Genetic analysis of silique and seed traits in Brassica juncea (L.) Czern. under differential doses of nitrogen application
Javed Akhatar, Anna Goyal, Meenakshi Mittal, Heena Verma, Kaur Gurpreet, Beerpal Kaur, Surinder S. Banga, Chhaya Atri

TL;DR
This study explores how nitrogen levels affect silique and seed traits in mustard, identifying genes that could help improve breeding through marker-assisted selection.
Contribution
The study identifies 16 key genes associated with silique and seed traits in mustard under varying nitrogen levels.
Findings
Genotype × nitrogen interactions significantly affect silique length, seeds per silique, and rupture energy.
RNA-seq data revealed expression-level variations for candidate genes like SHP2 and BG4.
The study provides insights for marker-assisted selection in mustard breeding.
Abstract
Mustard, a major source of edible and industrial grade oils in the Indian subcontinent and various regions of Australia, Eastern Europe, and Canada, is also a protein resource for the animal feed industry. Silique and seed size are key traits for mustard improvement, but their inheritance mechanisms are not fully understood. We evaluated 92 inbred lines for silique length, seeds per silique, seed size, and rupture energy required to shatter a pod at three levels of nitrogen application in two crop seasons. Genotypes showed large phenotypic variations and a continuous distribution for