structure, which is important motif for DNA-binding ability. Orange arrowheads indicate the positions of amino acid substitution between ThirLFY and TforLFY. (B) A molecular phylogenetic tree of LFY-homologous genes. Red spots indicate ThirLFY and TforLFY.
In order to investigate the copy numbers of LFY-homologous genes in the genome of T. hirta and T. formosana, Southern blot analysis was performed. Only a single band was detected for all five restriction enzymes (EcoRI, KpnI, HindIII, XbaI and BamHI) in both T. hirta and T. formosana, indicating that both species have a single copy of LFY-homologous gene (Figure 2).
Figure 2. Southern blot analysis of LFY-homologous genes in the genome of Tricyrtis hirta and T. formosana. Genomic DNAs were digested with each of EcoRI, KpnI, HindIII, XbaI and BamHI.
Functional analysis of LFY-homologous genes
In order to investigate the function of ThirLFY and TforLFY, we produced the overexpression constructs pIG-ThirLFY and pIG-TforLFY, and introduced them separately into A. thaliana. Totally 13 and 17 transgenic lines of ThirLFY- and TforLFY-overexpressing plants were obtained, respectively. Transgenic A. thaliana plants overexpressing ThirLFY or TforLFY exhibited early flowering compared with wild-type plants. The degree of early flowering in these transgenic plants was similar to that in transgenic plants overexpressing AtLFY (Figure 3A). Transgenic plants overexpressing ThirLFY or TforLFY produced secondary inflorescences from rosette leaf axils as in the case of those overexpressing AtLFY (Figure 3B).
Figure 3. Transgenic Arabidopsis thaliana plants overexpressing AtLFY, ThirLFY and TforLFY. (A) Flowering time of wild-type and transgenic plants. Values represent the mean±standard error of 20, 8, 13, 17 biological replicates for the wild-type, pIG-AtLFY, pIG-ThirLFY and pIG-TforLFY, respectively. Mean values with different letters indicate significant difference at the 0.05 level with the Tukey–Kramer’s test. (B) Typical phenotypes of 30-day-old wild-type and transgenic plants. Red arrowheads indicate secondary inflorescence from axils of rosette leaves. Scale bars=2 cm.
Expression analysis of LFY-homologous genes
In order to elucidate the expression patterns of ThirLFY and TforLFY, RT-PCR and in situ hybridization analyses were performed. RT-PCR analysis revealed that both ThirLFY and TforLFY were detected in apical flower buds, but not in roots, stems and leaves (Figure 4). When apical flower buds of potted plants had grown to ca. 5 mm in length, in situ hybridization analysis was performed using apical and lateral buds (Figure 5). ThirLFY transcripts were detected in both apical and lateral buds in T. hirta. On the other hand, TforLFY transcripts were detected only in apical buds but not in any lateral buds of T. formosana. No signals were detected with the sense probe as negative control in all samples.
Figure 4. Semi-quantitative expression of ThirLFY and TforLFY in various tissues of Tricyrtis hirta and T. formosana. R, roots; S, stems; L, leaves; AB, apical flower buds. TrAct2 was used as an internal standard.
Figure 5. In situ hybridization of ThirLFY and TforLFY in apical flower buds and lateral buds of Tricyrtis hirta (A–H) and T. formosana (I–P). (A, E, I, M) Apical flower buds. (B, F, J, N) First lateral buds below the apical flower buds. (C, G, K, O) Second lateral buds below the apical flower buds. (D, H, L, P) Third lateral buds below the apical flower buds. Scale bars=1 mm.
Discussion
In the present study, we have isolated LFY-homologous genes from two Tricyrtis spp., T. hirta and T. formosana, showing different types of inflorescence architecture. Deduced amino acid sequences of ThirLFY and TforLFY showed high homologies to those of LFY-homologous genes of other plant species and shared well-conserved N- and C-terminal regions (Figure 1A, B). Furthermore, overexpression of ThirLFY or TforLFY induced early flowering and development of secondary inflorescences (Figure 3), which are typical phenotypes of transgenic A. thaliana plants overexpressing LFY (Weigel and Nilsson 1995). These results indicate that ThirLFY and TforLFY function as LFY in T. hirta and T. formosana, respectively.
