Endogenous formation of phosphatidylhomoserine in Escherichia coli through phosphatidylserine synthase
Elise Zi Qi Ng, Eunju Lee, Shu-Sin Chng, Jungwook Kim, Xue Li Guan

TL;DR
This study reveals that Escherichia coli can produce a new phospholipid, phosphatidylhomoserine, using an enzyme called phosphatidylserine synthase A.
Contribution
The paper identifies phosphatidylhomoserine as an endogenously produced phospholipid in E. coli through phosphatidylserine synthase A.
Findings
Phosphatidylhomoserine (PHS) is endogenously produced in E. coli by phosphatidylserine synthase A (PssA).
E. coli and other Enterobacteriaceae can convert PHS into phosphatidylpropanolamine.
PHS production in E. coli reduces phosphatidylglycerol and phosphatidylethanolamine levels due to shared metabolic intermediates.
Abstract
Biological membranes, which comprise proteins, lipids, and glycans, serve as essential gatekeepers protecting cells from the external environment. In bacteria, phospholipids are a major class of membrane lipids, whose biology has extensively been studied in the Gram-negative organism Escherichia coli. As an adaptive mechanism, E. coli dynamically remodels its phospholipids in response to its environment, which may involve alterations of the structures and/or levels of existing lipids or the incorporation of exogenous substrates to form new phospholipid classes. Intriguingly, an unknown lipid was detected in E. coli and other Enterobacteriaceae. Detection of this lipid in E. coli grown in minimal media suggested its production using an endogenous metabolite. By coupling liquid chromatography mass spectrometry and metabolic incorporation, the lipid was identified as phosphatidylhomoserine…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
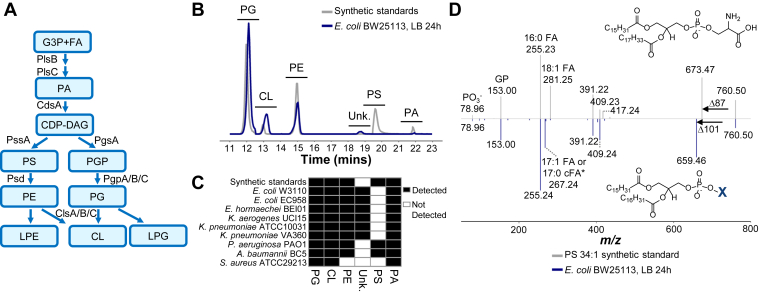 Figure 1
Figure 1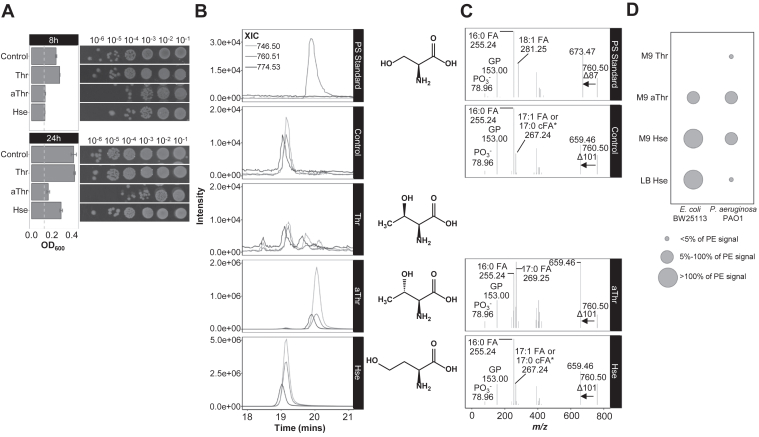 Figure 2
Figure 2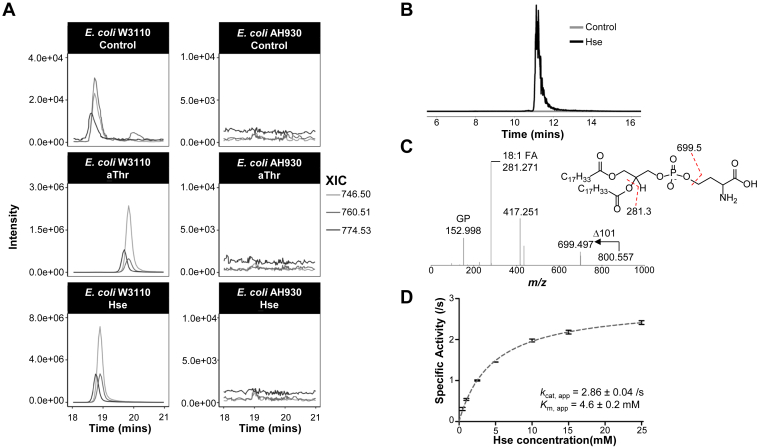 Figure 3
Figure 3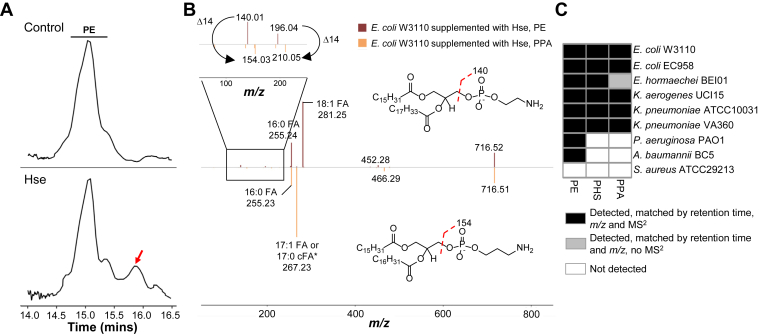 Figure 4
Figure 4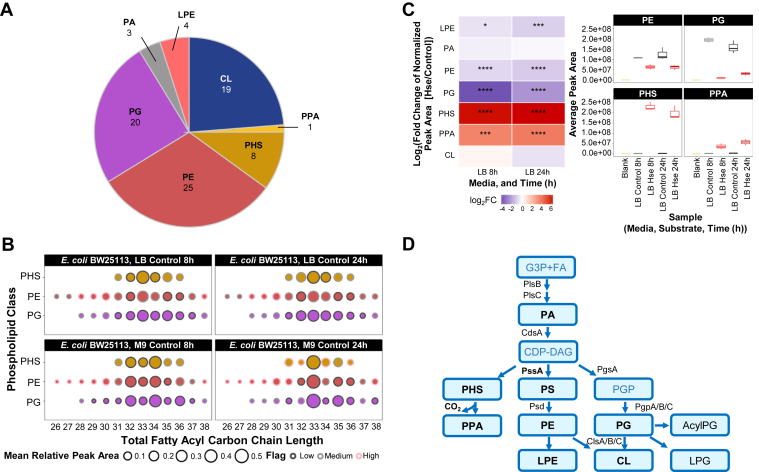 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeonatal Health and Biochemistry · Biochemical and Molecular Research · Sphingolipid Metabolism and Signaling
Phospholipids, besides lipopolysaccharides, glycans, and proteins, are the major components of Gram-negative bacterial membranes which form a protective barrier from the extracellular environment (1) and play important roles in diverse biological processes (2, 3, 4). As a unicellular organism, bacteria have evolved strategies to cope with its varying environment, and phospholipid remodeling presents one of the key mechanisms for rapid survival responses to perturbations (5, 6). Studies on the model organism, Escherichia coli, have laid a strong foundation for our current understanding of phospholipid metabolism, transport, and functions in Gram-negative bacteria (3, 4, 7, 8, 9, 10, 11).
