Nitrogen demand and availability: relative roles in driving C3 plant responses to elevated CO2
Xuan Hu, Mirindi Eric Dusenge

TL;DR
This paper explores how nitrogen demand and availability affect how C3 plants respond to increased CO2 levels.
Contribution
It introduces a framework linking nitrogen demand, availability, and acquisition strategies to plant responses under elevated CO2.
Findings
Nitrogen demand and availability are key factors influencing plant growth under elevated CO2.
Acquisition strategies determine how effectively plants can respond to higher CO2 levels.
C3 plants show varied responses depending on nitrogen dynamics and environmental conditions.
Abstract
This article comments on: Perkowski EA, Ezekannagha E, Smith NG. 2025. Nitrogen demand, availability, and acquisition strategy control plant responses to elevated CO2. Journal of Experimental Botany 76, 2908–2923 https://doi.org/10.1093/jxb/eraf118
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
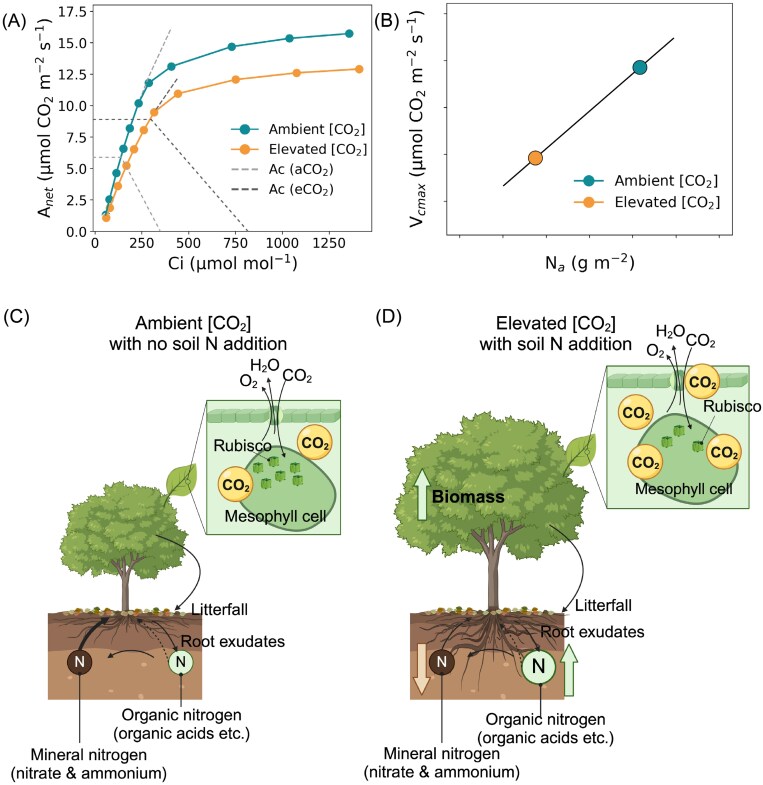 Fig. 1
Fig. 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant responses to elevated CO2 · Plant Water Relations and Carbon Dynamics · Atmospheric chemistry and aerosols
This article comments on:
Perkowski EA, Ezekannagha E, Smith NG. 2025. Nitrogen demand, availability, and acquisition strategy control plant responses to elevated CO_2_. Journal of Experimental Botany 76, 2908–2923 https://doi.org/10.1093/jxb/eraf118
C _ 3 _ plants generally show increased photosynthesis and whole-plant productivity in response to elevated atmospheric CO _ 2 _ concentrations [CO _ 2 _ ]. While it is widely recognized that nitrogen (N) mediates the responses of plants to elevated [CO _ 2 _ ], there has been ongoing debate on the relative importance of the N availability versus metabolic N demand in influencing the responses of leaf-level C _ 3 _ photosynthesis to elevated atmospheric [CO _ 2 _ ]. In this issue of the Journal of Experimental Botany, Perkowski et al. (2025) provide empirical evidence that N demand strongly dictates the responses of leaf-level photosynthesis while N availability substantially enhances whole-plant productivity in response to elevated [CO _ 2 _ ].