all silique-related traits, suggesting a quantitative inheritance. Genotype × nitrogen interactions were significant for silique length, seeds per silique, and rupture energy. Association analysis identified 59 significant SNPs, whose annotations facilitated the prediction of 16 important genes underlying…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
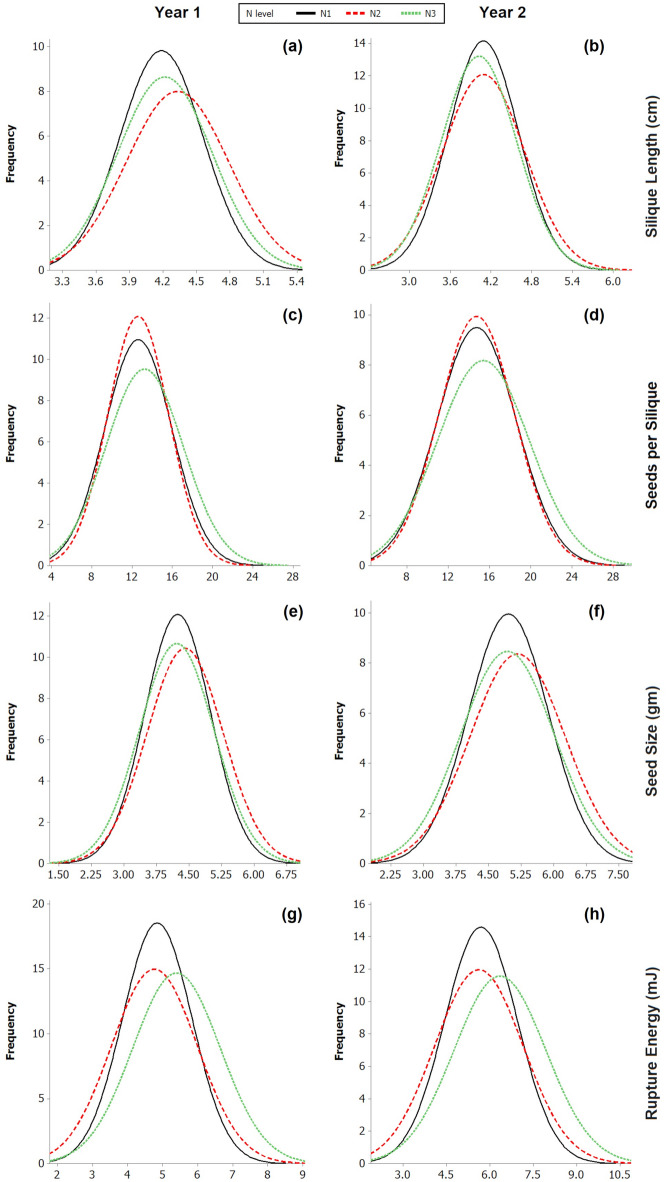 Figure 1
Figure 1 Figure 2
Figure 2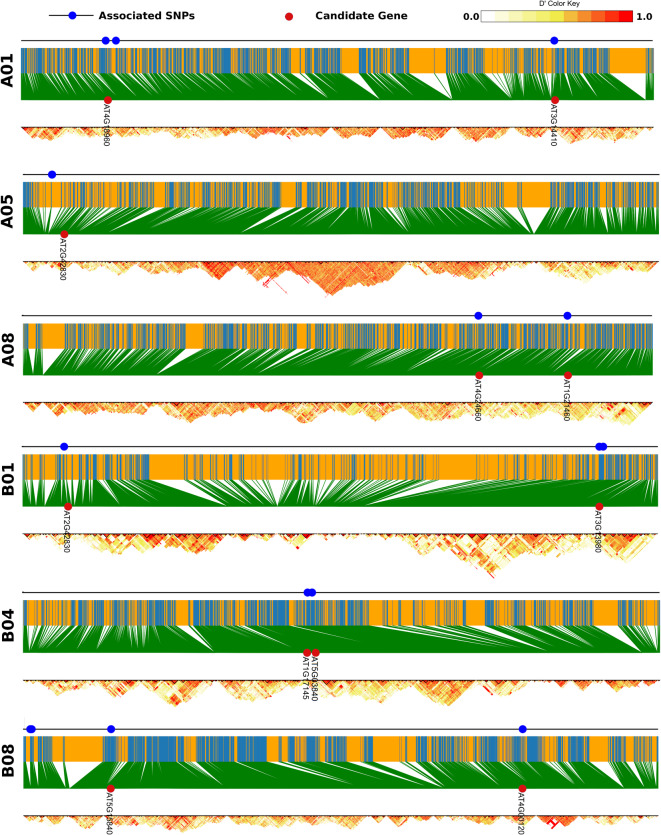 Figure 3
Figure 3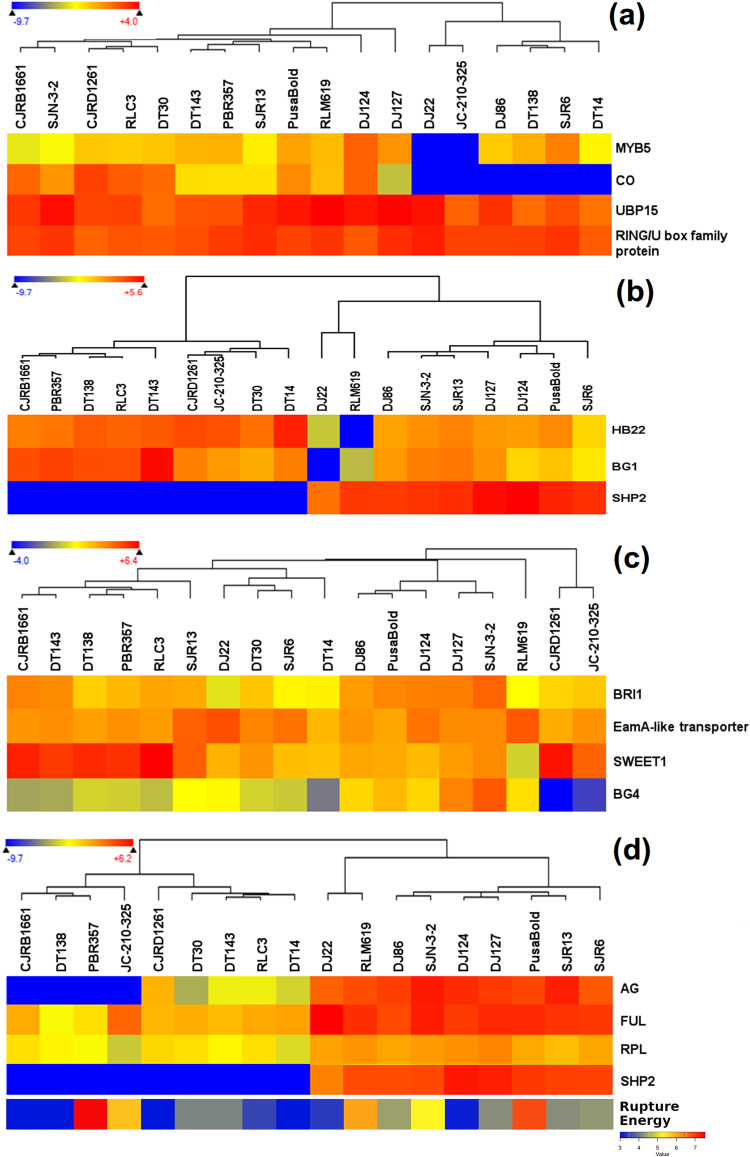 Figure 4
Figure 4- —http://dx.doi.org/10.13039/501100001407Department of Biotechnology, Ministry of Science and Technology, India
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNitrogen and Sulfur Effects on Brassica · Plant nutrient uptake and metabolism · Plant Disease Resistance and Genetics
Introduction
Indian mustard (Brassica juncea (L.) Czern and Coss.) belongs to the rapeseed-mustard group of crops, which ranks among the most traded agricultural commodities in the world. Mustard is widely cultivated in India, China, Eastern Europe, and Russia, with India being a major producer. India accounts for 19.8 percent of the area and 9.8 percent production of rapeseed-mustard in the world (http://www.drmr.res.in/about_rmcrop.php). Although mustard is primarily grown for edible oil and oilcakes, there is growing interest in its cultivation as a biofuel and industrial crop^1–3^.