As a diderm, E. coli has two membrane bilayers, which comprise of an outer membrane enriched in lipopolysaccharides, while phospholipids form the main lipid component of the inner membrane. Compositionally, phosphatidylethanolamine (PE) accounts for up to 70% of the total phospholipid content in E. coli, followed by phosphatidylglycerol (PG), cardiolipin (CL), and phosphatidic acid (PA) (7). Phospholipid synthesis in E. coli starts with the sequential acylation of glycerol-3-phosphate by acyltransferases, PlsB and PlsC to form PA (12, 13, 14). Cytidine diphosphate diacylglycerol (CDP-DAG), which is next formed from PA by CDP-DAG synthase, CdsA (15, 16), is the metabolic branch point for the formation of the two major diacyl phospholipids, PE and PG (Fig. 1A) (11, 17). The primary pathway for PE synthesis in E. coli is through phosphatidylserine (PS) decarboxylation (11). PS is produced through the transfer of serine to CDP-DAG by phosphatidylserine synthase, PssA (18, 19), followed by decarboxylation to yield PE through the actions of phosphatidylserine decarboxylase, Psd (20, 21, 22). PG, on the other hand, is formed from the metabolic intermediate, phosphatidylglycerol phosphate (PGP), synthesized by phosphatidylglycerol synthase, PgsA (23). PGP is in turn dephosphorylated to form PG and subsequently CL through the actions of PGP phosphatases, PgpA/B/C, and cardiolipin synthases, ClsA/B/C respectively (23, 24, 25, 26). Besides their main activities, it is unsurprising that enzymes can exhibit additional activities, either due to the presence of multiple functional domains or substrate promiscuity (27). For instance, PssA is able to incorporate phosphoglycerol, which ultimately leads to the formation of PG, albeit at low level (28). In E. coli, nonclassical phospholipids, phosphatidylalcohols, can be formed with the introduction of exogenous primary alcohols (29), through the phospholipase D (PLD) activity of ClsB (30). When cultured in the presence of mannitol, E. coli was also able to form two novel lipids, phosphatidylmannitol and diphosphatidylmannitol, contributed by the activity of cardiolipin synthase (31).Figure 1**Escherichia coli phospholipid pathway and discovery of a previously uncharacterized lipid.**A, general phospholipid pathway of E. coli. B, normal phase chromatography separation of E. coli phospholipid fraction (blue) and phospholipid synthetic standards (gray), for classical phospholipid classes, PE, PG, CL, PA, PS, found in Gram-negative bacteria. Peaks represent extracted ion chromatogram (XIC) of the main or major ions of each lipid class. An unknown peak in E. coli eluting close to (but before) PS 34:1 synthetic standard was detected. C, conservation of the unknown lipid in Enterobacteriaceae species. Lipids were isolated from various bacterial species and analyzed using LC/MS^2^. The heatmap represents the absence (white) or presence (black) of a phospholipid class, based on the analytical detection in this study. D, MS^2^ spectra of PS 34:1 (16:0/18:1) synthetic standard (top spectrum, gray) and the unknown ion detected in E. coli (bottom, inverted spectrum, blue). Fragmentation of PS-generated diagnostic ions from the neutral loss of serine (87 Da), phosphate (m/z 78.96), glycerophosphate and fatty acids, 18:1 (m/z 281.25) and 16:0 (m/z 255.23). For the ion found in E. coli, it exhibited a neutral loss of 101 Da. Other diagnostic ions include the signature of phosphate, glycerophosphate, and fatty acids, 16:0 (m/z 255.24) and 17:1 or c17:0 (m/z 267.24). Inset showed the structures of PS and partial structure of the unknown, with its headgroup ‘X’ to be further characterized. MS^2^ data was derived from one sample and is representative of the three biological replicates. CDP-DAG, cytidine diphosphate diacylglycerol; CdsA, phosphatidate cytidylyltransferase; cFA, cyclopropane fatty acid; CL, cardiolipin; ClsA/B/C, cardiolipin synthase A/B/C; Da, Dalton; FA, fatty acid; G3P, glycerol-3-phosphate; GP, glycerophosphate; LB, Luria-Bertani broth (Miller); LC/MS^2^, liquid chromatography-high resolution tandem mass spectrometry; LPE, lysophosphatidylethanolamine; m/z, mass-to-charge ratio; PA, phosphatidic acid; PE, phosphatidylethanolamine; PG, phosphatidylglycerol; PGP, phosphatidylglycerol phosphate; PgpA/B/C, phosphatidylglycerophosphatase A/B/C; PgsA, phosphatidylglycerol phosphate (PGP) synthase; PlsB, glycerol-3-phosphate acyltransferase; PlsC, 1-acyl-sn-glycerol-3-phosphate acyltransferase; PO3, phosphate; PS, phosphatidylserine; Psd, phosphatidylserine decarboxylase; PssA, phosphatidylserine synthase; Unk., unknown. ∗The analytical method does not distinguish between mono-unsaturated fatty acid and cyclopropane fatty acid.
The lipid composition of a unicellular bacterium is clearly shaped by the combination of its enzymatic machinery as well as substrate availability. In vitro substrate characterization with purified enzymes are fundamental and powerful techniques which have contributed significantly to our current understanding of the specific activities of lipid enzymes in diverse organisms. However, biological systems are inherently complex, and both intracellular and extracellular conditions can influence enzymatic activities, ultimately changing the biochemical profile of a bacterium (29). In addition, key lipid precursors such as serine and acetate intersect with pathways involved in protein synthesis and carbon metabolism (32). As substrates are funneled into different biochemical pathways, the lipid profile of a cell is therefore dependent on both the environmental conditions and its physiological state. Liquid chromatograph mass spectrometry (LC/MS) offers unprecedented resolution and sensitivity to probe the biochemistry and metabolic state of a cell. Indeed, whole cell lipidome analysis is a complementary approach to understand global cellular biochemistry (33, 34). This systems-scale approach had been applied by various groups to study lipid metabolism and functions in bacteria, and PE, PG, and CL had been coherently reported as the major phospholipids found in E. coli (29, 35, 36). However, there are existing variations in other components detected, which can arise from the bioanalytical methods, bacterial strains, and culture conditions used. For instance, the presence of phosphatidylserylglutamate (37), phosphatidylmannitol (31), and phosphatidylalcohols (30, 38) had been described in E. coli. Nevertheless, not all nonclassical phospholipids can be attributed to the existing biochemical machinery found in E. coli, and further works will be required to confirm their true existence and biological functions.