In the genus Cornus (Cornaceae), some amino acid substitutions were found in C-terminal region containing an essential domain for DNA-binding specificity among deduced amino acid sequences of LFY-homologous genes of six species showing different types of inflorescence architecture, suggesting that inflorescence architecture could be determined by different function of LFY-homologous genes (Liu J et al. 2013). On the other hand, in the present study, only three amino acid substitutions were found in N-terminal regions between ThirLFY and TforLFY: Leu/Ser at position 25, Pro/Ala at position 40, and Phe/Tyr at position 95 (Figure 1A, Supplementary Figure S1). In particular, at position 95, ThirLFY conserves hydrophobic residues as in the case of most LFYs from other plant species, whereas TforLFY conserves hydrophilic residues (Supplementary Figure S1). In LFY, N-terminal regions include some alpha helix structures and short leucine zippers called SAMs, which play an essential role in precise conformation and homodimerization (Sayou et al. 2016; Siriwardana and Lamb 2012). Therefore, it is possible that ThirLFY and TforLFY have different functions in floral development and inflorescence architecture determination in Tricyrtis spp. However, no apparent morphological difference was observed between transgenic A. thaliana plants overexpressing ThirLFY and those overexpressing TforLFY (Figure 3), suggesting that ThirLFY and TforLFY may have no functional difference.
Different expression patterns of LFY-homologous genes have been reported to resulted in different inflorescence architectures in many plant species (Blazquez et al. 1997; Kyozuka et al. 1998; Souer et al. 1998). In the present study, ThirLFY expression was detected in both apical and lateral buds in T. hirta which produces both apical and lateral flowers, whereas TforLFY expression was detected only in apical buds in T. formosana which produces only apical flowers (Figures 4, 5). Thus, different expression patterns of LFY-homologous genes are consistent with different flower production patterns among T. hirta and T. formosana, and two different types of inflorescence architecture in Tricyrtis spp. are likely to be caused by different expression patterns of LFY-homologous genes. Promoter analysis for ThirLFY and TforLFY using GUS reporter gene are now in progress.
In A. thaliana, LFY expression is directly or indirectly regulated by several transcriptional factors, such as FT and TFL1. FT promotes floral transition through activation of floral meristem identity genes such as LFY, while TFL1 represses expression of such genes to maintain vegetative state of meristem tissues (Kardailsky et al. 1999; Ratcliffe et al. 1999; Zhu et al. 2020). In addition, TFL1 have also been reported to be involved in inflorescence architecture determination. In A. thaliana, tfl1 mutants have terminal flowers and secondary inflorescences from rosette leaf axils, and such inflorescence architecture is apparently different from wild-type plants (Liljegren et al. 1999; Shannon and Meeks-Wagner 1991). Similar changes in inflorescence architecture have also been observed in tfl1 mutants of other plant species, such as Antirrhinum majus, Oryza sativa and Solanum lycopersicum (Cremer et al. 2001; Liu C et al. 2013; Pnueli et al. 1998). It is quite possible that, in addition to LFY-homologous genes, FT- and TFL1-homologous genes are also involved in inflorescence architecture determination in Tricyrtis spp. Thus, we are now examining isolation and characterization of FT- and TFL1-homologous genes in both T. hirta and T. formosana.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adachi Y, Mori S, Nakano M (2005) Agrobacterium-mediated production of transgenic plants in Tricyrtis hirta (Liliaceae). Acta Hortic 673: 415–419
- 2Blázquez MA, Soowal LN, Lee I, Weigel D (1997) LEAFY expression and flower initiation in Arabidopsis. Development 124: 3835–38449367439 10.1242/dev.124.19.3835 · doi ↗ · pubmed ↗
- 3Clough SJ, Bent AF (1998) Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 16: 735–74310069079 10.1046/j.1365-313x.1998.00343.x · doi ↗ · pubmed ↗
- 4Cremer F, Lönnig WE, Saedler H, Huijser P (2001) The delayed terminal flower phenotype is caused by a conditional mutation in the CENTRORADIALIS gene of snapdragon. Plant Physiol 126: 1031–104111457954 10.1104/pp.126.3.1031 PMC 116460 · doi ↗ · pubmed ↗
- 5Doyle JJ (1990) Isolation of plant DNA from fresh tissue. Focus 12: 13–15
- 6Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull 19: 11–15
- 7Felsenstein J (1985) Confidence limits on phylogenies: An approach using the bootstrap. Evolution 39: 783–79128561359 10.1111/j.1558-5646.1985.tb 00420.x · doi ↗ · pubmed ↗
- 8Immink RGH, Hannapel DJ, Ferrario S, Busscher M, Franken J, Lookeren Campagne MM, Angenent GC (1999) A petunia MADS box gene involved in the transition from vegetative to reproductive development. Development 126: 5117–512610529428 10.1242/dev.126.22.5117 · doi ↗ · pubmed ↗