Impact of elevated [CO2] on C3 plants
Atmospheric [CO_2_] has increased by 52% compared with the pre-industrial level (Friedlingstein et al., 2025), and is projected to continue to rise in the near future (Arias et al., 2023). Among the main environmental drivers of global change (e.g. CO_2_, temperature, ozone, and drought), elevated atmospheric [CO_2_] and its impact on plants have received relatively more attention. As a result, we have a fairly good understanding of how elevated [CO_2_] affects plants, spanning from the leaf level to the ecosystem scale, and ranging from short-term (minutes) to decade-long studies (Curtis, 1996; Leakey et al., 2009; Norby et al., 2010; Warren et al., 2015). Since plants use CO_2_ as the main substrate during photosynthesis, rising atmospheric [CO_2_] generally increases rates of net photosynthesis on both short -and long-term time scales (Curtis, 1996; Ainsworth and Long, 2005; Leakey et al., 2009; Way et al., 2015). However, the initial stimulation diminishes over time as plants acclimate to elevated [CO_2_] (Curtis, 1996; Ainsworth and Long, 2005; Leakey et al., 2009). At the leaf level, the acclimation of photosynthesis to elevated [CO_2_] involves several changes in physiology and biochemistry (Ainsworth and Long, 2005; Ainsworth and Rogers, 2007; Leakey et al., 2009). These changes mainly include decreases in stomatal conductance and the maximum carboxylation rate of Rubisco (Vcmax), which is a key enzyme during the CO_2_ fixation process. The decrease in Vcmax is driven by an imbalance between the source (leaves) and sink (other parts of the plant), caused by a relatively higher accumulation of sugars in the leaves under elevated [CO_2_]. This accumulation initiates hexokinase signalling pathways, which suppress Rubisco transcription, leading to reduced Rubisco content (Moore et al., 1999; Ainsworth and Rogers, 2007), which is reflected in decreases in Vcmax (Ainsworth and Long, 2005).
The role of N in C3 plant acclimation to elevated [CO2]
Photosynthetic acclimation to elevated [CO_2_] has been shown to be stronger in N-poor systems (Ainsworth and Long, 2005). Studies using whole-tree chambers (WTCs) and free air CO_2_ enrichment (FACE) technologies show that initial net primary productivity (NPP) stimulation decreases over time due to N limitation (Oren et al., 2001; Luo et al., 2004; Norby et al., 2010; Warren et al., 2015). In a WTC study, elevated [CO_2_] did not significantly affect tree growth at an N-poor site (Sigurdsson et al., 2013). Similarly, controlled studies on small plants showed stronger acclimation in plants grown in small pots (<5 litres) or with insufficient fertilization (Sage, 1994; Curtis and Wang, 1998). These findings further highlight the importance of N in driving the long-term acclimation of C_3_ plants to elevated [CO_2_]. The previously observed N-driven reduction in the stimulation of elevated [CO_2_] led to the development of the progressive nitrogen limitation (PNL) framework (Luo et al., 2004). The PNL framework suggests that N becomes progressively limiting because it is increasingly stored in the organic matter of additional long-lived plant biomass and soil organic matter, both enhanced by elevated [CO_2_]. Without N fertilization, this process gradually reduces the N available for further plant uptake over time. While the PNL framework explains commonly observed results in N-limited systems quite well, it does not adequately explain the reduction in Vcmax and associated leaf N commonly observed in elevated [CO_2_] studies with adequate plant soil N availability (Ainsworth and Long, 2005; Dusenge et al., 2024). By contrast, the eco-evolutionary optimality (EEO) theoretical framework suggests that the leaf-level demand to build and maintain the photosynthetic machinery under given environmental conditions is the main driver of leaf N content (Harrison et al., 2021). The EEO theory proposes that investment in photosynthetic machinery is optimized for prevailing environmental conditions (e.g. light and temperature) in a way that minimizes resource use (N and water), making it less dependent on soil N availability (Smith et al., 2019). Therefore, under elevated [CO_2_], plants invest relatively less N to build Rubisco, as it can optimally fix more [CO_2_] under these conditions. Consequently, plants grown under elevated [CO_2_] should exhibit reduced leaf N due to the decreased demand for Vcmax (Smith and Keenan, 2020), regardless of soil N availability. This is mainly because the Rubisco enzyme accounts for up to 50% of the total leaf N (Spreitzer and Salvucci, 2002).