In this study, we described the characterization of two novel endogenous phospholipids present in E. coli and other Enterobacteriaceae. Using liquid chromatography-high resolution tandem mass spectrometry (LC/MS^2^), substrate incorporation, genetics, and enzyme biochemistry, we provided structural and biochemical evidence of phosphatidylhomoserine (PHS) in E. coli. PHS is formed endogenously through the activity of EcPssA. The ability of EcPssA to use L-homoserine (Hse) as a substrate to form PHS was demonstrated in cells, as well as using purified enzyme system in vitro. E. coli can further decarboxylate PHS to form phosphatidylpropanolamine (PPA) endogenously, albeit at very low levels. When exposed to L-homoserine (Hse), the bacterium accumulated PHS and PPA, with reduction in PG and PE levels potentially due to the competition of the common building block, CDP-DAG. This work renewed our fundamental understanding of E. coli lipid metabolism at the cellular level; specifically, our findings re-established the baseline phospholipidome of E. coli and revealed the previously uncharacterized substrates of EcPssA. Furthermore, our works also demonstrated the type of substrates and degree of incorporation into phospholipids depend on the bacterial species and the type of growth media, highlighting environmentally driven plasticity and complexity of bacterial lipid metabolism.
Results
Baseline phospholipidome of E. coli harbors an unknown, which is conserved in Enterobacteriaceae
The phospholipidome of E. coli cultured in LB broth (Miller) (LB) was analyzed using normal phase chromatography with tandem mass spectrometry (MS^2^), which separated phospholipids by their headgroups (Fig. 1B). Consistent with existing reports (29, 35, 36), PE, PG, and CL are the major phospholipid classes detected in E. coli strain BW25113 under aerobic conditions in LB, a standard laboratory media. PA was detected at low levels, and an additional peak was observed, which eluted before but close to PS synthetic standard, with ions sharing the same mass-to-charge ratio (m/z) as PS 34:1. This peak was consistently found in three E. coli strains (E. coli BW25113, E. coli W3110, and E. coli EC958), as well as other Enterobacteriaceae (Klebsiella pneumoniae, Klebsiella aerogenes, Enterobacter. hormaechei) (Fig. 1C). However, it was not detectable in non-Enterobacteriaceae Gram-negative bacteria, Acinetobacter baumannii or Pseudomonas aeruginosa; instead, PS was detected (Fig. 1B). Both the unknown and PS were absent in the Gram-positive bacterium, Staphylococcus aureus.
Based on the difference in retention time from PS, we postulated E. coli and other Enterobacteriaceae can produce another class of phospholipids. MS^2^ analyses revealed that in contrast to a neutral loss of 87 Da (Da), arising from the loss of serine from PS, the fragmentation of the ion found in E. coli formed a neutral loss of 101 Da (Fig. 1D). The E. coli lipid possessed the signature phosphate (m/z 78.96) and glycerophosphate (m/z 153.00) fragments, which were also found in the PS fragmentation profile (Fig. 1D). The major form of this ion contained two fatty acyls, 16:0 fatty acid and a 17-carbon fatty acid with either one double bond or cyclopropane modification. In the parasite, Toxoplasma gondii (39), as well as human blood (40), phosphatidylthreonine (PT), which forms a neutral loss of 101 Da during fragmentation, had been previously described (40). Indeed, threonine is 14 Da larger than serine, and intuitively as one of the essential amino acids produced in the cells, it should be a readily available substrate. PT in T. gondii is synthesized by PT synthase (TgPTS) (39); however, no homolog of TgPTS is found in E. coli, based on BLAST sequence alignment query. One possibility would be to form PT in E. coli through the activity of PssA. However, Kanfer and Kennedy demonstrated that PssA does not harbor observable activity when threonine was used as a substrate (11).
Identification of PHS in E. coli through substrate incorporation in whole bacterium
To determine if E. coli can incorporate threonine or other metabolites which share the same molecular weight directly into its phospholipids, we next established bacterial culture conditions using M9, a salt-based minimal media, to introduce specific substrates in a defined fashion (Figs. 2A and S1). This approach built on the ability of bacteria to transport amino acids and derivatives using common transporters (41). By providing the appropriate substrate in excess, we postulate the bacterium will accumulate the lipid of interest through its endogenous lipid biosynthetic machinery.Figure 2**Identification of phosphatidylhomoserine through metabolic substrate incorporation in E. coli.**A, viability of Escherichia coli BW25113, grown in M9 minimal media supplemented with 0.1% (w/v) of substrates, L-threonine, L-allo-threonine, and L-homoserine, which are 14 Da larger than serine. Each spotting condition is represented with one technical spotting replicate within a set of biological replicates that includes two culture replicates, with three technical replicates for spotting. A total of three biological replicates were performed (n = 3). A_600_ bar graph was derived from the equivalent biological experiment and is representative of three biological replicates. Left: A_600_ (Gray dashed line: A_600_ of blank media control). Right: colony formation. B, extracted ion chromatograms (XIC) of three major ions of the unknown lipid class found endogenously in E. coli. Lipids analyzed were derived from E. coli BW25113 grown in M9 minimal media, supplemented with the substrates, L-threonine, L-allo-threonine, and L-homoserine (0.1% (w/v)). Data represented are from 8-h cultures. Treatment with L-allo-threonine and L-homoserine resulted in the accumulation of lipids at distinct retention times, with the latter co-eluting with the endogenous lipids found in E. coli. L-threonine treatment did not result in any incorporation, evident from the signals in the XIC. C, MS^2^ spectra of E. coli BW25113 lipids with m/z 760.50 under control (endogenous) and substrate-incorporated conditions. Both L-allo-threonine and L-homoserine incorporated lipids which formed a neutral loss of 101 Da. The XIC and spectra were derived from one set of experiment and are representative of three biological replicates. D, organism- and media-dependent incorporation of substrates. Both E. coli BW25113 and P. aeruginosa PAO1 were cultured in M9 minimal media and treated with the various substrates, L-threonine, L-allo-threonine, and L-homoserine (0.1% (w/v)). L-homoserine incorporation was additionally determined when bacteria were grown in rich media, LB. Incorporation of these substrates into phospholipids was determined and expressed relative to PE levels. aThr, L-allo-threonine; cFA, cyclopropane fatty acid; Da, Dalton; FA, fatty acid; GP, glycerophosphate; Hse, L-homoserine; LB, Luria-Bertani broth (Miller); m/z, mass-to-charge ratio; M9, M9 minimal media; MS^2^, tandem mass spectrometry; OD_600_, optical density at wavelength 600 nm; PE, phosphatidylethanolamine; PO_3_^-^, phosphate; PS standard, phosphatidylserine standard; Thr, L-threonine; XIC, extracted ion chromatogram. ∗The analytical method does not distinguish between mono-unsaturated fatty acid and cyclopropane fatty acid.