Crop yield is the main breeding objective in mustard, but this poses challenges due to the interplay of numerous genetic and environmental factors^1,4^. Research highlights that silique per plant, seeds per silique, and seed size have the most significant direct effects on crop yield^4–6^. In Brassica species, an evolutionary trade-off exists between seed mass (size), seed number, and silique development^7–10^. However, focusing on individual yield components—such as silique length, rapid seed development, and bold seed size—can lead to faster selection gains, as these traits are more stably inherited and less affected by environmental changes^11–14^. Silique length and rapid seed development are key indicators of yield potential^15–18^, while bold seed size is valued for seedling establishment^19–21^. Studies in crop Brassica show that over 70% of the total photosynthetic contribution to seed growth and oil accumulation in crop Brassica comes from silique walls^22–27^. Experiments involving defoliation and shading in B. napus and B. juncea reveal that leaf photosynthesis greatly influences silique number. Additionally, silique walls serve as vehicles for seed dispersal under natural conditions through a finely regulated silique-shattering mechanism. However, uncontrolled silique shattering can adversely affect yields in managed agriculture, making resistance to premature pod shattering a vital trait for securing productivity. Resistance to premature silique shattering was possibly the earliest and most important trait selected during crop domestication^8,28,29^. A reduction in seed shattering was favoured over complete non-shattering for the ease of threshing^30,31^. Genome-Wide Association Studies (GWAS) have allowed leveraging of high-density SNP markers to identify genetic variations linked to key traits like yield, oil content, disease resistance, and stress tolerance etc^32^. Such studies have also helped to identify genetic regions affecting pod length and seed size and pod shattering in Brassica species. Genes like BEE1, PEROXIDASE, TCP8, ADPG1, SHP1, and MYB116 linked to pod shatter resistance^8–33^. Regional association mapping, focussed on specific genomic regions identified using QTL mapping, has been frequently to conduct higher resolution of causal genes in rice, wheat, and mustard^34^. Studies in Arabidopsis have shown that seed size depends on the proliferation and expansion of cells during organ development^35,36^. TRANSPARENT GWAS has been instrumental in identifying numerous loci associated with silique-related traits such as silique length, seed number per silique, and thousand seed weight. For instance, a study identified nine loci associated with silique elongation length (SEL), with significant SNPs located on the A02 chromosome, which are crucial for enhancing seed yield by influencing seed number per silique and thousand seed weight TESTA GLABRA2 (TTG2), AUXIN RESPONSE FACTOR2 (ARF2) and KLUH/CYTOCHROME P450-78A5 (CYP78A5) are major genes which contribute to seed size development in Arabidopsis^36^. These mainly promote proliferation and elongation integument^37–40^. In contrast, APETALA2 (AP2) reduces the seed size by suppressing cell elongation in the integument^41^. Transcriptional profiling of the silique walls at different developmental stages has underlined the contribution of transcription factors^42^. Cytokinins act downstream of IKU pathway to seed development^43^. These also promote the integration of epigenetic and genetic forces for endosperm development^44^ and seed size^45^. GWAS has been instrumental in identifying numerous loci associated with silique-related traits such as silique length, seed number per silique, and thousand seed weight. INDEHISCENT (IND), ALCATRAZ (ALC), SHATTERPROOF1 (SHP1), SHATTERPROOF2 (SHP2), and FRUITFULL (FUL) are important regulators of silique dehiscence in Brassicaceae members^46–49^. The TCP3 gene, regulated by miR319, also plays a role in modulating silique development and shattering resistance by influencing the expression of downstream genes like FUL^50^.
Indian forms of B. juncea are resistant to premature silique shattering^51^. However, yield losses caused by uncontrolled silique shattering have been observed in newly developed F_1_ hybrids and ‘00’ genotypes. Environmental conditions, particularly aridity, have been shown to exacerbate silique shattering. This trait is also important for aridity resilience and the prevention of yield losses due to increased fragility of siliques caused by their repeated wetting during unseasonal rains. Studies on grain legumes and cowpea indicate that arid conditions increase the frequency of shattering-resistant alleles, suggesting a natural selection pressure for resilience in such environments^52,53^. Nitrogen application and planting density, also influence shattering resistance. Optimal nitrogen levels and planting densities have been reported to enhance silique characteristics, including lignin content and structural integrity, thereby improving resistance to shattering in B. napus^54^. Research on mustard is limited, particularly regarding the genetic basis of silique traits in response to deficiency or excess of applied nitrogen. The present investigations were conducted to unravel the genetics underlying the phenotypic variation for silique length, seeds per silique, seed size, and silique shattering in Indian mustard under differing levels of N application. Understanding their inheritance is critical to developing an efficient breeding program, as these traits directly determine crop productivity. We used an association panel comprising 92 SNP genotyped inbred lines of Indian mustard. The average distance of LD decay and low population stratification^55^ make this panel eminently suitable for enhanced mapping resolution during genome-wide association studies.
