From peptides to patterning: Redox control of the master regulator PLT2 in Arabidopsis roots
Anna Moseler, Blanca Jazmin Reyes-Hernández

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
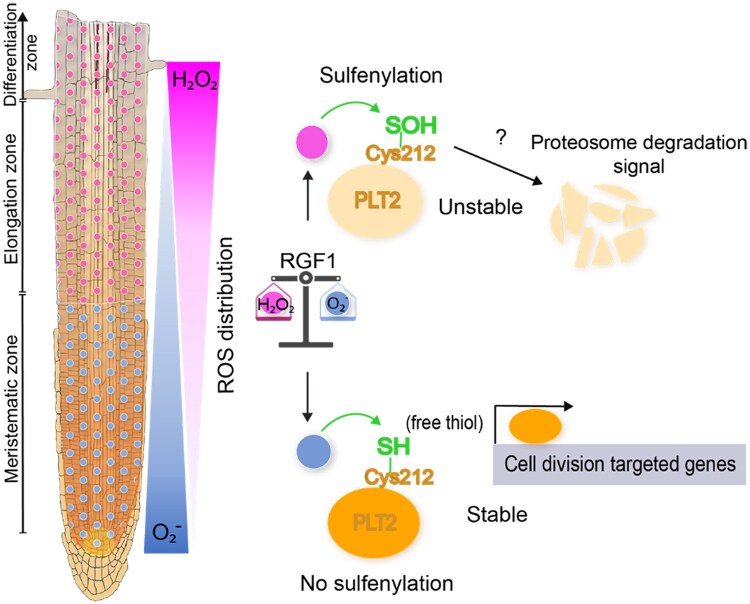 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Stress Responses and Tolerance · Plant nutrient uptake and metabolism
Root systems are essential for anchorage and for the uptake of water and nutrients. Their growth is tightly regulated through coordinated processes such as sustained cell proliferation in the meristematic zone and rapid cell elongation in the elongation zone. Root growth plasticity depends on the balance among these zones and their third developmental zone, the differentiation zone, where cells reach their final size and acquire specialized functions. A key mechanism underlying the coordination of root growth and differentiation is the precise regulation of specific target genes through both activation and repression, a process largely mediated by the PLETHORA (PLT) transcription factors, particularly PLT1 and PLT2, that establish spatial expression gradients across root zones (Aida et al. 2004; Santuari et al. 2016). In recent years, small peptides (typically fewer than 20 amino acids) have emerged as key signaling molecules in root development, acting in a hormone-like manner to modulate these transcriptional programs (Matsubayashi 2014).
A major breakthrough in understanding how PLT gradients are formed came with the discovery of the secreted peptides Root Meristem Growth Factors (RGFs; also referred to as GOLVEN or CLE-like), acting upstream of PLT transcription factors (Matsuzaki et al. 2010). Ten years later, the biochemical connection between RGFs and PLTs was elucidated, establishing that their interaction is mediated through a specific signaling mechanism. RGF1 modulates the balance between the reactive oxygen species (ROS) superoxide (O_2_⁻) and hydrogen peroxide (H_2_O_2_) in the root developmental zones. O_2_⁻ accumulates primarily in the meristematic zone, while H_2_O_2_ accumulates mainly in the differentiation zone; changes in this distribution in turn alter the stability of PLT2 (Yamada et al. 2020). However, the mechanism behind PLT2 stabilization remained unclear. Hsiao et al. (2025) now show that RGF1 signaling-dependent sulfenylation of a cysteine (Cys) in PLT2 is crucial for PLT2 stability.
PLT1 and PLT2 act redundantly to maintain root growth. While plt1 and plt2 single mutants exhibit slight meristem shrinkage, the plt1 plt2 double mutant shows a pronounced decreased root meristem (Aida et al. 2004). When wild-type and plt1 plants are treated with RGF1, the meristem expands but not in plt2. Using PLT-GFP lines, Hsiao and colleagues showed that RGF1 treatment increases the PLT2 distribution at the root tip, while PLT1 remains limited and slightly decreases. This indicates that PLT2 is the major contributor to the RGF1 signaling pathway.
In the next step, the researchers investigated how the ROS modulated by RGF1 impact PLT2. Since thiol (−SH) groups are common ROS targets, the authors replaced one or both Cys residues at positions 112 and 212 by serine. They complemented the plt2 mutant with the respective C112S, C212S and C112S/C212S PLT2 variants and observed their root responses to RGF1. The gPLT2^C212S^-GFP and gPLT2^C112S^/^C212S^-GFP expressed in plt2 lines showed broader PLT2 localization and larger meristematic zones, even without exogenous RGF1 application, highlighting a pivotal role of the 212th Cys for PLT2 stability. Monitoring ROS distribution in gPLT2^C212S^-GFP lines showed that RGF1 increased O_2_⁻ and decreased H_2_O_2_, especially at the boundary between the meristematic and elongation zones. Substituting the 212th Cys enhances PLT2 stability under oxidative conditions, suggesting that the redox state of the 212th Cys is crucial for its stability and function (Fig.). Reverse transcription quantitative PCR confirmed unchanged PLT2 transcript levels, while immunoblots revealed marked protein levels increased in the C212S lines after RGF1 treatment. Using a nonmobile PLT2-3xGFP line, the authors still observed expanded localization, excluding any effect due to PLT2 cell-to-cell movement. Together, these data demonstrate that Cys212 is essential for ROS-dependent PLT2 stabilization, indicating that the effect of RGF1 on ROS acts upstream of PLT to promote meristem expansion.