When E. coli was cultured in the defined minimal media containing glucose as the sole exogenous metabolite, the unknown feature was detected (Fig. 2B, control), suggesting the bacterium was able to produce this lipid from endogenous substrates. In contrast, the unknown feature was not detected in P. aeruginosa cultured in the same defined media (Fig. S2, control). Considering the substrates for the lipid formation were endogenously formed, we tested the bacterium’s ability to incorporate L-threonine (Thr), L-allo-threonine (aThr), and L-homoserine (Hse) into its lipids. Threonine is a proteinogenic amino acid, while L-allo-threonine is a rare isomer, which can be metabolized by E. coli (42). Homoserine, on the other hand, is a nonproteinogenic amino acid and a metabolic intermediate for the synthesis of methionine and threonine. Considering the multiple pathways an amino acid can be involved in, each metabolite was supplied in excess to ensure an adequate supply of substrates for lipid incorporation. At high concentration (15 mM (equivalent to 0.18% (w/v))), L-homoserine was previously shown to restrict the growth of E. coli in minimal media (43). In this study, E. coli cultured in minimal media with 0.1% (w/v) L-homoserine and L-allo-threonine exhibited reduced growth (Figs. 2A and S1). However, in both conditions, the bacterium was able to form colonies, and sufficient biomass was obtained for lipid analysis.
The lipid composition of E. coli treated with these substrates was analyzed using LC/MS^2^. Figure 2B showed the extracted ion chromatograms (XIC), based on the major ions of the endogenous lipid found in E. coli. Interestingly, E. coli treated with L-allo-threonine and L-homoserine, but not L-threonine, led to accumulation of lipids detected at distinct retention times. The lipids formed from L-allo-threonine treatment co-eluted with the PS standard, while the lipids formed from L-homoserine treatment co-eluted with the endogenous lipid detected in E. coli. In contrast, no observable accumulation of lipids was detected in the presence of L-threonine. MS^2^ profiles of the respective major ions formed from L-allo-threonine and L-homoserine incorporation revealed the signature of 101 Da neutral loss, as well as the expected sum fatty acid composition of the endogenous lipid (33 carbons, with one double bond or one cyclopropane fatty acid) (Fig. 2C). Based on the combination of retention time and spectra similarity, we concluded that the endogenous lipid produced by E. coli is PHS. The possibility of the E. coli lipid being PT was ruled out based on the distinct retention time of lipids accumulation in P. aeruginosa PAO1 treated with L-threonine (Fig. S2).
Indeed, P. aeruginosa PAO1 exhibited differences in incorporation of the different substrates, in contrast to E. coli (Fig. 2D). Unexpectedly, although PHS was not detected in P. aeruginosa PAO1 cultured in rich media (Fig. 1C) or minimal media (Fig. S2B, control), we observed an accumulation of PHS in P. aeruginosa PAO1 grown in minimal media supplemented with L-homoserine (Fig. 2D). This accumulation was not present when the bacterium was grown in rich media supplemented with L-homoserine, unlike in E. coli, which was able to accumulate PHS in both media types (Fig. 2D). Since L-homoserine is an intermediate in essential amino acid synthesis, its availability may be the limiting factor in PHS formation. To test the effects of L-homoserine concentration on incorporation into phospholipids, E. coli was cultured with varying concentrations of the substrate. The reduction of growth rate of the bacterium is dependent on the concentration of L-homoserine, consistent with earlier reports (43) (Fig. S3A). Nonetheless, the concentration of L-homoserine supplemented exogenously to E. coli influenced the degree of incorporation into phospholipids to form PHS (Fig. S3B). The bacterium, despite the slowed growth at higher L-homoserine concentration, was able to accumulate high levels of PHS, suggesting the formation of PHS is independent of the growth rate.
Substrate promiscuity of EcPssA is responsible for the formation of PHS
We next investigated the enzymatic machinery responsible for PHS formation. L-homoserine and L-threonine differ structurally in terms of the position of the hydroxy group. Since E. coli does not produce PT, and lacks a TgPTS homolog, we ruled out the possibility of the activity of a PTS-like enzyme. Kanfer and Kennedy reported the limited effects on PssA activity when threonine was used as a substrate (11), but this does not exclude the possibility the involvement of PssA in PHS synthesis. Homoserine harbors a free terminal hydroxyl-group, which is present in serine. Indeed, this terminal hydroxy-group is involved in the condensation of serine with CDP-DAG to form PS. We hence questioned if EcPssA is responsible for the synthesis of PHS. A mutant of PssA (E. coli strain AH930) was generated by DeChavigny et al., which lacked PE and required divalent metal ions, including Mg^2+^, to grow (18). LC/MS analysis of the lipid extracts of E. coli strain AH930 confirmed the absence of PHS, in contrast to its parental WT strain, E. coli W3110 (Fig. 3A, top panels). To determine if the incorporation of the exogenously introduced substrates was dependent on EcPssA, both WT E. coli W3110 and ΔpssA mutant, E. coli AH930, were treated with L-allo-threonine and L-homoserine. WT E. coli W3110 was able to incorporate both substrates (Fig. 3A, left panels) to form phosphatidyl-allo-threonine (PaT) and PHS, consistent with the findings using E. coli strain BW25113 (Fig. 2B). E. coli AH930, on the other hand, did not accumulate any PHS or PaT (Fig. 3A, right panels). These results confirmed PHS is formed endogenously in E. coli through the activity of EcPssA, using L-homoserine as a substrate. Additionally, the substrate preference of EcPssA also extends to L-allo-threonine.Figure 3**Formation of phosphatidylhomoserine through the activity of EcPssA.**A, PS biosynthetic pathway is responsible for the incorporation of PHS in Escherichia coli. Extracted ion chromatograms of major PHS ions in WT E. coli W3110 (top left panel), which was used to further probe for their presence in both WT and AH930 ΔpssA mutant, untreated and treated with 0.1% (w/v) L-homoserine and L-allo-threonine. WT E. coli W3110 was able to produce PHS and PaT in the presence of L-homoserine and L-allo-threonine, while the signals were close to noise level for E. coli ΔpssA mutant (AH930), under all the conditions. B, extracted ion chromatogram of 18:1/18:1 PHS produced in a reaction mixture containing EcPssA (black line) or lacking the enzyme (gray line). C, MS^2^ spectra of enzymatically synthesized 18:1/18:1 PHS and its chemical structure. D, Michaelis–Menten kinetics of EcPssA with L-homoserine. Initial reaction velocities were measured using varying concentrations of L-homoserine (0.5, 1.0, 2.5, 5.0, 10.0, 15.0, and 25.0 mM) in the presence of 0.3 mM 18:1/18:1 CDP-DAG at 25 °C. Data points represent the mean ± SD (n = 3). /s, per second; aThr, L-allo-threonine; CDP-DAG, cytidine diphosphate diacylglycerol; EcPssA, E. coli phosphatidylserine synthase; FA, fatty acid; GP, glycerophosphate; Hse, L-homoserine; kcat, app, apparent catalytic constant; Km, app: apparent Michaelis constant; m/z, mass-to-charge ratio; MS^2^, tandem mass spectrometry; PaT, phosphatidyl-allo-threonine; PHS, phosphatidylhomoserine; PS, phosphatidylserine; XIC, extracted ion chromatogram.