Results
Phenotypic Variation and correlation analysis for traits-related to silique
The target silique attributes differed significantly at three different doses of N fertilization. The distribution of the variation was near normal (Table 1, Fig. 1). Analysis of variance (ANOVA) suggested the significance of variations due to genotypes, years, and N levels for silique length (SL), seed size (SS) and rupture energy (RE) (Table 2). SPS, SS, and RE had significant genotype x N-level interactions (GNI). However, genotypes × year interactions (GYI) were significant only for the SL. N-level × Year and Genotype × N-level × Year interactions were non-significant. Descriptive statistics for SL, SPS, SS and RE are available in Table 1. SL varied from 3.29 to 5.47 cm over all N levels during Y1. However, the trait values (2.87–6.28 cm) varied more during Y2. SL averaged at 4.14 cm for the inbred lines, with a maximum PL of 6.28 cm recorded at N3. The trait variations were at their maximum at N2 for SL. SPS ranged from 5.00 to 28.00 for Y1 and 5.88 to 28.93 during Y2. SPS averaged at 13.69 over 2 years and a maximum SPS was recorded at N3. RLM-619-AB and ELM-108 (N1), DT-70, and DT-25 (N2) and ELM-134 and ELM-151 (N3) revealed a higher number of SPS at the respective levels of N application. The mean SS was higher at N2 during both years, with a maximum of 5.17 gm during Y2. It ranged from 1.55 to 7.28 g and 2.05 to 7.68 g during Y1 and Y2, respectively. MCN-12-60 showed maximum SS at N1. RE, averaged over all genotypes, was higher in Y2 compared to Y1. The RE values ranged from 2.80 to 8.80 mJ for Y1 and 2.78 to 10.35 mJ for Y2. RE ranged from 3.05 to 9.58 mJ, when averaged over 2 years. MCP-12-606, MCN-12-40, and MCN-12-45 showed maximum RE at N1, N2, and N3 levels, respectively. Overall, SL and SS were higher at N2, whereas SPS and RE showed better performance at N3. The coefficients of variation for SL and SS were lower than those recorded for SPS and RE (Fig. 1). SPS, SS, and RE revealed highly significant correlation values (Fig. 2).Table 1. Basic descriptive statistics of silique traits in B. juncea during 2015–16 and 2016–17.Trait (unit)YearMean ± SERangeCoefficient of variationN1N2N3N1N2N3N1N2N3MinMaxMinMaxMinMaxSilique Length (SL) (cm)Y14.19 ± 0.044.33 ± 0.054.22 ± 0.043.365.153.295.473.445.348.910.610.1Y24.09 ± 0.054.09 ± 0.064.02 ± 0.062.955.202.945.552.876.2812.714.813.8YP4.14 ± 0.034.21 ± 0.044.11 ± 0.033.484.903.435.253.684.887.310.07.4Seeds per Silique (SPS)Y112.61 ± 0.3112.61 ± 0.2813.24 ± 0.355.0024.006.0020.007.0028.0033.829.735.2Y214.77 ± 0.3714.77 ± 0.3415.42 ± 0.415.8826.747.0623.537.2528.9333.731.135.3YP13.69 ± 2.7613.69 ± 2.4814.30 ± 2.775.4425.376.5321.777.1227.0528.725.727.5Seed Size (SS) (g)Y14.24 ± 0.074.41 ± 0.084.22 ± 0.082.676.512.197.081.557.2823.224.025.5Y24.96 ± 0.095.17 ± 0.104.94 ± 0.103.117.472.817.682.057.6123.725.927.4YP4.60 ± 0.674.79 ± 0.684.56 ± 0.752.896.912.557.251.807.4520.820.423.0Rupture Energy (RE) (mJ)Y14.85 ± 0.104.77 ± 0.124.22 ± 0.122.807.452.358.802.818.6728.233.028.6Y25.70 ± 0.135.62 ± 0.156.35 ± 0.153.098.672.7810.353.0910.0730.436.231.5YP5.28 ± 0.985.20 ± 1.055.88 ± 1.113.057.612.579.583.059.3728.328.927.0Fig. 1Frequency distribution curve of pod traits at three N level in 2 years (Year 1 and Year 2); (a,b) silique length, (c,d) seeds per silique, (e,f) seed size and (g,h) rupture energy required to shatter a silique in Brassica juncea association panel. [14.7 cm (H) × 8.4 cm (W)].Table 2. Analysis of variation (ANOVA) with different source of variation including interaction for silique traits.Source of variationDFMean sum of squareSLSPSSSREN level21.19631.2376.36545.665Year17.2181313.756149.007214.642Replication13.108**46.4410.00911.918Genotype910.53973.2575.3659.954Genotype × Year910.6985.8570.6441.027N level × Genotype1820.16248.4182.5095.341N level × Year20.5980.1960.0420.288N level × Genotype × Year1820.2160.9400.0260.065*, ** and *** Significance level @ 0.05, 0.01 and 0.001, respectively.Fig. 2. Pearson’s correlation analysis for four traits; (a) silique length, (b) seeds per silique, (c) seed size and (d) rupture energy. The correlation coefficient between years across N level are shown in upper right diagonal boxes for each traits and lower left diagonal depict the scatter plots of coefficient value. [19.8 cm (H) × 19.0 cm (W)].
Genome-wide association mapping for silique traits
GWAS were conducted for four silique traits using year-wise and pooled transformed trait values. MVP facilitated the identification of significant marker trait associations (MTAs). SNPs identified as significant based on quantile–quantile (QQ) plots (Supplementary Table 1 and Supplementary Fig. S1) or environment were classified as consistent loci. Results are described separately for the traits investigated.