The plt2 mutant expressing either gPLT2-GFP or the corresponding C212S variant were further used to analyze the GFP signal intensities in different developmental zones. The authors detected a more stable signal for gPLT2^C212S^-GFP in the elongation zone, where higher H_2_O_2_ levels were detected, compared to the gPLT2-GFP. Additionally, the team showed that after exogenous application of H_2_O_2_, the stabilization of PLT2 leads to an increase in root meristem size, highlighting that H_2_O_2_ mainly targets the 212th Cys to modulate PLT2 stability. Generally, the reaction of H_2_O_2_ typically oxidizes thiols to sulfenic acid, but excess of H_2_O_2_ can cause overoxidation to sulfinic or sulfonic acid, leading to protein degradation. To detect specific modifications of 212th Cys of PLT2 in vivo, the authors isolated PLT2-GFP and found that this residue was more sulfenylated after H_2_O_2_ treatment, indicating that H_2_O_2_ promotes sulfenylation to modulate PLT2 stability.
The authors then examined how the C212S variant affects target gene expression. RNA-seq analysis of root tips revealed over 8,500 differentially expressed genes (DEGs) in the gPLT2-GFP line compared to the plt2 mutant and over 10,500 DEGs in the gPLT2^C212S^-GFP line, indicating that the PLT2^C212S^ variant has a stronger impact on downstream gene regulation than the wild-type PLT2. Comparison with a previous root developmental zone–specific transcriptome analysis (Li et al. 2016) revealed enhanced modulation of zone-specific genes by the PLT2^C212S^ variant: 883 out of 1,098 genes were upregulated in the meristematic zone, and 339 of 377 genes were downregulated in the elongation zone. Gene ontology (GO) analysis showed enrichment of cell proliferation-related terms among upregulated genes and cell differentiation and cell wall biogenesis among the downregulated genes. These data indicate that the stabilization of PLT2 amplifies its transcriptional regulation, potentially shifting the balance toward meristem activity over differentiation.
In summary, this work confirms previous studies showing the role of the RGF1 peptide in regulating PLT2 protein stability through modulation of ROS levels during root meristem development and further demonstrates that PLT2 protein stability is controlled post-translationally by sulfenylation of the 212th Cys residue by H_2_O_2_, thereby limiting its spread in the elongation zone. Further studies are needed to unravel how individual cells perceive and integrate the concentration-dependent information provided by RGFs to generate precise root meristem patterning during root development. Another important question is how the sulfenylation of PLT2 induces the degradation of the protein. Furthermore, new tools like genetically encoded sensors for H_2_O_2_ can help to further dissect H_2_O_2_ changes dynamically and at high resolution (Nietzel et al. 2019). Nevertheless, the knowledge obtained in this study can support the development of strategies to enhance root development in adverse environmental conditions. For instance, crops expressing modified versions of PLT2 that are less sensitive to oxidative degradation could maintain root function during prolonged stress, potentially helping sustain plant productivity.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aida M, Beis D, Heidstra R, Willemsen V, Blilou I, Galinha C, Nussaume L, Noh Y-S, Amasino R, Scheres B. The PLETHORA genes mediate patterning of the Arabidopsis root stem cell niche. Cell. 2004:119(1):109–120. 10.1016/j.cell.2004.09.01815454085 · doi ↗ · pubmed ↗
- 2Hsiao Y, Shiue S, Yen M, Lai J, Yamada M. Sulfenylation of a cysteine modulates PLT 2 protein stability through ROS under the RGF 1 signal in root meristem development. Plant Physiol. 2025.
- 3Li S, Yamada M, Han X, Ohler U, Benfey PN. High-resolution expression map of the Arabidopsis root reveals alternative splicing and linc RNA regulation. Dev Cell. 2016:39(4):508–522. 10.1016/j.devcel.2016.10.01227840108 PMC 5125536 · doi ↗ · pubmed ↗
- 4Matsubayashi Y . Posttranslationally modified small-peptide signals in plants. Annu Rev Plant Biol. 2014:65(1):385–413. 10.1146/annurev-arplant-050312-12012224779997 · doi ↗ · pubmed ↗
- 5Matsuzaki Y, Ogawa-Ohnishi M, Mori A, Matsubayashi Y. Secreted peptide signals required for maintenance of root stem cell niche in Arabidopsis. Science. 2010:329(5995):1065–1067. 10.1126/science.119113220798316 · doi ↗ · pubmed ↗
- 6Nietzel T, Elsässer M, Ruberti C, Steinbeck J, Ugalde JM, Fuchs P, Wagner S, Ostermann L, Moseler A, Lemke P, et al The fluorescent protein sensor ro GFP 2-Orp 1 monitors in vivo H 2O 2 and thiol redox integration and elucidates intracellular H 2O 2 dynamics during elicitor-induced oxidative burst in Arabidopsis. New Phytol. 2019:221(3):1649–1664. 10.1111/nph.1555030347449 · doi ↗ · pubmed ↗
- 7Santuari L, Sanchez-Perez GF, Luijten M, Rutjens B, Terpstra I, Berke L, Gorte M, Prasad K, Bao D, Timmermans-Hereijgers JLPM, et al The PLETHORA gene regulatory network guides growth and cell differentiation in Arabidopsis roots. Plant Cell. 2016:28(12):2937–2951. 10.1105/tpc.16.0065627920338 PMC 5240741 · doi ↗ · pubmed ↗
- 8Yamada M, Han X, Benfey PN. RGF 1 controls root meristem size through ROS signalling. Nature. 2020:577(7788):85–88. 10.1038/s 41586-019-1819-631801996 PMC 6930331 · doi ↗ · pubmed ↗