To further confirm the utilization of L-homoserine by EcPssA as a substrate, enzymatic assays using purified EcPssA was performed. Incubation of L-homoserine and 18:1/18:1 CDP-DAG as substrates in the presence of EcPssA led to the formation of 18:1/18:1 PHS, as confirmed by LC/MS^2^ (Fig. 3, B and C). Using a spectrophotometric-based kinetic assay, we found that EcPssA displayed an apparent kcat (kcat, app) of 2.86 ± 0.04 s^−1^ and apparent Km (Km, app) of 4.6 ± 0.02 mM (Fig. 3D). In comparison, kcat and Km for L-serine is 69.7 ± 7.5 s^−1^ and 0.14 ± 0.03 mM respectively, as previously determined (44). These findings collectively confirmed EcPssA can use L-homoserine as a substrate, albeit with low efficiency, and is responsible for the production of endogenous PHS.
Decarboxylation of PHS to form PPA endogenously in Enterobacteriaceae
The ability of E. coli to synthesize other phospholipids is not restricted to PHS. Interestingly, in WT E. coli treated with L-homoserine, we detected an accumulation of another peak that closely eluted with PE (Fig. 4A), which was confirmed using MS^2^ to be the decarboxylated form of PHS, PPA (Fig. 4B, bottom). PPA forms fragment ions (m/z 154.03 and 210.05), which are 14 Da larger than the dehydrated phosphoethanolamine and glycerophosphoethanolamine fragments (m/z 140.01 and m/z 196.04) (45) formed from PE (Fig. 4B, top). To determine if PPA is produced in bacteria endogenously, the lipidomics data of Enterobacteriaceae, P. aeruginosa and A. baumannii were screened for the headgroup diagnostic ions, parent ion mass, and retention time of PPA. PPA was detected in all Enterobacteriaceae analyzed at very low level, but not in P. aeruginosa and A. baumannii (Fig. 4C), which is consistent with the absence of PHS in the latter bacterial species. The results overall suggested E. coli and other Enterobacteriaceae can decarboxylate PHS, albeit at a lower efficiency in contrast to the formation of PE from PS.Figure 4**Endogenous formation of phosphatidylpropanolamine.**A, representative chromatogram of Escherichia coli W3110 cultured without and with 0.1% (w/v) L-homoserine, focusing on the elution region of phosphatidylethanolamine (PE). Bacteria cultured with L-homoserine accumulated an additional peak eluting after PE (red arrow). B, decarboxylation of PHS in E. coli treated with 0.1% (w/v) L-homoserine to form phosphatidylpropanolamine (PPA). MS^2^ spectra of PE (top, dark red) and PPA (bottom, orange). The decarboxylation of PHS was evident from the fragment ions with m/z 154.03 and 210.05 which are 14 Da larger than the phosphoethanolamine-linked fragment ions formed by PE. The data were derived from one set of experiment and are representative of three biological replicates. C, detection of endogenous PPA in Enterobacteriaceae at baseline. PPA was detected in Enterobacteriaceae and not in other bacterial species. cFA, cyclopropane fatty acid; Da, Dalton; FA, fatty acid; Hse, L-homoserine; m/z, mass-to-charge ratio; MS^2^, tandem mass spectrometry; PE, phosphatidylethanolamine; PHS, phosphatidylhomoserine; PPA, phosphatidylpropanolamine. ∗The analytical method does not distinguish between mono-unsaturated fatty acid and cyclopropane fatty acid.
Effects of L-homoserine on global phospholipidome of E. coli
Based on the combination of genetics, substrate incorporation, and LC/MS^2^ characterization, we identified PHS as a phospholipid class found in E. coli under baseline condition, when grown in both rich (LB) and minimal media. This also led us to uncover the presence of an even lower abundant lipid PPA, which is formed from the decarboxylation of PHS endogenously. Figure 5A summarized the classes of phospholipids and the associated number of molecular ions characterized in E. coli BW25113 when cultured in LB and M9 minimal media, using LC/MS^2^. The number of PHS and PPA ions detected is expectedly less than the major diacylated PE and PG (Fig. 5A and Table S1A), given their natural low abundance. The relative distribution of phospholipids with distinct fatty acyl chain length was dependent on the time point and the type of media (Fig. 5B). The major PHS in LB (both time points) as well as M9 minimal media at 24-h harbored a total of 33 carbons in the fatty acyl chains, and for bacteria grown in M9 minimal media at 8-h, the major forms comprised 32 and 34 carbons (Fig. 5B and Table S1B). Bacteria grown in LB at 8-h are in the deceleration phase of growth (Fig. S4) and it is expected there was accumulation of phospholipids with odd-numbered sum composition, since it is well-characterized that stationary phase bacterium accumulated fatty acids with 17-carbons and cyclopropane ring (35, 46, 47). In general, the carbon chain length of the highest abundance molecular ion of PHS mirrored that of both PE and PG (Fig. 5B).Figure 5**Baseline phospholipidome of Escherichia coli and the effects of L-homoserine on phospholipid composition.**A, number of phospholipids detected and characterized in E. coli BW25113 grown in LB- and M9-based media. The numerical values represent the number of ions detected and characterized, with MS^2^ data for each ion from at least one of the samples analyzed in E. coli BW25113 grown in LB- and M9-based media for 8 and 24 h. Each ion is annotated at the species level (sum of carbon atoms and double bonds). B, relative distribution of total carbon numbers of fatty acyl chains found in the diacylated phospholipids, PG, PE, and PHS. The size of the circles denoted the relative peak area of a specific carbon number (sum of both fatty acyl chains), normalized to all ions detected for a given phospholipid class. Colors of the borders of circles denote the coefficient of variance (CV, %) from three biological replicates. C, changes in phospholipid class distribution during culture in LB supplemented with 0.1% (w/v) L-homoserine. Heatmap represented the fold change of the proportion of the peak areas attributed by each phospholipid class, relative to the total peak areas of all phospholipid classes detected. Data were generated from three biological replicates, and statistical analysis was performed using Student’s t test, with Bonferroni correction. Symbols represented adjusted p-values: ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001, ∗∗∗∗p < 0.0001. Box plots representing the raw peak areas were presented for the four classes of lipids, PE, PG, PHS, and PPA, with the most significant differences when E. coli was treated with L-homoserine, to confirm these specific changes observed were not due to the normalization method. D, phospholipid pathway of E. coli updated based on this study. Phospholipid names in black were detected in this study, with bold representing phospholipid classes confirmed with MS^2^ characterization. Acyl phosphatidylglycerol (AcylPG) and lysophosphatidylglycerol (LPG) were detected but no MS^2^ data was available due to the low abundance. AcylPG, acyl phosphatidylglycerol; CDP-DAG, cytidine diphosphate diacylglycerol; CdsA, phosphatidate cytidylyltransferase; CL, cardiolipin; ClsA/B/C, cardiolipin synthase A/B/C; CO_2_, carbon dioxide; FA, fatty acyl; G3P, glycerol-3-phosphate; Hse, Homoserine; LB, Luria-Bertani broth (Miller); LPE, lysophosphatidylethanolamine; LPG, lysophosphatidylphosphatidylglycerol; M9, M9 minimal media; MS^2^, tandem mass spectrometry; PA, phosphatidic acid; PE, phosphatidylethanolamine; PG, phosphatidylglycerol; PGP, phosphatidylglycerol phosphate; PgpA/B/C, phosphatidylglycerophosphatase A/B/C; PgsA, phosphatidylglycerol phosphate (PGP) synthase; PHS, phosphatidylhomoserine; PlsB, glycerol-3-phosphate acyltransferase; PlsC, 1-acyl-sn-glycerol-3-phosphate acyltransferase; PPA, phosphatidylpropanolamine; PS, phosphatidylserine; Psd, phosphatidylserine decarboxylase; PssA, phosphatidylserine synthase.