The new study by Perkowski et al. (2025) systematically tested the relative importance of the PNL versus EEO frameworks in the context of plant responses to elevated [CO_2_]. The authors grew soybean plants (Glycine max) at nine N fertilization levels under either an ambient atmospheric [CO_2_] of 420 ppm or an elevated atmospheric [CO_2_] of 1000 ppm. Since soybean can fix atmospheric N via its symbiotic relationship with N-fixing bacteria, half of the plants were allowed to fix atmospheric N, while the other half were sterilized to disable this ability, in order to also investigate the role of the N acquisition strategy. As expected from the EEO framework, leaf N overall decreased in plants grown under elevated [CO_2_]. While soil N fertilization increased leaf N, this stimulation was less pronounced under elevated [CO_2_], leading to a greater reduction in leaf N at higher soil N levels compared with ambient [CO_2_]. Similarly, Vcmax at 25 °C (Vcmax25) decreased by an average of 16% under elevated [CO_2_], and these reductions were not modified by soil N fertilization treatments. Rates of net photosynthesis at prevailing growth [CO_2_] (Agrowth) were still higher in elevated [CO_2_] plants compared with their ambient [CO_2_] counterparts. As a result, photosynthetic N use was substantially increased by elevated [CO_2_]. As previously observed in FACE studies (Ainsworth and Long, 2005), total leaf area and total biomass increased under elevated [CO_2_]. While soil N fertilization also increased these traits, the increases were more pronounced under elevated [CO_2_] than in ambient [CO_2_] conditions, as would be expected from the PNL framework. Soil N fertilization had a stronger effect on total leaf area and total biomass in uninoculated compared with inoculated plants. The study by Perkowski et al. (2025) also demonstrated that elevated [CO_2_] increased below-ground biomass as a strategy to acquire more N, a pattern that was more pronounced in uninoculated plants. Additionally, increased soil N fertilization reduced the carbon costs to acquire N more significantly in uninoculated than in inoculated plants. Surprisingly, inoculation with symbiotic N-fixing bacteria did not overall influence either leaf-level or whole-plant responses to elevated [CO_2_].
Implications and future prospects
The study by Perkowski et al. (2025) reconciled the PNL and EEO frameworks by demonstrating that the EEO framework explains leaf-level responses to elevated [CO_2_], while the PNL framework explains whole-plant responses. Specifically, they showed that leaf demand for building a photosynthetic machinery that is adjusted to the plant’s growing environment drives the common enhancement of Agrowth under elevated [CO_2_], despite apparent decreases in photosynthetic capacity (Vcmax) that were unaltered by soil N availability (Fig. 1A, B). The authors found a stronger reduction in leaf N content under elevated [CO_2_] when N was more available, suggesting that plants reallocate N to other non-photosynthetic metabolic processes, ultimately contributing to enhanced plant growth with higher photosynthetic N use efficiency. In contrast, their results show that the PNL framework nicely explains the enhancement of whole-plant responses to elevated [CO_2_] under higher soil N availability, as evidenced by substantial increases in leaf and total biomass (Fig. 1C, D). Findings from this study are consistent with recent results from a field study on mature boreal conifers exposed to both warming and elevated [CO_2_] (Dusenge et al., 2024). In a boreal peatland dominated by tamarack and black spruce, 2 years of both below-ground and above-ground warming enhanced plant-available soil N (Iversen et al., 2023), which led to an increase in leaf N across the warming treatments. However, despite this increase in leaf N, it did not affect the acclimation of Vcmax25 and leaf N (i.e. decreases) to 1 year of elevated [CO_2_], which remained consistent across all warming treatments (Fig. 1A, B). While these field observations suggest that the results from Perkowski et al. (2025) may be applicable to other plant functional types (PFTs), they still need to be tested across a wider range of PFTs, with longer treatment durations, and ideally under field conditions, before any definitive conclusions can be drawn. We also need a deeper understanding of plant N allocation under elevated [CO_2_]. While logistically and financially challenging, these research questions could be integrated into ongoing FACE experiments to improve modelling of the coupling of carbon and N cycles under current and future climatic conditions.