Silique length (SL)
Five significant SNPs were found to be associated with SL, with the PVE ranging from 3.80 to 9.82%, on the chromosome B1, B3, B4 and B8, of which four were identified at N3 alone (Table 3; Supplementary Fig. S2). UBIQUITIN SPECIFIC PROTEASE 15 (UBP15), RING/U-box superfamily protein, and CONSTANS (CO) were predicted on chromosomes B03 (N1Y1), B04 (N3Y1), and B08 (N3Y1) respectively. MYB Domain 5 MYB5 was envisioned on chromosome B01 for the data pooled over years.Table 3. Identified predicted genes and functions by GWAS and RAM analysis at N level and across the year for silique traits.Predicted geneTraitN LevelCHRGWASRAMGene functionN1N2N3SNP position/IDNo. SNPsR^2^Nearest SNP (kb)SNP intervalNo. SNPsNearest SNP (kb)AT3G13540—myb domain protein 5—MYB5SL––YPB0141678091-9523.8036.8041639724-4164176250.24Formation of the seed coat and underlying endosperm layersAT1G17110—UBIQUITIN-SPECIFIC PROTEASE 15—UBP15Y1––B03280801119.8247.792803305-2813913441.89Regulates the seed sizeAT1G17145—RING/U-box superfamily protein––Y1B041945304817.8144.8219407232-1941237380.74Regulate Seed and Organ SizeAT5G15840—CONSTANS—CO**––Y1B08974677219.0649.179704021-970407940.19Regulates flowering timeAT2G42830—SHATTERPROOF2—SHP2SPP, REY2–Y1, Y2, YPA05, B011402422-1412718; 30156151218.11650, 2902063201-206371480.25Formation of the podAT4G24660—HOMEOBOX PROTEIN 22—HB22/ZINC FINGER HOMEODOMAIN 2—HB22SPS––Y2A0818859189-1885919028.3619.8518877624-1888040490.08Floral development, female gametophyte developmentAT5G12050—BIG GRAIN 1—BG1Y1, YP––A1016086496-16086528214.6618.9616086496-16103308418.96Regulate biomass, grain size and yieldAT3G14410—Nucleotide/sugar transporter family proteinSS––Y1, Y2, YPA0136029492-3602976039.38–12.3011.0236025796-3602589557.33Nucleotide/sugar transporter family proteinAT4G39400-BRASSINOSTEROID INSENSITIVE 1—BRI1**––Y2A063089305118.688.1830902485-3090254441.26Enhances cell elongation; promotes pollen development, chilling and freezing tolerance; controls vasculature developmentAT1G21460—Bidirectional sugar transporter SWEET1**–Y1, YP–A0822546205-22546302208.93–9.284.9722550690-2255556140.58Tonoplast transporterAT3G13980—SKI/DACH domain protein/BIG GRAIN 4—BG4*––Y1, Y2, YPB0141400644-41400793512.98–15.1812.7141415171-4141958090.58Regulates grain sizeAT1G12500—Probable sugar phosphate/phosphate translocatorY2, YP––B032121316-212143133.81–5.4912.962132714-2136878160.34Nucleotide-sugar transporterAT1G77610—EamA-like transporter family protein**Y2, YP––B0621574550-2157471828.61–7.5912.6721587193-2158719920.02Amino acid transporter crucial for the amino acid homeostasis of siliqueAT4G18960—AGAMOUS1—AG1REY2Y1, Y2, YPA015791617 648111227.7884.05870187-587830142.22Regulate SHP1and *SHP2AT5G60910—FRUITFULL—FUL**––Y2B0259332298114.5246.059373033-59384081390.06Expression of the valve margin identity genes (SHP1 and SHP2)AT4G00120—INDEHISCENT—IND–Y1, Y2, YP–B085501595018.3924.754995359-5499551024.12Pod valve margin development*SNPs identified within the gene; CHR, Chromosome; GWAS, Genome Wide Association Studies; RAM, Regional Association Mapping; R^2^, Phenotypic variation explained.
Seeds per Silique
We identified 16 significant SNPs associated with this trait (Table 3, Supplementary Fig. S3), explaining 8.36% to 18.11% of the observed trait variation. Among them, 11 SNPs were located on chromosome A05 (1402422 to 1412718) at N3 for Y1 and YP. Annotation of the genomic regions around peak SNPs allowed prediction of SHATTERPROOF2 (SHP2), repeatedly over Y1, Y2, and YP at N1 and N3 for both association with SPS. Additionally, HOMEOBOX PROTEIN 22 (HB22) was identified on chromosome A08 (N3Y2). Furthermore, BIG GRAIN1 (BG1) was located on chromosome A10 (N1, Y1, YP). It was positioned 18.96 kb from the peak SNP A10-16,086,496.
Seed size
A total of 34 SNPs were recorded for the trait, with R^2^ values ranging from 3.81 to 15.18% (Table 3; Supplementary Fig. S4). These included several transporter genes such as: AT1G12500 (Probable sugar phosphate/phosphate translocator) and AT1G77610 (EamA-like transporter family protein) were located on chromosomes B03 and B06, respectively, at N1. AT1G21460 (Bidirectional sugar transporter SWEET1) was associated with a cluster of 20 SNPs (22546205-22546302) on chromosome A08. Nine SNPs were significant at the N3 level, among which three were associated with AT3G14410 (Nucleotide/sugar transporter family protein) on chromosome A01. SNPs linked to this gene were predicted repeatedly during Y1, Y2, and YP at N3. BRASSINOSTEROID INSENSITIVE 1 (BRI1) was also predicted on chromosome A06, positioned 8.18 kb away from the associated NP. BIG GRAIN 4 (BG4) was linked to five SNPs on chromosome B01 (Y1, Y2, YP).