To understand how the incorporation of L-homoserine affects global phospholipid remodeling in E. coli, we analyzed the lipids in E. coli BW25113, treated with L-homoserine. Cells were grown in LB, which was more favorable for growth of the bacterium under control and L-homoserine–treated conditions, in contrast to M9 minimal media (43) (Fig. S4). L-homoserine treatment led to pronounced accumulation of PHS and PPA at both 8-h and 24-h time points (Fig. 5C and Table S2). A concomitant and significant reduction in PE and PG levels was observed, suggesting potential substrate funneling of CDP-DAG, a common intermediate metabolite for the synthesis of PE, PG, and PHS. In contrast to the formation of PE from PS, which leads to undetectable levels of PS and high levels of PE, the relative level of PHS is higher than PPA, suggesting less efficient decarboxylation of PHS compared to PS. The phospholipid metabolic pathway of E. coli, based on the collective biochemical and genetics findings from this study, is summarized in Figure 5D.
Discussion
In this work, we reported the discovery and characterization of PHS and PPA, two new phospholipid members in the baseline lipidome of E. coli, which were also detected in other Enterobacteriaceae species analyzed. We further provided genetic and biochemical evidence that PHS is formed through the activity of EcPssA. Treatment of E. coli with exogenous L-homoserine led to the accumulation of PHS and its decarboxylated form, PPA, as well as a concomitant reduction of PG and PE. To date, PHS has only been reported in Mycoplasma fermentans, with its structure resolved based on the analysis of hydrolyzed products (48). Our works provided the first structural characterization of intact PHS molecules and the biochemical pathway involved in its endogenous production in E. coli. Endogenous production of PPA, on the other hand, has yet to be reported in any organism.
In E. coli, the main enzymatic activity of PssA is the production of PS using CDP-DAG and serine as the primary substrates (11). Using an in vitro purified enzyme system, EcPssA was able to incorporate phosphoglycerol to form PG, albeit at low levels, but it lacked activity towards threonine (11). We provided biochemical evidence that EcPssA is able to incorporate L-homoserine into PHS. In addition, at the cellular level, the bacterium is able to use both L-homoserine and L-allo-threonine, to form PHS and PaT, respectively. Both L-homoserine and L-allo-threonine are isomers of L-threonine, demonstrating that EcPssA has restricted activity towards L-threonine possibly due to its stereochemistry (44). Homoserine differs from serine by an insertion of an additional CH_2_ unit, but it retains a free terminal hydroxy group, the functional group in serine involved in condensation with CDP-DAG. The catalytic efficiency (kcat/Km) of EcPssA for L-serine and L-homoserine differ significantly, with L-serine being the preferred substrate by approximately 270-fold. This difference likely stems from the confined nature of the amino acid–binding pocket, as suggested by the serine-bound model of EcPssA, which reveals multiple ionic and polar interactions that are critical for recognition of the cognate substrate and precise positioning of its nucleophilic hydroxyl group (44). The intracellular concentration of serine in WT E. coli has been reported to be ∼70 μM (49), whereas the concentration of homoserine has, to our knowledge, not been quantified, likely due to its low abundance. Taken together, the kinetic parameters of EcPssA support a physiological preference for L-serine over L-homoserine. Interestingly, E. coli was able to incorporate L-allo-threonine but not L-threonine, which are diastereomers. L-allo-threonine and L-threonine differ in the spatial position of the methyl group bound to the β-carbon, which may be a possible steric hindrance for the interaction of L-threonine with the active site of EcPssA. This postulation will require detailed structural analysis of purified EcPssA and its interactions with potential substrates. PHS had been produced in vitro with a reconstituted enzymatic reaction using cabbage or Streptomyces sp. PLD, with phosphatidylcholine and homoserine as substrates (50). Although E. coli harbors PLD activities (30, 51, 52), our works ruled out the possibility of PHS formation through other PLD, based on our findings that ΔpssA mutant does not incorporate L-homoserine into its lipids.
The specificity of incorporation is dependent on the organism, as well as the growth media. This is evident from our findings that unlike E. coli, P. aeruginosa does not produce detectable levels of PHS at baseline in both LB and M9 minimal media. This is not due to the sequence differences in the PS synthase in the two bacteria species, since P. aeruginosa was able to incorporate L-homoserine into its phospholipids when grown in M9 minimal media supplemented with the substrate. The formation of PHS in E. coli is dependent on the concentration of L-homoserine, suggesting that PHS synthesis is regulated by substrate availability. Homoserine is an endogenous metabolic intermediate which is formed from aspartate and serves as a precursor for the amino acids, threonine and methionine. Therefore, it is not surprising to observe low levels of PHS in E. coli and its absence in P. aeruginosa, if L-homoserine turnover and hence availability differs between bacterial species. Moreover, cultivation in rich and minimal media creates distinct pools of metabolites, which influence both the metabolic state of the bacterium and substrate availability (53, 54, 55). Interestingly, L-homoserine at high concentration (15 mM (equivalent to 0.18% (w/v))) leads to growth restriction of E. coli when grown in minimal media, but not rich media (43). Despite the growth restrictions in minimal media, E. coli was able to produce high levels of PHS in the presence of exogenous L-homoserine, suggesting EcPssA remained active despite the impeded growth rate. Whether the incorporation of L-homoserine to phospholipids is linked to restricted growth or served as a protective mechanism to buffer this metabolite will warrant in-depth investigation.
The substrate promiscuity of phospholipid metabolizing enzymes is clearly not restricted to PssA, evident from the production of PPA when E. coli was treated with L-homoserine. PPA is likely to be formed through decarboxylation of PHS, through the activity of Psd. In contrast, PPA had been reported in eukaryotic system through the incorporation of propanolamine as a substrate in cell cultures (56, 57, 58). In the budding yeast, Saccharomyces cerevisiae, PPA, formed from the exogenous provision of propanolamine to the cells can rescue the growth of psd1/psd2 double mutant which lacked PE (58). PHS, produced in vitro using PLD, had been shown to weakly activate the PS flippase, Atp8a1 (59). The precise functions of endogenously produced PHS and PPA in E. coli, however, will require further investigation. Nonetheless, the renewed knowledge on the substrate promiscuity of enzymes involved in phospholipid synthesis, along with the structural specificity of the enzyme-substrate relationship could potentially guide the design of novel probes for metabolic labeling (60, 61) to facilitate future studies on lipid metabolism, transport, distribution, and functions.