Conceptual representation of C3 plant responses to elevated [CO2] and soil N availability. (A) Response of net photosynthesis at different air CO2 concentrations, also known as the A–Ci curve. The data used to construct this curve are from the study by Dusenge et al. (2024), and they focus on black spruce (Picea mariana) needles measured at a leaf temperature of 25 °C. In this study, mature black spruce trees were exposed to both ambient atmospheric [CO2] and elevated [CO2] (ambient+500 ppm) for 1 year. The observed down-regulation of Vcmax (~19%) is comparable with the 16% observed in Perkowski et al. (2025). (B) Decreases in Vcmax are related to decreases in leaf N (Ainsworth and Long, 2005). (C, D) Schematic representation of the effects of elevated [CO2] and high plant-available soil N on the leaf and whole-plant level. At the leaf-level, Rubisco content decreases, regardless of soil N availability, because the leaf can fix more CO2 molecules with fewer Rubisco enzymes. At the whole-plant level, exposure to elevated [CO2] coupled with high plant soil N availability leads to increased biomass (leaves, stems, and roots), as shown by Perkowski et al. (2025). Specifically, under elevated [CO2], plants produce more leaves with reduced Vcmax and N per leaf. These results also highlight how the EEO framework partly acknowledges the importance of plant soil N availability for whole-plant responses, where plants use this extra N to build more optimally coordinated leaves and other plant organs.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ainsworth EA , Long SP. 2005. What have we learned from 15 years of free‐air CO 2 enrichment (FACE)? A meta‐analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO 2. New Phytologist 165, 351–372.15720649 10.1111/j.1469-8137.2004.01224.x · doi ↗ · pubmed ↗
- 2Ainsworth EA , Rogers A. 2007. The response of photosynthesis and stomatal conductance to rising [CO 2]: mechanisms and environmental interactions. Plant, Cell & Environment 30, 258–270.10.1111/j.1365-3040.2007.01641.x 17263773 · doi ↗ · pubmed ↗
- 3Arias PA , Bellouin N, Coppola E, et al 2023. Technical summary. In: Intergovernmental Panel on Climate C, ed. Climate Change 2021—The Physical Science Basis: Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 35–144.
- 4Curtis PS. 1996. A meta‐analysis of leaf gas exchange and nitrogen in trees grown under elevated carbon dioxide. Plant, Cell & Environment 19, 127-137.
- 5Curtis PS , Wang X. 1998. A meta-analysis of elevated CO 2 effects on woody plant mass, form, and physiology. Oecologia 113, 299–313.28307814 10.1007/s 004420050381 · doi ↗ · pubmed ↗
- 6Dusenge ME , Warren JM, Reich PB, et al 2024. Photosynthetic capacity in middle‐aged larch and spruce acclimates independently to experimental warming and elevated CO 2. Plant, Cell & Environment 47, 4886–4902.10.1111/pce.1506839101396 · doi ↗ · pubmed ↗
- 7Friedlingstein P , O'Sullivan M, Jones MW, et al 2025. Global Carbon Budget 2024. Earth System Science Data 17, 965–1039.
- 8Harrison SP , Cramer W, Franklin O, et al 2021. Eco-evolutionary optimality as a means to improve vegetation and land-surface models. New Phytologist 231, 2125–2141.34131932 10.1111/nph.17558 · doi ↗ · pubmed ↗