Rupture energy
Four identified genes together explained 48.8 percent of the observed phenotypic variation (Table 3; Supplementary Fig. S5). AT4G18960: AGAMOUS1 (AG1) was annotated for SNPs found on chromosome A01 at N1 (Y2) and N3 (Y1, Y2, YP). AT5G60910—FRUITFULL (FUL) was proposed near the SNP, B02_59332298 SNPs at N3 (Y2). AT4G00120—INDEHISCENT (IND) -was repeatedly envisioned on chromosome B08 at N2. It was expected at 24.7 kb from the peak SNP B08_55015950. SHP2 was predicted on chromosome B01.
Gene prediction on the basis of regional association mapping
Regional analysis revealed a total of 136 SNPs located close to the predicted candidate genes included in the validation GWAS results (Table 3). Many of these flanked the predicted candidate genes or were present within the predicted genes (CO, AT1G77610, SWEET1, BR1, AG1, and FUL). A larger number of associated SNPs were recognised for AT1G17145, TFL1, MYB5, HB22, AT1G12500, and BG4. LD block estimation of predicted candidate genes on selected chromosomes fell in the region of high LD (red diagonal area) depicted in Fig. 3.Fig. 3LD plot shows pairwise LD (D) estimation between SNP calculated within 1 Mb region; Honey coloured bar depict chromosome length and blue line are SNP position connected with green lines to LD region (yellow dots—low LD and red dots—high LD). Blue dots show trait associated SNPs and red dots depict the candidate gene location on the chromosome (colour figure online). [16.0 cm (H) × 12.7 cm (W)].
Expression patterns of predicted candidate genes
RNA-seq data from 18 mustard genotypes was used to establish expression-level variations for 15 candidate genes (Fig. 4, Supplementary Table 2). Clustering based gene expression profiles produced two broad genotypic groups. However, genotypic groupings based on the gene expression profiles of some predicted candidates differed from the grouping based on their phenotypic performance. These included the upregulation of SHP2 for SPS and the upregulation of AG1, FUL, and SHP2 with resistance to silique shattering. Downregulation of BG4 was indicated for two genotypes (CJRD1261 and JC-210-325) with the smallest seeds.Fig. 4. Expression level variations for the identified candidate genes**.** (a) Silique length, (b) seeds per silique, (c) seed size and (d) rupture energy. Almost all genotypes showing upregulation of SHP2 had bold seeds. Similarly, upregulation of AG, FUL and SHP2 was observed in the genotypes with shattering resistant genotypes. Cv. PBR-357 was the only exception to these trends. This genotype is bold seeded and possess shattering resistant silique. [38.5 cm (H) × 25 cm (W)].
Discussion
Crop performance in mustard depends on the number of siliques per plant. They not only house the developing seeds but also influence canopy architecture, light interception, photosynthetic potential, and the source strength of plants. Available studies shows that optimizing silique number and seed size is crucial for improving Brassica productivity. Genetic analyses have identified several QTLs associated with silique traits, particularly in rapeseed-mustard crops. However, many previous studies focused on B. napus, limiting their direct applicability to B. juncea. GWAS approaches have significantly expanded genetic information regarding silique size, seed size, and silique dehiscence. Several candidate genes, including UBP15, CO, SHP1, SHP2, BG1, BG4, BRI, AT1G12500, AT1G77610, SWEET1, and AT3G14410, have been identified for their roles in silique formation, grain development, and nutrient transport. Additionally, FUL, IND, and AG1 play key roles in silique shattering and seed dispersal.
Crop performance in mustard is directly associated with the number of siliques per plant, which influence canopy architecture, light interception, photosynthetic potential, and source strength. Maximum silique area index nearly equals leaf area index at the start of ripening^16^. Silique growth ends when a genetically primed seed size is achieved, which is critical for crop stand establishment under rainfed conditions. Large seeds produce seedlings with higher dry weight, root length, and root to shoot ratio compared to those raised from small seeds^56^. Seed size is also a key to cultivar acceptance by growers and processors, as bold seeds are preferred by the trade in India. Optimizing silique number and seed size is essential for improving Brassica productivity. Genetic analyses have produced conflicting genetic models to explain the inheritance of silique traits. These include identifying quantitative trait loci (QTLs) for silique traits in rapeseed-mustard crops^57–61^ and characterizing co-localizing QTL relationships among them^62,63^. The association panel used for current studies exhibited wide phenotypic variations and continuous distribution for all silique-related traits, underscoring their quantitative inheritance. A significant correlation between test traits is expected for traits sharing a common metabolic pool. Seeds per silique and seed size are negatively correlated, requiring a trade-off between number and size. Changes in growth conditions influence genetic variance within traits, which in turn affects genetic interactions among traits sharing a common developmental pathway.
However, most of the previous studies focused on B. napus, limiting their direct applicability to B. juncea^62^. The association panel used for current studies exhibited wide phenotypic variations and continuous distribution for all silique-related traits, underscoring their quantitative inheritance. We assessed our association panel under varying nitrogen (N) fertilization levels, recognizing that different test environments significantly influence the estimation of genetic parameters. These environmental variations play a crucial role in shaping phenotypic expression, affecting trait heritability and genetic interactions. Studies have shown that nitrogen availability impacts yield-related traits, genetic polygenicity, and selection patterns in many crops, including B. juncea^64^.