Cellular lipidome is evidently shaped by both endogenous and exogenous chemicals, which can be directly incorporated into lipids (30, 31). This is demonstrated in the present study, where L-homoserine and L-allo-threonine were incorporated into phospholipids in E. coli. Nonclassical lipids can be produced through various mechanisms: (i) broad substrate specificity of known enzymes, (ii) presence of domains with additional activities in known enzymes, (iii) presence of unknown enzymes, and (iv) nonenzymatic reactions. As a unicellular organism which resides in both the natural physical environment and human hosts (62), E. coli is exposed to varying environments with different biochemicals. However, laboratory media for cultivating bacteria such as LB (63) had been tailored towards promoting growth but does not truly reflect the environment the organism is exposed to. Our works provided the first cellular evidence that E. coli is able to produce other lipids endogenously using its existing known biochemical resources, specifically, PssA and endogenous homoserine. A more exacting picture of the global lipidome of E. coli and other bacteria species will require more systematic analysis involving conditions that mimic the external growth environment of the bacteria, including pH, temperature, and other exogenous compounds. With a more thorough understanding of the biochemistry of E. coli and other microbes, they can further serve as powerful biological factories for the production of atypical lipids. In this study, the metabolic plasticity of E. coli had served as an important source of lipid production which facilitated the characterization of PHS and PPA.
This work combined analytical chemistry, genetics, and biochemistry to confirm the structure and biosynthetic mechanism of PHS, a previously uncharacterized phospholipid class in E. coli. However, two bioanalytical limitations exist. Firstly, there is a lack of pertinent standards for the nonclassical lipids, as well as phospholipids with cyclopropane fatty acyls, which is common in E. coli and other bacteria (35, 46, 64, 65). It is hence important to note that the presented values based on peak areas do not reflect the actual lipid concentration, since peak areas depend on both concentration and ionization efficiency of the lipid. Furthermore, bacterial cell shape and biomass change during growth (66, 67). Hence, the pairwise (control vs treatment) comparison of the relative levels of lipid classes were based on normalization to the total phospholipids detected to control the amounts of materials. Raw peak areas were presented to ensure the observed changes in PHS, PPA, PG, and PE were not due to the normalization method or lack of internal standards. Secondly, MS^2^ was performed using collision-induced dissociation, which lacked the resolving power for double bonds position, and fatty acyl chain modifications including branch chain and cyclopropane rings. Hence, to avoid misreporting of lipid structures, we had examined the lipid composition at the carbon chain length level within each given class, and due consideration was given for annotating fatty acids sharing the same m/z for monounsaturated and cyclopropane forms. The fine structures of the fatty acyl constituents can be further resolved using alternative methods, including ultraviolet photodissociation mass spectrometry (35, 64), ozone dissociation (68) or Paternò-Büchi derivatization, and LC/MS^2^ (69). Certainly, more work will be required to establish both qualitative and quantitative bacterial lipidomics and their applications in research to deepen our current understanding of bacterial lipid metabolism and functions.
Experimental procedures
Bacteria strains
Bacteria and strains used in this study include E. coli BW25113 (KEIO library, WT), P. aeruginosa PAO1 and S. aureus ATCC 29213 (from Kevin Pethe), E. coli W3110 and AH930 (ΔpssA mutant) (from William Dowhan, (18)), E. coli EC958 (70), K. pneumoniae ATCC10031 (ATCC), K. pneumoniae VA360, A. baumannii BC5, Escherichia hormaechei BEI01, K. aerogenes UCI15 (BEI resources).
General bacteria culture conditions
Bacteria precultures were grown overnight in LB broth (Miller), incubated at 37 °C with shaking at 150 rpm in Infors HT Ecotron incubator. The optical density (OD) at wavelength 600 nm (OD_600_) of the overnight cultures was measured using Thermo Scientific Multiskan GO Microplate Spectrophotometer. The cultures were then inoculated in LB at an OD_600_/ml of 0.01 or 0.05 for growth at 37 °C with shaking at 150 rpm, up to the respective time points, for sampling. For W3110 and AH930, the media were supplemented with 23.4 mM MgCl_2_.
For initial characterization of lipidome of E. coli, K. pneumoniae, K. aerogenes, A. baumannii, P. aeruginosa, and S. aureus, cultures were sampled at 24-h. The cultures were centrifuged at 3000g for 10 min at 4 °C to remove the media. The samples were washed twice in cold M9 wash media (1× commercial M9 without any supplementation of salt or carbon source), and the final pellets were snap frozen with liquid nitrogen and stored at −80 °C.
Substrate incorporation for WT E. coli BW25113 and P. aeruginosa PAO1
Overnight bacterial precultures were centrifuged at 3000g for 10 min at room temperature to remove the rich media and washed once with M9 wash media. The pellets were resuspended in M9 minimal media containing 0.1 mM CaCl_2_, 2 mM MgSO_4_, 0.2% (w/v) glucose, and supplemented with the respective substrate (0.1%, w/v in water) to achieve a final A_600_/ml of 0.01. A starting concentration of 0.1% (w/v) of the substrates was used to ensure adequate supply for downstream biosynthesis pathways and potential incorporation into lipids. Substrates used include L-homoserine, L-threonine, and L-allo-threonine. The cultures were incubated at 37 °C with shaking at 150 rpm. After 8 h and 24 h, the OD_600_ of the cultures were measured, and sampling was performed for viability testing by spotting on LB agar. The choice of these time points was based on the logarithmic phase of E. coli BW25113 and P. aeruginosa PAO1 in minimal media supplemented with glucose (control) (8-h), and 24-h represented the stationary phase of growth. To determine the incorporation of L-homoserine in rich media, parallel cultures were set up and grown in LB or M9, supplemented with 0.1% (w/v) L-homoserine. Due to observable difference in growth rates, which is dependent on the bacterial species, culture media, and treatment, all sampling were fixed at these two time points.
For lipid analysis, the cultures were centrifuged at 3000g for 10 min at 4 °C to remove the media. The samples were washed once in cold M9 wash media, and the final pellets were snap frozen with liquid nitrogen and stored at −80 °C. Three sets of biological replicates were collected.
Characterization of phospholipids and substrate incorporation in PssA E. coli mutant
ΔpssA mutant (AH930) and the parental WT (W3110) were grown in LB media supplemented with 23.4 mM MgCl_2_ using the above culture conditions. For substrate incorporation, the bacterial cultures were grown in LB media with 23.4 mM MgCl_2_ supplemented with the respective substrates at a final concentration of 0.1% (w/v in water) for 8 h and 24 h.
Sampling was performed by centrifugation at 3000g for 10 min at 4 °C, followed by two washes with M9 wash media. The final pellets were snap frozen with liquid nitrogen and stored at −80 °C. Three sets of biological replicates were collected, except for the substrate incorporation for 24 h which had two sets of biological replicates.
Lipid extraction and analysis using LC/MS2
Lipids were extracted using a modified Bligh-Dyer method (71). Briefly, chloroform:methanol, 1:2 (v/v) were added to the bacterial pellets and the samples were incubated at 6 °C for 4 h. After the incubation, water and chloroform were added, followed by centrifugation for phase separation. The lower organic phase was collected and the upper phase was re-extracted with chloroform. The organic extracts were pooled and dried using either a SpeedVac (Labconco) or nitrogen stream. Lipid extracts were stored at −80 °C until further analysis.
For lipid characterization, mass spectrometry (MS) and MS^2^ were performed using a SCIEX TripleTOF 6600 quadrupole time-of-flight mass spectrometer, coupled to an Agilent 1290 liquid chromatography system. Normal phase chromatography (Intersil SIL column, 3 μm, 2.1 × 150 mm, GL Sciences) was used for separation, and lipids were analyzed in the negative electrospray ionization mode. The mobile phase A consisted of chloroform/methanol/ammonia hydroxide (89.5/10/0.5, v/v/v) and mobile phase B consisted of chloroform/methanol/water/ammonia (55/39/5.5/0.5). Automated MS^2^ analyses were performed using collision-induced dissociation in the information-dependent acquisition mode. The lipidomics minimal reporting checklist (72) is made available in Supplemental Material.
LC/MS data analysis
For qualitative analysis, PeakView (SCIEX) was used for the visual review of spectra and extraction of ion chromatograms and MS^2^ spectra. Data lists were then exported into .txt files, and the XIC and MS^2^ spectra were plotted using the R package ggplot. All MS^2^ spectra and XICs presented are from one experiment and are representative of the three independent biological replicates. To determine the relative incorporation of substrates into phospholipids, peak areas of XIC of three most abundant molecular species of the respective lipid classes (if present) were integrated in PeakView and calculated as a percentage of the equivalent PE molecular species. For assessing the global lipidome, full dataset was extracted using mass spectrometry-data independent analysis software, version 4.8 (73). The relative peak areas of each sample were calculated by dividing the peak area of individual phospholipids by the sum of all identified phospholipids, after filtering lipids with less than 40% CV between the three biological batches. Fold change between conditions was computed, and statistical analysis was performed using Student’s t test with Bonferroni correction. For the relative distribution of phospholipids by their total fatty acyls carbon chain length within each phospholipid class, ions with the same carbon number within a phospholipid class were summed up and further normalized to the sum of all molecular ions of the given class. All statistical analysis and graphical production were performed using the R packages rstatix and ggplot (74, 75).
For lipid annotation, the following criteria must be fulfilled, with the exception for the currently characterized lipids, which were discussed in the results section: (i) retention time of lipid class, based on synthetic lipid standards (Avanti Polar Lipids), (ii) MS^2^ spectra matched for at least one sample of a given E. coli strain.
Functional activity of purified EcPssA using L-homoserine as a substrate
The activity of purified E. coli (K-12 strain MG1655) PssA (EcPssA) using L-homoserine as a substrate was assessed using two approaches, as described by Lee et al. (44). Firstly, formation of PHS was determined using LC/MS using a reconstituted enzymatic assay, carried out following a modified approach described by Dowhan (76). The assay mixture was prepared by mixing 0.4 mM 18:1/18:1 CDP-DAG, 1 mM L-homoserine, and 1 mg/ml of bovine serum albumin. The reaction was initiated by adding 100 nM purified EcPssA in buffer A, composed of 30 mM Tris–HCl, pH 7.5, 400 mM NaCl, 10% (v/v) glycerol, and 0.05% (w/v) n-octyl-β-D-glucoside (OG) and subsequently incubated at 37 °C for 30 min. The reaction was terminated using five volumes of chloroform/methanol (2:1, v/v) mixture, followed by vigorous vortexing. Lipid extraction was carried out using the Folch method (77) and analyzed using LC/MS analysis (44).
EcPssA activity was further assessed by a coupled spectrophotometric assay. The assay quantitatively monitors the release of CMP from CDP-DAG and L-homoserine as substrates, based on the decrease in NADH absorbance at 340 nm, as previously described by Carman and Dowhan (78, 79). The reaction mixture contained 50 mM Tris–HCl, pH 7.5, 0.1 M KCl, 10 mM MgCl_2_, 1 mg/ml bovine serum albumin, 1 mM ATP, 1 mM phosphoenolpyruvate, 0.2 mM NADH, 0.2 U/ml of cytidine monophosphate kinase, 0.4 U/ml of pyruvate kinase, 0.4 U/ml of lactate dehydrogenase, 2.4 mM Triton X-100, and 0.3 mM 18:1/18:1 CDP-DAG substrate and varying concentration of L-homoserine in a total volume of 0.18 ml. Enzyme reactions were conducted at 25 °C and initiated by the addition of EcPssA. The sample was briefly sonicated before absorbance measurements. The curve was fitted to the Michaelis–Menten equation using nonlinear regression in OriginPro 2021. All experiments were performed in triplicate, and data are presented as mean ± SD.
Data availability
All data used in the article are available from the corresponding author upon reasonable request.
Supporting information
This article contains supporting information.
Conflict of interest
The authors declare that they have no conflicts of interests with the contents of this article.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Nikaido H.Molecular basis of bacterial outer membrane permeability revisited Microbiol. Mol. Biol. Rev.6720035936561466567810.1128/MMBR.67.4.593-656.2003 PMC 309051 · doi ↗ · pubmed ↗
- 2Carty C.E.Ingram L.O.Lipid synthesis during the Escherichia coli cell cycle J. Bacteriol.1451981472478700732510.1128/jb.145.1.472-478.1981 PMC 217296 · doi ↗ · pubmed ↗
- 3Raetz C.R.Dowhan W.Biosynthesis and function of phospholipids in Escherichia coli J. Biol. Chem.2651990123512382404013 · pubmed ↗
- 4Shibuya I.Metabolic regulations and biological functions of phospholipids in Escherichia coli Prog. Lipid Res.311992245299128766710.1016/0163-7827(92)90010-g · doi ↗ · pubmed ↗
- 5Parsons J.B.Rock C.O.Bacterial lipids: metabolism and membrane homeostasis Prog. Lipid Res.5220132492762350045910.1016/j.plipres.2013.02.002PMC 3665635 · doi ↗ · pubmed ↗
- 6Zhang Y.M.Rock C.O.Membrane lipid homeostasis in bacteria Nat. Rev. Microbiol.620082222331826411510.1038/nrmicro 1839 · doi ↗ · pubmed ↗
- 7Dowhan W.A retrospective: use of Escherichia coli as a vehicle to study phospholipid synthesis and function Biochim. Biophys. Acta 183120134714942292563310.1016/j.bbalip.2012.08.007PMC 3513495 · doi ↗ · pubmed ↗
- 8Raetz C.R.Enzymology, genetics, and regulation of membrane phospholipid synthesis in Escherichia coli Microbiol. Rev.42197861465936215110.1128/mr.42.3.614-659.1978 PMC 281446 · doi ↗ · pubmed ↗