Nitrogen (N) is not only a vital plant nutrient but it is also a key signaling molecule, influencing various physiological and developmental processes^65^. A strong correlation among these traits was expected, given their shared metabolic pathways. Seeds per silique and seed size exhibit a negative correlation, reflecting an evolutionary trade-off between number and size. This relationship is commonly observed across plant species, where resource allocation dictates a balance between seed quantity and individual seed robustness. Marginal variations in genetic relatedness were recorded across three levels of N fertilization, highlighting the impact of environmental conditions on genetic variance. These changes further influence genetic interactions among traits that share developmental pathways, reinforcing the complexity of trait inheritance and expression.
GWAS allowed the prediction of several candidate genes related broadly to sugar transport, shoot apical meristem, floral and seed development. Important genes namely, UBP15 and CO were identified for their roles in the inheritance of silique length. CO also functions as a metabolic switch that influences seed size and endosperm cellularization through ABSCISIC ACID INSENSITIVE 5 (ABI5)^66^. CO further promotes flowering at the shoot apex^67^. Seed formation is a crucial reproductive process, and MADS-box genes SHATTERPROOF 1 and 2 (SHP1 and SHP2) specify integument identity in Arabidopsis ovules. Our studies highlighted the importance of genes associated with silique formation (SHP2), grain development (BG1 and BG4), cell elongation (BRI), and grain filling (AT1G12500**,** AT1G77610**,** SWEET1**,** and AT3G14410) in determining silique traits. These genes were mapped to chromosomes in both the A (01, 06, 08) and B (01, 03, 06) genomes. Many of our findings align with previous reports. BG1 and BG4, auxin-regulated genes, positively influence biomass, grain size, and yield in rice^68^. UBP15 and AT1G17145 regulate seed size by modulating cell proliferation in the ovule integument^69,70^. MYB5 plays a role in trichome development and seed coat formation^71^. BRI, predicted on chromosome A06 in B. juncea, enhances cell elongation in Arabidopsis^72^. Mustard seeds are energy rich and store many essential nutrients, including carbohydrates, proteins, and oils, which are crucial for seed development and plant growth. Transporters facilitate nutrient import pathways, ensuring efficient allocation to developing seeds. We could identify several transporter genes, including, each playing a role in sugar and nutrient translocation. SWEETs, are involved in sucrose transporte from leaves to storage organs which are vital for phloem loading and source-to-sink transport^73–75^. AT1G77610 (B06) has been implicated in seed size regulation, controlling bidirectional amino acid translocation and inorganic nitrogen allocation to maintain amino acid homeostasis in ovules and siliques^76^. Previous studies, including those by^77^, have reported small-effect QTLs on LG1, LG6, and LG13 in B. juncea. Additionally, past research has identified QTLs for seed number and size on chromosomes A02^6,61,78^ and A09^61,79,80^, reinforcing the complexity of genetic control over silique traits. Despite these findings, many identified QTLs and predicted gene candidates explain only a fraction of observed variation, suggesting the involvement of additional genetic factors. Current GWAS underlined the importance of SHP2, IND, AG1, and FUL in regulating silique shattering, Barring AG1, all genes contributing to silique dehiscence were predicted on B-genome chromosomes. SHP1 and SHP2 and FUL MADS—box genes control the development of dehiscence zone and valve margins in the siliques. FUL is important for silique morphogenesis. According to our results, SHP2 is also important for determining the number of seeds per silique aside controlling the energy required to rupture a silique. SHP2 has been shown to regulate differentiation of separation layer and lignification of the silique margins^81^. General upregulation of AG1, FUL, and SHP2 was recorded in mustard genotypes with difficult to shatter siliques. In contrast, SHP2 was downregulated in most shattering prone mustard genotypes. FUL controls valve elongation and differentiation of siliques in Arabidopsis^46,81^. Transgenics with constitutive expression of FUL fail to shatter as dehiscence zone failed to form in siliques of transgenic plants^46^. AG1 was not implicated in its role in silique dehiscence. However, SHP1 and SHP2 are AG1 paralogs^82^. AG1 and SHP2 produce non-shatter pods if they are effectively mutated. IND, was also indicated at N2 over the crop seasons for the rupture energy trait. It controls the lignification of margin cells through localised depletion of auxin at the valve margins^83^. Homologs of SHP1/2, FUL, ADPG1, NST1/3 and IND were also reported to associated with shattering in B. juncea and B. napus^84^.
Summarizing, our studies have significantly expanded genetic information regarding silique size, seed size, and silique dehiscence. Several candidate genes, including UBP15, CO, SHP1, SHP2, BG1, BG4, BRI, AT1G12500, AT1G77610, SWEET1, and AT3G14410, have been identified for their roles in silique formation, grain development, and nutrient transport. Additionally, SHP2, FUL**,** IND**,** and AG1 were found to play key roles in silique shattering and seed dispersal.
Materials and methods
Plant material
The present studies involved field testing of 92 genotypes (comprising the B. juncea association panel being maintained at Punjab Agricultural University, Ludhiana. The set included Indian, Australian, Chinese, and east European germplasm lines. Details of the association panel are available at https://static-content.springer.com/esm/art%3A10.1007%2Fs11103-020-01076-x/MediaObjects/11103_2020_1076_MOESM1_ESM.pdf. These genotypes are being maintained by the Punjab Agricultural University.
Field evaluation
Field experiments were planted according to an alpha-lattice design with three doses of nitrogen (N) fertilization. The experiment was replicated twice and repeated for two crop years [designated as Y1 (2015–16) and Y2 (2016–17), respectively] in the farms of Punjab Agricultural University, Ludhiana. The nitrogen (N) doses used were low (N1: 75 kg/ha nitrogen application), recommended (N2: 100 kg/ha nitrogen application), and high (N3: 125 kg/ha nitrogen application). Nitrogen fertiliser was applied in the form of urea (46% N), half after pre-sowing irrigation and the remaining half, 22 days after crop germination. Each germplasm line was raised in four rows, each two metres long per replication. Standard agronomic recommendations were followed for remaining crop inputs^85^. Germplasm lines were assessed for silique length (cm) and seeds per silique, seed size, and silique strength measured as rupture energy required to break open a ripe, dry silique.
Seeds per silique (SPS)
Seeds from the 25 siliques from each of the 92 genotypes comprising the B. juncea association panel were collected, counted, and averaged to obtain an estimate of seeds per silique.
Seed size (SS)
Weight of 1000 seeds was obtained in grams.
Silique length (SL) and rupture energy (RE) needed to shatter a Silique
Silique length and rupture energy required to shatter a pod were estimated with a pendulum machine^86,87^. Rupture energy was used as a measure of shattering resistance. Five physiologically mature siliques per plant were sampled for the experiments. These were detached from the centre of the main racemes of five plants per replication. These Pods (25/germplasm line/replication) were later stored at room temperature in plastic tubes supplemented with coarse silica granules to equilibrate their moisture content. Siliques were then oven-dried at 70 °C for 24 h, before recording silique length and rupture energy (mJ) as described earlier^8^.
Statistical analysis
An analysis of variance (ANOVA) was conducted to infer variation due to genotypes, nitrogen levels, and crop years along with their interactions. GLM (generalized linear model) was applied in alpha-lattice design using SAS software version 9.4. Correlations between levels of nitrogen and across the years of each trait were analysed and visualised using the R-package "Performance Analytics”.
SNP genotyping
We performed genotyping by sequencing^88^ to identify single-nucleotide polymorphisms (SNPs) for association analysis. Experimental protocols used for isolating DNA, genotyping by sequencing (GBS), quality control, SNP identification, and imputation are available elsewhere^55,89^. Software ‘GAPIT’^90^ was used to develop the numerical format of SNP genotypes.
Genome-wide association mapping (GWAS)
The Johnson transformation, as implemented in Minitab v16.0, was used to normalise phenotypic data, whenever required. An arbitrary threshold (−log10(P) > 3) was used to declare the significance of SNP-trait associations. GWAS was performed using MVP (Memory-efficient Visualization-enhanced and Parallel-accelerated Tool) (https://github.com/XiaoleiLiuBio/MVP). GLM (General Linear Model), MLM (Mixed Linear Model), and FarmCPU models were applied with principal components (PCs) as covariates. Manhattan plots were drawn with the MVP tool.
In silico analysis to identify trait-associated candidate genes and LD blocks
Genomic regions (50 kb), upstream or downstream of the apex SNPs, were annotated to envision trait-associated candidate genes. Using A. thaliana gene models, the software Blast2GO Pro^91^ was used. The genes associated with traits of interest were annotated close to the significant SNPs present on chromosomes A01, A05, A08, B01, B04, and B05. Thus, these genes were selected for association with LD blocks using “LDblockShow” (https://github.com/BGI-shenzhen/LDBlockShow).
Candidate gene based regional association mapping
We used an enhanced SNP set for regional association mapping (RAM) to identify SNPs closer to the gene(s) predicted from association analysis. This enhanced SNP set was developed through high-density imputation using a specifically designed mustard imputation reference panel^55,89^. SNPs with P values above a statistical threshold [− log10 (P) > 2] were rated as significant.
Gene expression analysis
Gene expression assays were conducted to validate predicted genes, using a combined analysis of transcriptome sequences available from 18 B. juncea genotypes (RLM-619, Pusa Bold, RLC3, PBR357, DJ22, DJ86, DJ124, DJ127, SJN-3-2, SJR6, SJR13, DT14, DT30, DT138, DT143, CJRB-1661, CJRD1261, JC-210-325, and JC-210-325). This germplasm set was selected to reflect the germplasm variation for silique traits. RNA was extracted from young leaves and siliques harvested from plants raised under a controlled environment (16 h light; 24 °C or 18 °C day and night temperatures; 80% relative humidity) in a Conviron growth chamber. RNA extraction and shotgun sequencing were outsourced. For reference-guided transcriptome assembly, DNASTAR software by Lasergene with default parameters (RNA-Seq®. Version 15.1. DNASTAR. Madison, WI) was used. The assembled genome sequences were functionally annotated using the Blast2GO Pro pipeline against A. thaliana non-redundant protein database from NCBI. Differential expression analysis of predicted genes was performed with the software edgeR 2.4.3^92^. RPKM values (reads per kilobase per million mapped reads) were used to compare gene expression variation for selected set of genes.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kadkol, G. P. Brassica shatter-resistance research update. In Proceedings of the 16th Australian Research Assembly on Brassicas Conference, Ballarat Victoria, 14–16 September 2009, 104–109 (2009).